БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
24. ИЗМЕНЧИВОСТЬ И ГЕНЕТИКА
24.9. Мутации
Мутацией называют изменение количества или структуры ДНК данного организма. Мутация приводит к изменению генотипа, которое может быть унаследовано клетками, происходящими от мутантной клетки в результате митоза или мейоза. Мутация может вызвать изменения какого-либо признака в данной популяции. Если мутация возникла в гаметах, то она передается по наследству, тогда как мутации соматических клеток наследуются только дочерними клетками, образующимися в результате митоза. Последние называют соматическими мутациями.
Мутации, выражающиеся в изменении числа или структуры хромосом, известны под названием хромосомных мутаций или хромосомных аберраций (перестроек). При некоторых аберрациях хромосомы изменяются так сильно, что это видно под микроскопом. В настоящее время термин «мутация» используют главным образом для обозначения изменений ДНК в одном локусе, когда происходит так называемая генная, или точковая мутация.
Представление о мутации как о причине внезапного появления нового признака было впервые выдвинуто в 1901 г. голландским ботаником Гуго де Фризом, изучавшим наследственность у энотеры Oenothera lamarckiana. Спустя 9 лет Т. Морган начал изучать мутации у дрозофилы, и вскоре при участии генетиков всего мира у нее было идентифицировано более 500 мутаций.
24.9.1. Частота мутаций и их причины
Мутации возникают случайным образом и спонтанно, т. е. любой ген может мутировать в любой момент. Частота возникновения мутаций у разных организмов различна.
В результате работ Г. Дж. Мёллера в двадцатые годы XX в. было установлено, что частоту мутаций можно повысить по сравнению с их спонтанным уровнем, воздействуя на организм рентгеновскими лучами. В дальнейшем выяснилось, что частоту мутаций можно значительно повысить при помощи высокоэнергетического электромагнитного излучения, такого как ультрафиолет, рентгеновские и γ-лучи. Частицы высокой энергии — α- и β-частицы, нейтроны и космическое излучение — также обладают мутагенным действием, т. е. способны вызывать мутации. Мутагенным действием обладают и разнообразные химические вещества, в частности иприт, кофеин, формальдегид, колхицин, некоторые компоненты табака и все возрастающее число лекарственных препаратов, пищевых консервантов и пестицидов.
24.9.2. Хромосомные мутации
Хромосомные мутации могут быть результатом изменения числа хромосом или их структуры. Некоторые типы хромосомных мутаций изменяют действие нескольких генов и оказывают на фенотип гораздо более глубокое воздействие, чем генные мутации. Изменения числа хромосом обычно происходят в результате ошибок при мейозе, но они возможны и при нарушениях митоза. Эти изменения выражаются либо в анэуплоидии — утрате или добавлении отдельных хромосом, либо в эуплоидии (полиплоидии) — добавлении целых гаплоидных наборов хромосом.
Анэуплоидия
Анэуплоидия может выражаться в появлении в дочерних клетках добавочной хромосомы (n + 1), (2n + 1) и т. д. или в нехватке какой-либо хромосомы (n - 1), (2n - 1) и т. д. Анэуплоидия может возникнуть, если в анафазе I мейоза гомологичные хромосомы одной или нескольких пар не разойдутся. В этом случае оба члена пары направляются к одному и тому же полюсу клетки, и тогда разделение гомологичных хромосом в анафазе II может привести к образованию гамет, содержащих на одну или несколько хромосом больше или меньше, чем в норме (рис. 24.31). Это явление известно под названием нерасхождения. Когда гамета с недостающей или лишней хромосомой сливается с нормальной гаплоидной гаметой, образуется зигота с нечетным числом хромосом: вместо каких-либо двух гомологов в такой зиготе их может быть три или только один.
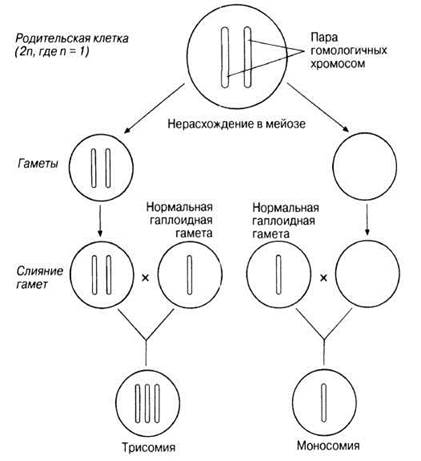
Рис. 24.31. Нерасхождение хромосом при образовании гамет и результаты слияния аномальных гамет с нормальными гаплоидными клетками. Это приводит к различным типам полисемии, при которой число хромосом может быть равно 2n + 1 (трисомия), 2n + 2 (тетрасомия), 2n + 3 (пентасомия) и т. д.; или же 2n - 1 (моносомия), — в зависимости от числа гомологичных хромосом, не разошедшихся нормальным образом (см. также рис. 25.34.)
