БИОЛОГИЯ РАЗМНОЖЕНИЯ РАСТЕНИЙ - Н. Л. Колясникова - 2017
РАЗДЕЛ 3. ТИПЫ ОПЫЛЕНИЯ
Опыление - процесс переноса пыльцы на рыльце пестика. Различают два типа опыления: самоопыление и перекрестное опыление. У покрытосеменных растений преобладает перекрестное опыление. Реже встречается самоопыление. Постоянное самоопыление считают тупиком эволюции, ведущим к деградации.
3.1 Самоопыление
При самоопылении пыльца одного растения попадает на рыльце пестика этого же растения. Этот тип свойственен только обоеполым цветкам.
Различают приспособления к самоопылению: автогамия, гетоногамия, клейстогамия.
Автогамия осуществляется по-разному: при прямом соприкосновении - контактная автогамия, при высыпании пыльцы и оседании под собственным весом - гравитационная автогамия, под действием ветра - ветроавтогамия, с помощью мелких насекомых - трипсоавтогамия. Контактная автогамия характерна для седмичника, майника, копытня, гравитационная описана у одноцветки, ветроавтогамия встречается у голубики.
Гейтоногамия - это соседственное опыление, когда пыльца с одного цветка соцветия прорастает на рыльце пестика другого цветка этого же соцветия. Оно может происходить в конце сезона цветения. Ей способствует ветер, животные, насекомые. Встречается данный способ опыления у видов сем. Астровые, Капустные. Растения с таким способом опыления характеризуются мелкими цветками в густых соцветиях.
Клейстогамия - опыление в закрытом цветке. Различают облигатную и факультативную клейстогамию. Клейстогамные цветки мельче по размерам, части цветка редуцированы. Лепестки рудиментарны или отсутствуют. Число тычинок уменьшено, пыльцевые зерна мельче. Примеры растений - фиалка удивительная, кислица обыкновенная (рис. 20, 21).
Рис. 20. Двойное оплодотворение в клейстогамных цветках кислицы обыкновенной: 1 - опустошенный пыльник клейстогамного цветка после прорастания пыльцевых трубок, остались оболочки пыльцевых зерен, пыльцевые трубки и дегенерировавшие пыльцевые зерна; 2 - проникновение пыльцевой трубки между клетками стенки
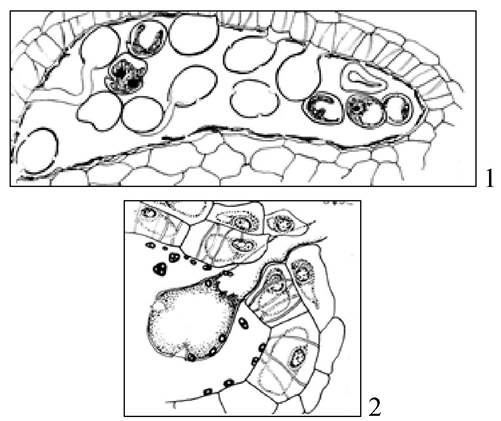
Рис. 21. Пыльцевые трубки в пыльнике клейстогамного цветка у фиалки кочкарной - 1; прорастание пыльцевых трубок через разрыв в стенке пыльника клейстогамного цветка у фиалки кочкарной - 2
Помимо клейстогамных цветков у этих растений образуются и хазмогамные цветки с открытым цветением. Факультативная клейстогамия не постоянна, возникает при определенных условиях. У них нет признаков редукции в цветке. Она связана с неблагоприятными факторами среды, встречается у ячменя, овса, пшеницы, ковыля и др.
3.2 Перекрестное опыление
При перекрестном опылении пыльца с цветков одного растения попадает на рыльца пестиков других растений данного вида.
Различают следующие приспособления к перекрестному опылению: двудомность, дихогамия, самонесовместимость, суточный ритм цветения.
Двудомность - разделение мужских и женских цветков по разным растениям одного вида. К двудомным видам относятся: облепиха, ива, тополь, осина, щавель, крапива, спаржа и др. Адаптивное значение разделения полов заключается в обеспечении перекрестного опыления и в дифференциации половых форм по экологическим нишам. По фенологии первыми зацветают мужские растения. Длительность цветения женских растений дольше. Тычиночные цветки крупнее пестичных. Число мужских цветков в соцветии и на растении в целом больше, чем женских.
Дихогамия - разновременное созревание пыльников и рылец. Это функциональная раздельнополость. Цветок выступает то в мужской, то в женской фазе. У разных видов встречаются разные типы: сначала мужская стадия - протерандрия, или сначала женская - протерогиния.
