БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
23. НЕПРЕРЫВНОСТЬ ЖИЗНИ
23.6. ДНК
23.6.2. Репликация ДНК
Модель структуры ДНК в виде двойной спирали, предложенная Уотсоном и Криком, описана в разд. 3.6.3. Одна из самых привлекательных особенностей этой модели состоит в том, что она одновременно показывает, каким способом может происходить репликация ДНК. Уотсон и Крик высказали предположение, что две цепи, образующие спираль, могут раскручиваться и разделяться; при этом они служат матрицами, к которым путем спаривания оснований пристраивается комплементарная цепочка нуклеотидов. Таким образом из каждой исходной молекулы ДНК возникают две копии с идентичной структурой.
В 1956 г. Корнбергу удалось продемонстрировать in vitro синтез молекулы ДНК, используя в качестве матрицы одиночную цепь ДНК. Корнберг выделил из Е. coli и очистил фермент, который способен связывать друг с другом свободные нуклеотиды в присутствии АТФ как источника энергии с образованием комплементарной цепи ДНК. Он назвал этот фермент ДНК-полимеразой. Как показали дальнейшие эксперименты, нуклеотиды, используемые в клетке, содержат две дополнительные фосфатные группы. Это активирует нуклеотиды. По мере прикрепления каждого нуклеотида к растущей цепи ДНК две дополнительные фосфатные группы отщепляются. Освобождающаяся при этом энергия используется оставшейся фосфатной группой нуклеотида для образования связи с остатком сахара в молекуле соседнего нуклеотида. Процесс репликации показан на рис. 23.20. Он начинается с раскручивания двойной спирали ДНК, контролируемого ферментом геликазой. Затем ДНК-полимераза прикрепляется к одноцепочечной ДНК и начинает перемещаться вдоль цепи. Всякий раз, когда она доходит до очередного основания в цепи ДНК, свободные нуклеотиды приближаются к цепи, и тот из них, который содержит комплементарное основание, образует с ним водородные связи. Свободный нуклеотид удерживается ферментом на месте до тех пор, пока он не присоединится к предыдущему нуклеотиду, наращивая таким образом новую цепь ДНК. Это наращивание может происходить только в направлении 5' → 3'. Как можно видеть на рис. 23.20, это означает возможность непрерывного копирования только одной цепи ДНК (на рисунке верхняя), так как ДНК-полимераза движется в том же направлении, что и раскручивающий фермент. Этот процесс называют непрерывной репликацией. Копирование другой цепи (нижней на рис. 23.20) должно всякий раз начинаться вновь, потому что ДНК-полимераза должна удаляться от раскручивающего фермента в направлении 5' → 3'. В результате в цепи возникают небольшие разрывы, потому что ДНК-полимераза не может соединить 3'-конец одного новосинтезируемого участка ДНК с 5'-концом следующего. Для того чтобы ликвидировать этот разрыв, необходим другой фермент — ДНК-лигаза. Такую репликацию называют прерывистой.
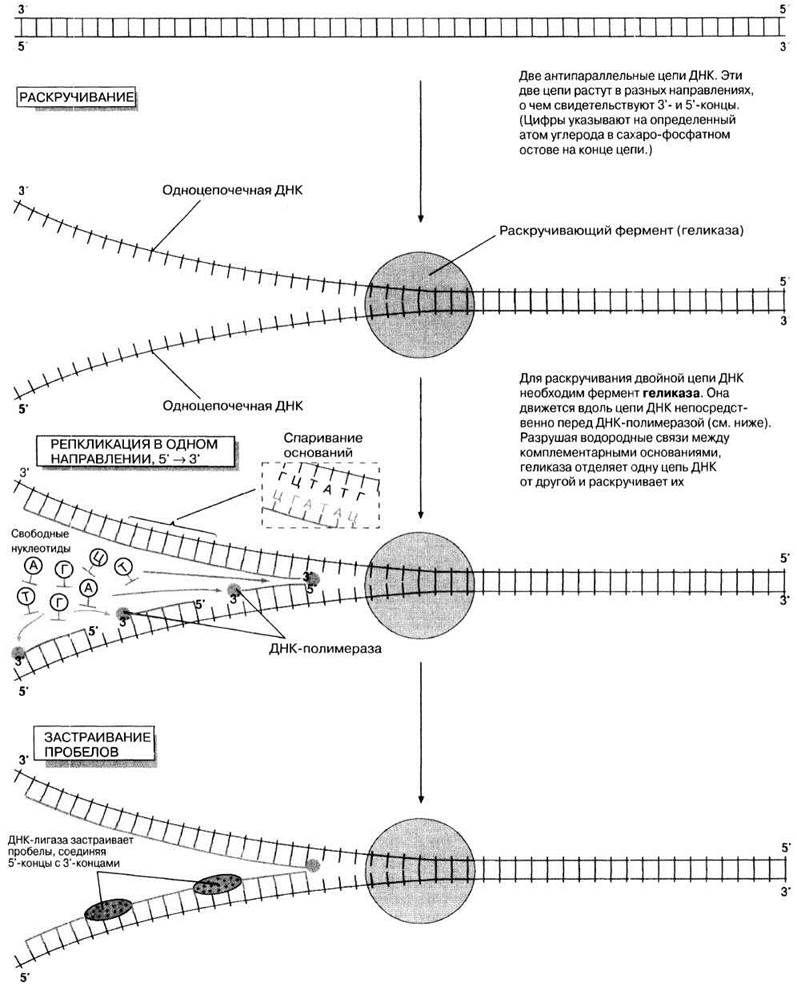
Рис. 23.20. Репликация ДНК. Упрощенная схема, на которой показаны главные принципы. В репликации участвуют также другие молекулы и ферменты, не показанные на схеме.