Зигота, в которой число хромосом меньше диплоидного, обычно не развивается, но зиготы с лишними хромосомами иногда способны к развитию. Если это происходит у животных, то из таких зигот в большинстве случаев развиваются особи с резко выраженными аномалиями. Среди наиболее часто встречающихся хромосомных мутаций, возникающих у человека в результате нерасхождения хромосом, — одна из форм трисомии, называемая синдромом Дауна (2n = 47). Такое состояние, названное по фамилии врача, описавшего его впервые в 1866 г., вызывается нерасхождением хромосом 21-й пары. Эта мутация описана в разд. 25.7.6.
Возможно также нерасхождение мужских и женских половых хромосом, которое приводит к анэуплоидии, влияющей на вторичное половые признаки и фертильность, а иногда и на умственные способности (разд. 25.7.7 и 25.7.8).
Эуплоидия (полиплоидия)
Гаметы и соматические клетки с увеличенным числом хромосом, кратным их гаплоидному числу, называют полиплоидными. Приставки три-, тетра- и т. д. указывают, во сколько раз увеличено число хромосом, т. е. степень плоидности: 3n — триплоид, 4n — тетраплоид, 5n — пентаплоид и т. д. У растений полиплоидия встречается гораздо чаще, чем у животных. Например, из 300 000 известных видов покрытосеменных примерно половина — полиплоиды. Относительная редкость полиплоидии у животных объясняется тем, что увеличение числа хромосом значительно повышает вероятность ошибок при образовании гамет в мейозе. Что касается растений, то большинство из них способно к вегетативному размножению, а поэтому они могут воспроизводиться и в полиплоидном состоянии. Полиплоидные организмы часто обладают благоприятными признаками — более крупными размерами, выносливостью, устойчивостью к заболеваниям. Это их свойство называют гибридной мощностью (разд. 27.4.2). Большинство наших культурных растений — полиплоиды, образующие крупные плоды, запасающие органы, цветки или листья.
Существуют две формы полиплоидии — аутополиплоидия и аллополиплоидия.
АУТОПОЛИПЛОИДИЯ. Аутополиплоидия может возникнуть как естественным, так и искусственным путем в результате увеличения числа хромосомных наборов у данного вида. Например, если произойдет репликация хромосом (в интерфазе) и хроматиды нормально разойдутся (в анафазе), но цитоплазма не разделится, то образуется тетраплоидная (4n) клетка с большим ядром. Такая клетка затем делится и дает тетраплоидные дочерние клетки. Количество цитоплазмы в этих клетках возрастает, чтобы сохранилось постоянное ядерно-плазменное отношение, и это ведет к увеличению общих размеров растения или какой-нибудь его части. Аутополиплоидию можно вызвать с помощью алкалоида колхицина, выделяемого из клубнелуковицы безвременника (Colchicum). В концентрации порядка 0,01% колхицин подавляет образование веретена деления, разрушая микротрубочки, так что хромосомы не расходятся во время анафазы. Колхицин и близкие к нему вещества использовали для создания ценных сортов таких культур, как табак, томаты и сахарная свекла. Аутополиплоиды могут быть так же плодовиты, как диплоиды, если они содержат четное число хромосомных наборов.
У животных иногда наблюдается видоизмененная форма полиплоидии, при которой образуются отдельные полиплоидные клетки и ткани. Этот процесс, называемый эндомитозом, состоит в репликации хромосом, не сопровождающейся разделением клетки. Гигантские хромосомы в клетках слюнных желез дрозофилы и тетраплоидные клетки в печени человека возникают в результате эндомитоза.
АЛЛОПОЛИПЛОИДИЯ. Аллополиплоидия возникает в случае удвоения числа хромосом у стерильного гибрида, в результате чего он становится фертильным. Гибриды F1, получаемые при скрещиваниях между разными видами, обычно стерильны, так как их хромосомы не могут образовывать гомологичные пары во время мейоза. Это явление называют гибридной стерильностью. Однако если число хромосом становится кратным их исходному гаплоидному числу, например, 2 (n1 + n2), 3 (n1 + n2) и т. д. где n1 и n2 — гаплоидные числа хромосом у родительских видов, то возникает новый вид, который дает фертильных гибридов при скрещивании с такими же полиплоидами, но стерилен при скрещивании с одним из родительских видов.