Протерандрия встречается часто. Широко представлена у растений класса двудольные. Иногда тычиночная и пестичная стадии четко разграничены по времени. Рыльца становятся восприимчивыми к пыльце, когда все тычинки данного цветка завяли.
Протерогиния встречается у растений класса однодольные и примитивных двудольных (сем. Лютиковые).
Самонесовместимость - это неспособность пыльцевых трубок прорастать в столбик на всю его длину и совершать оплодотворение. Гены, ответственные за самонесовместимость обозначаются S. Если пыльца и рыльце несут одинаковые аллели S-гена, то оплодотворение не происходит. Различают гетероморфную и гомоморфную самонесовместимость. Гетероморфная делится на ди- и тристилию. При гомоморфной самонесовместимости цветки одинаковы по строению. Гомоморфная самонесовместимость встречается чаще. Различают два генетических контроля: спорофитный и гамето- фитный. При спорофитной несовместимости способность пыльцы к прорастанию определяется генотипом спорофита, то есть растения, на котором идет образование микроспор. При спорофитном контроле подавление прорастания пыльцы происходит рано, уже на рыльце. Поверхность рылец сухая, покрыта пелликулой. Столбик закрытый. Пыльцевые зерна трехядерные. Это тип самонесовместимости встречается у растений сем. Астровые, Капустные, Гвоздичные и др.
При гаметофитной несовместимости способность к прорастанию пыльцы определяется ее генотипом (то есть генотипом самого гаметофита) (рис. 22).
Рис. 22. Рост пыльцевых трубок и оплодотворение при гаметофитной несовместимости
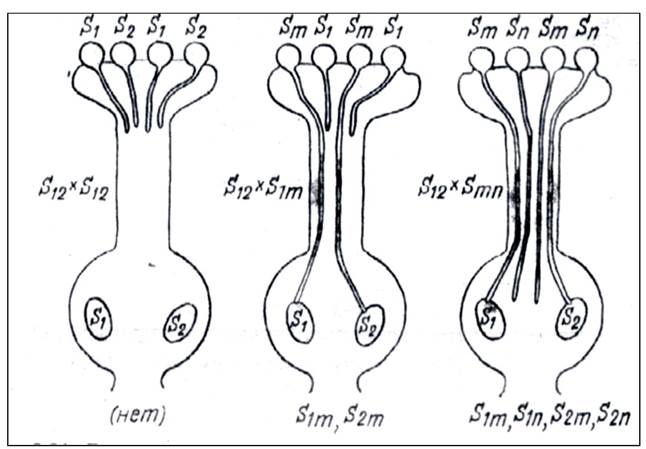
При гаметофитном контроле пыльцевые трубки сначала растут быстро, затем рост их тормозится и прекращается. Рыльца влажные, столбик - открытый. Пыльцевые зерна двуядерные. Гаметофитная самонесовместимость отмечена у растений сем. Бобовые, Лилейные, Розоцветные, Пасленовые и др.
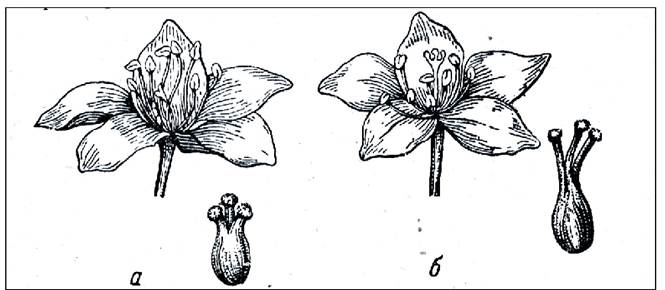
Дистилия - разностолбчатость. Впервые описана у примулы. Встречаются два типа цветков: длинностолбчатые и короткостолбчатые. Для них характерен диморфизм пыльцы: у короткостолбчатых растений пыльца крупнее, сосочки рыльца короче. Легитимным (законным) является опыление пыльцой с коротких тычинок на рыльце короткого столбика и, наоборот, пыльцой с длинных тычинок на рыльце длинного пестика. Также дистилия детально изучена у гречихи (рис. 23).
Рис. 23. Дистилия у гречихи: а - короткостолбчатая; б - длинностолбчатая форма
При тристилии (дербенник) три типа цветков - короткие столбики и длинные и средние тычинки, средние столбики и длинные и короткие тычинки, длинные пестики и средние и короткие тычинки.