Данные в пользу механизма полуконсервативной репликации
Способ репликации ДНК, предложенный Уотсоном и Криком и показанный на рис. 23.20, известен под названием полуконсервативной репликации, так как при этом каждая новая двойная спираль сохраняет одну из двух цепей исходной двойной спирали ДНК. Этот механизм основан на данных, полученных Мезелсоном и Сталем в серии классических экспериментов в 1958 г. Клетки Е. coli содержат одну кольцевую хромосому; при культивировании этих клеток в течение многих поколений на среде, содержащей тяжелый изотоп азота (15N), вся их ДНК оказывалась меченной этим изотопом. Клетки с меченой ДНК переносили на среду, содержавшую обычный изотоп азота 14N. По прошествии срока, соответствующего времени генерации Е. coli (50 мин при 36 °С) — времени, необходимого для одного клеточного деления и одной репликации ДНК, — отбирали пробы клеток, выделяли из них ДНК и центрифугировали в течение 20 ч при 40 000 g в растворе хлористого цезия (CsCl). При этом тяжелые молекулы CsCl начинали осаждаться, образуя градиент плотности, которая нарастала от верхушки к дну пробирки. ДНК собиралась на том уровне, на котором плотность раствора CsCl была равна ее плотности. При исследовании в ультрафиолете ДНК в центрифужной пробирке имела вид узкой полосы. Расположение полос ДНК, выделенной из клеток, культивированных на средах с 15N и 14N, и интерпретация полученных данных представлены на рис. 23.21. Схема полуконсервативной репликации включена в рис. 23.22. Эти эксперименты убедительно показали, что репликация ДНК происходит полуконсервативным способом.
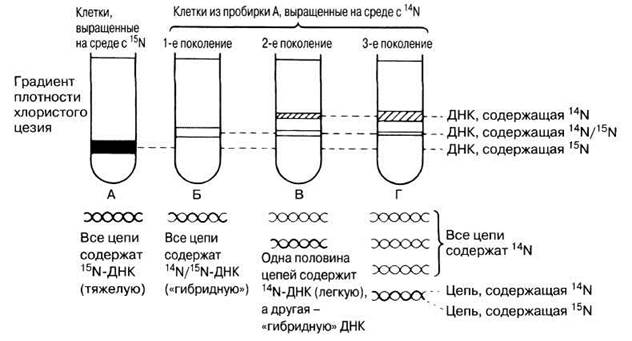
Рис. 23.21. Результаты экспериментов Мезелсона и Сталя и их интерпретация. Ширина полос ДНК в центрифужных пробирках отражает относительные количества молекул ДНК различных типов. В пробирке В отношение ширины полос 1:1, а в пробирке Г— 3:1.
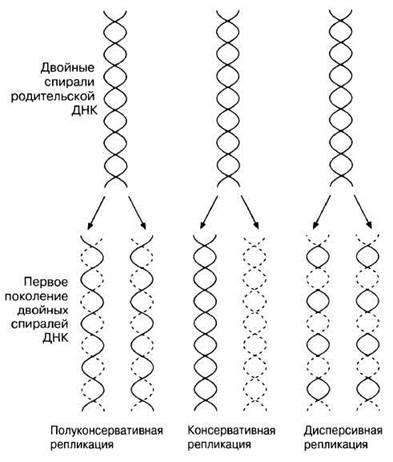
Рис. 23.22. Схема, поясняющая три теории репликации ДНК.
23.2. Для объяснения процесса репликации ДНК были выдвинуты три гипотезы. Одна из них получила название полуконсервативной репликации и описана выше. Две другие известны как консервативная репликация и дисперсивная репликация.
Все три гипотезы представлены на рис. 23.22.
Нарисуйте схемы распределения различных типов ДНК в градиенте плотности, которые обнаружили бы Мезелсон и Сталь для двух первых поколений клеток, earn бы были верны гипотезы консервативной и дисперсивной репликации.