Для большинства аллополиплоидных видов характерно диплоидное число хромосом, которое представляет собой сумму диплоидных чисел родительских видов. Например, Spartina anglica (2n = 122) — фертильный гибридный аллополиплоид, полученный в результате скрещивания Spartina maritima (stricta) (2n = 60) со Spartina altemiflora (2n = 62). Гибрид F1, полученный от скрещивания между этими двумя видами, стерилен и получил название Spartina townsendii (2n = 62). Большинство аллополиплоидных растений отличается по своим признакам от обоих родительских видов; среди них многие из весьма ценных культур, выращиваемых человеком. Например, вид пшеницы Triticum aestivum (2n = 42), из которого получают муку, был выведен путем скрещиваний и отбора на протяжении 5000 лет. В результате скрещивания пшеницы-однозернянки (2л = 14) с неидентифицированным диплоидом (2n = 14) был получен новый вид пшеницы — эммер (2n = 28). Пшеницу эммер скрещивали с еще одним дикорастущим видом (2n = 14), в результате чего была создана форма Triticum aestivum (2n = 42), которая представляет собой гексаплоид (6n) исходной пшеницы-однозернянки. Другой пример межвидовой гибридизации — скрещивание редьки с капустой — будет описан в разд. 27.9.
У животных аллополиплоидия почти неизвестна, так как у них межвидовые скрещивания происходят редко. В результате полиплоидии генофонд не получает новых генов (разд. 27.4.2), но возникают новые комбинации генов.
Структурные изменения хромосом
При кроссинговере во время профазы I мейоза происходит реципрокный обмен генетическим материалом между гомологичными хромосомами. Это ведет к изменению последовательности аллелей в родительских группах сцепления, в результате чего получаются рекомбинанты, но без потери каких-либо генных локусов. Сходные эффекты возникают при таких хромосомных перестройках, как инверсии и транслокации. При перестройках других типов — делециях и дупликациях — число генных локусов в хромосомах изменяется, и это может оказать глубокое влияние на фенотипы. Структурные изменения в хромосомах, связанные с инверсиями, делениями, дупликациями, а в некоторых случаях и транслокациями, можно наблюдать под микроскопом, когда в профазе I мейоза гомологичные хромосомы приступают к конъюгации. Гомологичные хромосомы образуют синапсы (конъюгируют; разд. 23.3), а в тех участках, которые затронула перестройка, одна из гомологичных хромосом образует петлю или же перекручивается. Какая из хромосом образует петлю и как расположатся ее гены зависит от типа перестройки.
Инверсия возникает в результате вырезания какого-либо участка хромосомы, который поворачивается на 180°, а затем вновь встраивается на прежнее место. При этом никаких изменений генотипа не происходит, но возможны фенотипические изменения (рис. 24.32). Это свидетельствует о значении последовательности генов в данной хромосоме — так называемый эффект положения гена.
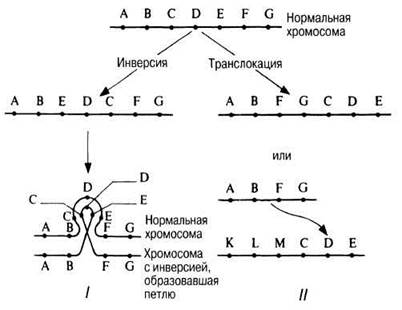
Рис. 24.32. Схематические изображения инверсии и транслокации и их влияния на расположение генов А — G. I Образование петли в профазе мейоза, вызванное инверсией. II. Участок хромосомы, несущий гены С, D и Е, оторвался и присоединился к хромосоме, несущей гены К, L и М.
При транслокации от одной из хромосом отрывается участок, присоединяющийся либо к другому концу той же хромосомы, либо к одному из концов другой, негомологичной, хромосомы (рис. 24.31). И в этом случае в фенотипе может проявиться эффект положения. Реципрокная транслокация между негомологичными хромосомами может привести к образованию двух новых пар гомологичных хромосом. В тех случаях, когда при синдроме Дауна сохраняется нормальное диплоидное число хромосом, причиной этого служит транслокация сверхкомплектной хромосомы 21 на одну из более крупных хромосом, обычно номер 15.
Самая простая форма хромосомной мутации — делеция, или нехватка, т. е. утрата хромосомой какого-нибудь участка, промежуточного или концевого, в результате чего в хромосоме недостает некоторых генов (рис. 24.33). Делеция может произойти в одной из двух гомологичных хромосом; в таких случаях аллели, находящиеся в другой, нормальной, хромосоме, экспрессируются, даже если они рецессивные. Если же делеция затронула одни и те же локусы в обеих гомологичных хромосомах, это обычно ведет к летальному исходу.
Иногда какой-либо участок хромосомы удваивается, так что возникает дупликация — повторение набора генов, локализованных в этом участке. Дополнительный набор может оказаться включенным в ту же хромосому, располагаться на одном из ее концов или же присоединяться к какой-нибудь другой хромосоме (рис. 24.33).