Дистилия контролируется одним супергеном: генотип растений с короткостолбчатыми цветками SS и Ss, а растений с длинностолбчатыми цветками - ss. Между цветками одного морфотипа само- и перекрестное оплодотворение не осуществляется (иллегитимный вариант опыления), но оно возможно между растениями разных морфотипов (легитимный вариант опыления). При тристилии генетическое строение различных форм основано на двух локусах М и S: длинностолбчатые цветки: mmss, среднестолбчатые: Мmss или ММss,короткостолбчатые: МmSs, mmSs, ММSs, ММSS, mmSS.
Псевдосовместимость. У многих растений появление псевдосовместимости связано с искусственным опылением. На уровень псевдосовместимости влияют свет, температура, влажность. Существует псевдосовместимость «конца сезона», или при опылении бутонов, при удалении верхушки столбика, при использовании растительных гормонов.
Суточный ритм цветения. Большинство цветков реагируют на смену дня и ночи. Некоторые цветки открываются и закрываются несколько раз. Другие распускаются лишь один раз. В зависимости от приуроченности раскрывания цветков к определенному суточному периоду различают растения с утренним (одуванчик), дневным (люцерна), вечерним (матиолла), ночным (тропические растения), круглосуточным (брусника) и порционным взрывчатым (сем. Мятликовые) цветением. Суточный ритм распускания цветков рассматривается рядом ученых как проявление биоритмов.
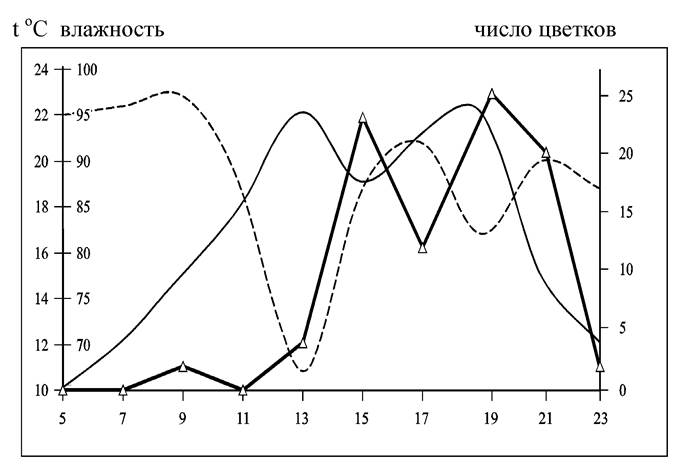
Методика наблюдений за суточным ходом цветения растений предложена А.Н. Пономаревым. Суточную динамику распускания цветков определяют в 50 соцветиях исследуемого вида в течение суток в трехкратной повторности. Через каждые два часа подсчитывается число вновь распустившихся цветков. При этом необходимо регистрировать температуру и относительную влажность воздуха (рис. 24).
Рис. 24. Суточный ход распускания цветков Astragalus onobrychis 30.06.1992 г.: температура, °С, - влажность, - число цветков
Различают два способа перекрестного опыления: биотическое и абиотическое.
Биотическое опыление. Опыление первоначально осуществлялось жуками. Они питались пыльцой. Затем с появлением нектарников главными опылителями стали перепончатокрылые, двукрылые, чешуекрылые. Таким образом, пыльца и нектар стали первичными аттрактантами. Пыльца содержит белки, жиры, углеводы, ферменты, витамины. У энтомофильных растений в пыльцевых зернах больше жиров и белка, у анемофильных - углеводов. Пыльца относительно долговечна. Нектар - это водный раствор сахаров: фруктозы, сахарозы и др. Также нектар включает аминокислоты, белки, органические кислоты, витамины и др. Концентрация сахара достигает 74% (у конского каштана). Выделение нектара зависит от времени дня, возраста и стадии развития цветка. Нектарники встречаются на разных органах цветка - на чашелистике, лепестках, цветоложе, тычиночных нитях.
Для привлечения опылителей помимо пищи служат вторичные аттрактанты: окраска и запах. Окраска обусловлена пигментами: флавоноидами, каротиноидами, антоцианом, антофеином. Часто окраска неоднородна, имеет узор или рисунок. Он служит указателем пути к нектару. У некоторых растений в течение жизни наблюдается изменение окраски, что связано с изменением рН клеточного сока вакуолей в клетках лепестков после осуществившегося оплодотворения. Например, у медуницы цветки сперва розово-пурпурные, затем - синие. Это сигнал для опылителей, что пыльцы нет.
