БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
27. МЕХАНИЗМЫ ВИДООБРАЗОВАНИЯ
27.8. Внутривидовое видообразование
Внутривидовое видообразование предполагает участие нескольких факторов, однако во всех случаях непременным условием является прекращение обмена генами между популяциями. В результате каждая субпопуляция становится генетически изолированной. Изменения частоты аллелей и генотипов в отдельных популяциях, обусловленное действием естественного отбора на диапазон фенотипов, создавшихся в результате мутаций и половой рекомбинации, ведет к образованию рас и подвидов. Если генетическая изоляция сохраняется в течение длительного периода времени, а затем подвиды встречаются вновь в той же самой области, то они либо снова скрещиваются, либо скрещивание между ними оказывается невозможным. В случае успешного скрещивания их все еще можно считать принадлежащими к одному виду. Невозможность скрещивания означает, что произошло видообразование, и прежние подвиды следует теперь рассматривать как самостоятельные виды. Полагают, что именно таким образом могут происходить эволюционные изменения.
Начальным фактором в процессе видообразования может служить снижение интенсивности давления отбора в популяции. Это может привести к повышению внутривидовой изменчивости. Если новые фенотипы обладают адаптациями к условиям среды, имеющимся в краевых участках ареала, то популяция имеет возможность расширить область своего распространения. В тех случаях, когда обмен генами внутри популяции не ослабевает, данный вид проявляет локализованную фенотипическую изменчивость (экотипы), все еще сохраняет общий генофонд и продолжает оставаться единым видом. В таких ситуациях обычно создаются клины.
Видообразование может происходить только при условии возникновения преград, ведущих к репродуктивной изоляции между членами данной популяции. Репродуктивная изоляция создается изолирующими механизмами (термин, введенный Т. Добржанским) того или иного типа.
27.8.1. Изолирующие механизмы
Изолирующий механизм — это средство, создающее и поддерживающее репродуктивную изоляцию внутри какой-либо популяции. Репродуктивная изоляция может быть обусловлена механизмами, действующими до или после оплодотворения. В табл. 27.3 приведена в несколько модифицированном виде классификация изолирующих механизмов, предложенная Добржанским.
Таблица 27.3. Изолирующие механизмы (по Добржанскому)
Презиготические механизмы (преграды для образования гибридов) |
|
Сезонная изоляция |
Брачный период или сроки цветения двух видов приходятся на разное время года; например, в Калифорнии Pinus radiata цветет в феврале, а Pinus attenuata — в апреле |
Экологическая изоляция |
Два вида, обитающие в одной и той же области, предпочитают различные местообитания. Например, Viola arvensis растет на известковых почвах, а Viola tricolorпредпочитает кислые почвы |
Поведенческая изоляция |
Животные проявляют специфические для каждого вида ритуалы ухаживания, на которые реагируют только особи того же вида, что предотвращает возможность спаривания между особями разных видов. Это наблюдается, например, у определенных видов рыб, птиц и насекомых |
Механическая изоляция |
У животных успешной копуляции препятствуют различия в строении половых органов; у растений опыление близкородственных видов осуществляется различными животными |
Постзиготические механизмы (преграды, возникающие у гибридов) |
|
Нежизнеспособность гибридов |
Образование гибридов возможно, но они не достигают зрелости, как, например, гибриды между северной и южной расами леопардовой лягушки (Rana pipiens) в Северной Америке |
Стерильность гибридов |
Гибриды не способны производить функциональные гаметы, как, например, мул (2n = 63), получающийся в результате скрещивания между лошадью (Equus equus, 2n = 60) и ослом (Equus hemionus, 2n = 66) |
Неполноценность гибридов |
Гибриды F1 фертильны, но поколение F2 и потомки от возвратных скрещиваний между гибридами F1 и родительскими формами либо не развиваются, либо стерильны, как, например, у гибридов между разными видами хлопчатника (Gossypium) |
27.8.2. Аллопатрическое видообразование
Для аллопатрического (от греч. állos — другой, patris — родина) видообразования характерно, что на одной из его стадий существенную роль играет пространственное разобщение. Такое разобщение, создаваемое географическими преградами (например, горными хребтами, морями или реками) или же различиями в предпочитаемых местообитаниях, может препятствовать потоку генов: организмы или их гаметы теряют возможность встретиться и это ведет к репродуктивной изоляции. Адаптация к новым условиям или случайный дрейф генов в небольших популяциях приводят к изменениям частоты аллелей и генотипов. В результате длительного разобщения популяций между ними может возникнуть генетическая изоляция, сохраняющаяся даже в том случае, если они вдруг снова окажутся вместе. Таким образом могут возникать новые виды. Например, разнообразие видов вьюрков семейства Geospizidae и их распространение на островах Галапагосского архипелага считают результатом аллопатрического видообразования.
По мнению Дэвида Лэка, первоначально вьюрки прибыли на острова с южно-американского материка и здесь в отсутствие конкуренции со стороны местных видов (слабое давление отбора) произошла их адаптивная радиация, приведшая к возникновению разнообразных видов, приспособившихся к различным экологическим нишам. Эволюция разных видов, происходившая, как полагают, в условиях географической изоляции, зашла так далеко, что когда впоследствии они снова встречались на некоторых островах, то могли сосуществовать как самостоятельные виды.