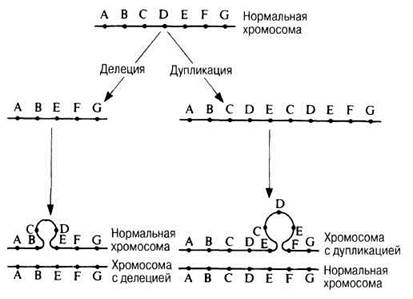
Рис. 24.33. Схематическое изображение делеции и дупликации и их влияния на расположение генов А — G. В обоих случаях образуются петли.
24.9.3. Генные мутации
Что такое генная мутация?
Внезапные спонтанные изменения фенотипа, которые не связаны с хромосомными аберрациями (что подтверждается данными микроскопических исследований), можно объяснить только изменениями в структуре отдельных генов. Генная или точковая мутация (поскольку она относится к определенному генному локусу) — результат изменения нуклеотидной последовательности молекулы ДНК в определенном участке хромосомы. Такое изменение последовательности оснований в одном гене воспроизводится при транскрипции мРНК и может привести к изменению последовательности аминокислот в полипептидной цепи, образующейся в результате трансляции на рибосомах.
Типы генных мутаций
Существуют различные типы генных мутаций, связанные с добавлением, выпадением или перестановкой оснований в данном гене. Они проявляются в форме дупликаций, вставок, делеций, инверсий или замены оснований. Во всех случаях они приводят к изменению нуклеотидной последовательности, а часто и к образованию измененного полипептида. Например, делеция вызывает сдвиг рамки считывания, последствия которого описаны в разд. 23.7.
Последствия генных мутаций
Генные мутации, возникающие во время формирования гамет, передаются всем клеткам потомков и могут оказывать влияние на дальнейшую судьбу вида. Соматические генные мутации, происходящие в организме, наследуются только теми клетками, которые образуются из мутантной клетки в результате митоза. Они могут оказать воздействие на тот организм, в котором они возникли, но со смертью данной особи исчезают из генофонда популяции. Соматические мутации, вероятно, возникают очень часто и остаются обычно незамеченными, но в некоторых случаях при этом образуются клетки с повышенной скоростью роста и деления. Эти клетки могут дать начало опухолям — либо доброкачественным, которые не оказывают влияния на другие ткани, либо злокачественным, паразитирующим на других тканях (рак, меланомы, саркомы).
Эффекты генных мутаций чрезвычайно разнообразны. Большая часть мелких генных мутаций фенотипически не проявляется, поскольку они рецессивны, однако известен ряд случаев, когда изменение всего лишь одного основания в генетическом коде оказывает глубокое влияние на фенотип. Одним из примеров служит серповидноклеточная анемия — заболевание, вызываемое у человека заменой одного основания в одном из генов, ответственных за синтез гемоглобина. Более подробно это заболевание и его причины описаны в разд. 25.7.2.
24.9.4. Значение мутаций
Хромосомные и генные мутации оказывают разнообразные воздействия на организм. Во многих случаях эти мутации летальны и нарушают развитие; у человека, например, около 20% беременностей заканчиваются самопроизвольным выкидышем в сроки до 12 нед. при этом у половины абортусов выявляются хромосомные аномалии. Если в результате некоторых хромосомных мутаций определенные гены оказываются рядом, то иногда их совместный эффект может привести к появлению какого-либо благоприятного признака. Кроме того, сближение некоторых генов делает менее вероятным их разделение в результате кроссинговера, а в случае благоприятных генов это создает преимущества.
Генная мутация может привести к тому, что в определенном локусе окажется несколько аллелей. Это увеличивает гетерозиготность данной популяции, делает более разнообразным ее генофонд и ведет к усилению внутрипопуляционной изменчивости. Перетасовка генов как результат кроссинговера, независимого распределения, случайного оплодотворения и мутаций может повысить непрерывную изменчивость, но ее эволюционная роль часто оказывается преходящей, так как возникающие при этом изменения могут быстро сгладиться. Что же касается генных мутаций, то некоторые из них увеличивают дискретную изменчивость, и это может оказать на популяцию более глубокое влияние. Большинство генных мутаций рецессивны по отношению к «нормальному» аллелю, который успешно выдержал отбор на протяжении многих поколений. Будучи рецессивными, мутантные аллели могут оставаться в популяции на протяжении многих поколений, пока им не удастся встретиться, т. е. оказаться в гомозиготном состоянии и проявиться в фенотипе. Время от времени могут возникать и доминантные мутантные аллели, которые немедленно проявляются в фенотипе (разд. 27.5, Biston betularia).
Материал, изложенный в этой главе, дает представление о происхождении внутрипопуляционной изменчивости и о механизмах наследования признаков, но он не объясняет, каким образом могло возникнуть поразительное разнообразие живых организмов, описанное в гл. 2. Попытка ответить на этот вопрос составляет содержание трех следующих глав.