Существует предпочтение окраски для разных опылителей. Пчелы не различают красный цвет, но восприимчивы к ультрафиолету. При одновременном цветении растений с синими и желтыми венчиками, пчелы в первую очередь опыляют синие цветки. В географическом плане тоже есть дифференциация по окраске. В тропиках преобладают красные цветки, в высокогорьях - белые. Желтые цветки - в пустынях и в Приполярье. Синие цветки - на равнинах в умеренном климате.
Аромат энтомофильных цветков разнообразен. Различают более 500 запахов. Изредка это трупный запах гниения у аронников, рафлезий. Иногда запах обусловлен аминами (боярышник, рябина). Чаще запах связан с эфирными маслами. Источник запаха - лепестки, тычинки, стаминодии, нектарники, пыльца. Интенсивность запаха колеблется и достигает максимума в определенное время суток. Например, петуния - ночью, клевер - днем.
Характеристика способов биотического опыления представлена в таблице 4.
Таблица 4. Способы биотического опыления
№ п/п |
Способ опыления |
Опылители |
Признаки цветка |
Примеры семейств растений (видов) |
1 |
Мелиттофилия |
Пчелы, осы, шмели |
Цветок зигоморфный с удобной посадочной площадкой, Цвет яркий, желтый или синий |
Яснотковые Норичниковые Бобовые |
2 |
Миофилия |
Мухи, комары |
Короткая трубка венчика цветка с открытым нектаром, цвет венчика желтый, если с неприятным запахом - красный |
Зонтичные Гвоздичные |
3 |
Психофилия |
Дневные бабочки |
Цветок актиноморфный, трубчатый или со шпорцем. Цвет яркий синий, красный |
Лилейные Гвоздичные |
4 |
Фаленофилия |
Ночные бабочки |
Цветок актиноморфный, трубчатый. Цвет белый |
Пасленовые |
5 |
Кантарофилия |
Жуки |
Цветок крупный, актиноморфный, плоский, чашеобразный. Окраска цветка часто зеленая, белая |
Магнолиевые Пальмы |
6 |
Мирмекофилия |
Муравьи |
Цветки мелкие, сидячие, нектарники неглубокие, пыльца мелкая и клейкая |
Горец, Дивала, Лобулярия |
7 |
Трипсофилия |
Трипсы |
Цветки зигоморфные, двугубые на коротких цветоножках |
Лобелия |
8 |
Орнитофилия |
Птицы |
Цветки зигоморфные, трубчатые, одиночные, прочные. Цвет венчика красный, оранжевый без запаха |
Банановые Миртовые Лилейные Бобовые Кактусовые |
9 |
Хироптерофилия |
Летучие мыши |
Цветок крупный, толстая цветоножка, неприятный запах. Цвет белый, кремовый |
Розоцветные Астровые Агавовые |
10 |
Опыление млекопитающими |
Мыши, лемуры, опоссумы |
Растение низкорослое. Цветки собраны в корзинки или головки. Запах дрожжевой. Много нектара |
Протейные Миртовые |
Абиотическое опыление. Различают два способа опыления: с помощью ветра - анемофилия и с помощью воды - гидрофилия. Характеристика способов абиотического опыления представлена в таблице 5.
Таблица 5. Способы абиотического опыления
№ п/п |
Способ опыления |
Фактор опыления |
Признаки цветка |
Примеры семейств растений |
1 |
Анемофилия |
Ветер |
Цветки не имеют околоцветника, собраны в соцветие, без окраски, запаха, нектара. Пыльца мелкая, экзина тонкая, гладкая |
Мятликовые Астровые Березовые Буковые |
2 |
Гидрофилия |
Вода |
Цветок без околоцветника, без запаха, нектара. Пыльца без экзины, крупная. |
Рдестовые Водокрасовые |
3.3 Половой полиморфизм
Разнообразие половых форм - это приспособление к перекрестному опылению.
Е.И. Демьянова [13] предлагает следующую классификацию половых форм у растений:
1. Гермафродитные. Популяция состоит из растений только с обоеполыми цветками.
2. Однодомные:
- Собственно однодомные. Мужские и женские цветки формируются на одной особи (береза, дуб, кукуруза и др.);
- Андромоноэцичные. На одном растении встречаются обоеполые и мужские цветки (сем. Сельдерейные, Розоцветные и др.);
- Гиномоноэцичные. На одном растении обоеполые и женские цветки (сем. Астровые и др.);
- Тримоноэцичные. На одном растении обоеполые, женские и мужские цветки (клен платановидный и др.).