27.8.3. Симпатрическое видообразование
Генетические различия могут накапливаться и в таких аллопатрических популяциях, которые были географически изолированы на протяжении гораздо более коротких периодов времени. Если эти популяции впоследствии встречаются, то в зонах перекрывания могут возникнуть гибриды. Например, на Британских островах обитают черная ворона (Corvus соrone) и серая ворона (Corvus соrone comix). У черной вороны окраска целиком черная, и эта форма распространена в Англии и на юге Шотландии. У серой вороны черные только бока, а спина и брюшко серые; обитает она в северной части Шотландии. Гибриды между серой и черной воронами занимают узкую полосу в центральных районах Шотландии (рис. 27.15). У этих гибридов низкая фертильность и это эффективно препятствует обмену генами между популяциями серой и черной ворон (постзиготические изолирующие механизмы).
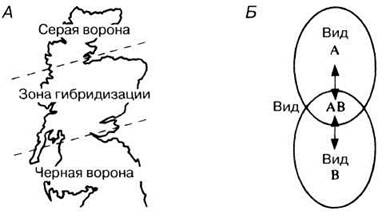
Рис. 27.15. Зона гибридизации как преграда, препятствующая обмену генами между двумя популяциями. Благодаря наличию этой зоны, пересекающей Шотландию поперек, два вида ворон остаются обособленными (А). Б. Существование таких преград между соседними популяциями наблюдается часто; они действуют следующим образом: в тех местах, где географические ареалы видов А и В перекрываются, в результате скрещиваний между ними появляются гибриды с пониженной фертильностью; в дальнейшем вид А свободно скрещивается с гибридом А В, и АВ — с видом В, но присутствие гибрида АВ препятствует свободному скрещиванию между популяциями А и В.
Со временем может возникнуть отбор, направленный против скрещивания между этими двумя формами, что приведет к видообразованию. Поскольку такое видообразование происходит в конечном счете в одной географической области, его называют симпатрическим видообразованием (от греч. syn — вместе, patris — родина).
Симпатрическое видообразование не связано с географическим разобщением популяций в период создания генетической изоляции. Для него необходимо развитие какого-либо механизма репродуктивной изоляции, возникающего в результате отбора в пределах географически изолированной области. Этот механизм может быть структурным, физиологическим, поведенческим или генетическим.
Симпатрическое видообразование чаще рассматривают как механизм, позволяющий объяснить, каким образом близкие виды, которые, вероятно, произошли от общего предка в результате временной изоляции, могут сосуществовать как обособленные виды в одной и той же географической области. Например, на Галапагосских островах вьюрок Camarhynchus pauper обитает только на острове Чарльз, где он сосуществует с родственной формой С. psittacula, широко распространенной на всех центральных островах (рис. 27.16). Виды вьюрков при выборе брачных партнеров руководствуются, по-видимому, величиной клюва. Диапазоны величины клюва у С. pauper на острове Чарльз и у С. psittacula на острове Албемарл примерно одинаковы, но на острове Чарльз у С. psittacula клюв длиннее. Такое отличие уже достаточно велико, чтобы эти два вида, которые различаются по характеру питания, не привлекали друг друга во время брачного периода. Таким образом, виды сохраняют обособленность и способны к сосуществованию.

Рис. 27.16. Распространение двух видов вьюрков на Галапагосских островах как пример сосуществования видов после симпатрического видообразования.
27.8.4. Кольцевые виды
Симпатрическое видообразование особого типа наблюдается в той точке, где две популяции, занимающие краевые участки клины, встречаются и обитают совместно в одной области, замыкая таким образом кольцо. Например, чайки рода Larus образуют непрерывную популяцию, окружающую Северный полюс кольцом между 50 и 80° с. ш. В это кольцо входит 10 рас или подвидов, различающихся главным образом по общим размерам тела и окраске ног, спины и крыльев. Между всеми этими расами происходит свободный обмен генами, если не считать того места, где на Британских островах «концы кольца» смыкаются. Здесь, в крайних точках ареала, чайки ведут себя как два отдельных вида — серебристая чайка (Larus argentatus) и клуша (L. fuscus). Они различаются по внешнему виду, по крику, по характеру миграций и скрещиваются между собой лишь в редких случаях. Отбор, направленный против их гибридизации, происходит в условиях симпатрии.
У видов с половым размножением симпатрическое видообразование без географической изоляции маловероятно. Однако у организмов, размножающихся бесполым путем, в том числе у высших растений с вегетативным размножением, один мутант, достаточно отличающийся от родительской популяции, чтобы быть генетически изолированным, может симпатрически дать начало новому виду. Примером служит полиплоидия у Spartina (разд. 24.9.2).
27.3. Десять подвидов популяции Larus argentatus-fuscus образуют непрерывное кольцо, которое тянется от Британских островов, через Скандинавию, Россию, Берингов пролив, Аляску и Канаду, и возвращается к Британским островам. Если бы подвиды, обитающие в районе Берингова пролива, исчезли, то каковы были бы последствия для всей этой популяции?