3. Двудомные:
- Собственно двудомные. Популяция состоит из растений только с женскими и растений только с мужскими цветками (осина, тополь, ива, конопля и др.);
- Андродиэцичные. Популяция включает растения с обоеполыми цветками и растения только с мужскими цветками (чемерица, лютик, калужница и др.)
- Гинодиэцичные. Популяция состоит из растений с обоеполыми цветками и растений с женскими цветками (сем. Губоцветные, Гвоздичные и др.).
4. Трехдомные:
- Триэцичные. В популяции растения с обоеполыми цветками, растения с мужскими цветками и растения с женскими цветками (ясень обыкновенный, джузгун, шпинат, лютик стоповидный и др.);
- Полиэцичные. Наблюдается разнообразие сочетаний обоеполых, мужских и женских цветков в пределах трех и большего числа особей (виды клена).
Однодомные растения. Большинство исследователей считают, что однодомные растения произошли от гермафродитных. Они в большей мере характерны для класса Однодольные. Также наблюдается связь между однодомностью и анемофилией (рис. 25).
Рис. 25. Побег березы с мужскими и женскими соцветиями

По расположению мужских и женских цветков на растении выявлены некоторые закономерности. Чаще женские цветки находятся в нижней части соцветия и открываются первыми. По размерам они мельче.
По продолжительности цветения женские цветки превосходят мужские, иногда разница между цветением женских и мужских цветков составляет несколько дней. Тогда они выступают как двудомные.
Внешние условия, такие как температура, освещенность, минеральное питание, могут усиливать признаки одного пола. Также меняется соотношение цветков с возрастом. Например, у клещевины в молодом возрасте образуются преимущественно женские цветки, по мере старения - мужские. У некоторых видов соотношение цветков меняется в обратном направлении.
У андромоноэцичных растений соотношение между обоеполыми и мужскими цветками различное. Например, у каштана преобладают мужские цветки, кроме того они обычно мельче обоеполых цветков и расположены по периферии соцветия. По количеству пыльцы, темпам роста пыльцевых трубок мужские и обоеполые цветки не отличаются.
У гиномоноэцичных растений (преимущественно в сем. Астровые - тысячелистник, полынь, астра, девясил) соотношение обоеполых и женских цветков в соцветии стабильно, закреплено генетически. По краю корзинки находятся женские цветки, в центре - обоеполые.
У тримоноэцичных растений (сем. Злаковые) в нижней части соцветия мужские цветки, в средней - обоеполые, в верхней части - женские.
Двудомные растения. Соотношение полов подробно изучено именно у древесных растений. В большинстве случаев мужские растения преобладают над женскими (рис. 26). Также соотношение может быть 1:1 (осина, облепиха). Реже женские растения преобладают над мужскими.
Рис. 26. Кошачья лапка двудомная: 1 - женское растение; 2 - мужское растение

На соотношение полов оказывают влияние многие факторы:
- в чистых посевах однолетних растений преобладают женские растения из-за гибели мужских;
- в засушливых условиях уменьшается число женских растений;
- более устойчивы к действию ядовитых веществ и поражению болезнями женские растения;
- на засоленных почвах преобладают мужские растения (тополь, ива);
- на границе ареала встречается только одна половая форма и растения переходят к вегетативному размножению.
По фенологии первыми зацветают мужские растения. Длительность цветения женских растений продолжительнее. Число мужских цветков в соцветии и на растении в целом больше женских.
В эволюционном плане ученые считают, что обоеполые цветки первичны. Иногда у двудомных растений появляются обоеполые цветки, их рассматривают как атавизмы.
Адаптивное значение разделения полов заключается в обеспечении перекрестного опыления и в дифференциации половых форм по экологическим нишам.
Андродиэцичные растения встречаются довольно редко, т.к. непродуктивно формировать мужские цветки (они не дают семян).
Гинодиэцичные растения встречается часто. В основном она характерна для растений класса Двудольные, выявлена у 613 видов. Также наблюдается связь между гинодиэцией и энтомофилией.
С морфолгической точки зрения женские цветки мельче по размерам, чем обоеполые. У женских цветков рыльце и столбик более развиты, часто больше семязачатков. Рыльце созревает раньше, на стадии бутона. Но иногда женские цветки дают меньше нектара, вплоть до отсутствия нектарников.
Трехдомные растения. Триэцичные растения встречаются редко. Соотношение половых форм может быть разным. Чаще в популяции преобладают женские и мужские, меньше обоеполых растений. Первыми зацветают женские растения, затем мужские, далее обоеполые.