ГЕТЕРОФІЛІЯ У РОСЛИН - О.М. НЕДУХА - 2011
РОЗДІЛ ПЕРШИЙ. МОРФОЛОГІЧНА ТА СТРУКТУРНО-ФУНКЦІОНАЛЬНА ХАРАКТЕРИСТИКИ ВЕГЕТАТИВНИХ ОРГАНІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
1.7. ЕПІДЕРМІС ЛИСТКІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
1.7.5. Структура поверхні листків гетерофільних рослин
1.7.5.2. Sagittaria sagittifolia
Надводні листки однієї особини S. sagittifolia - стрілолиста звичайного у фазі бутонізації мали різну форму (рис. 1.3.3, а-в; див. підрозділ 1.3 і вставку IV). Кожна рослина, яку брали для дослідження мали надводні листки двох форм (стрілоподібну й продовговату), підводні листки були сильно видовжені, мали лінійнійну форму. Кількість підводних листків у різних рослин коливалась від п'яти до девяти, надводних листків - від трьох до десяти.
Надводні листки. Стрілоподібні листки. Дослідження ультраструктури верхньої та нижньої поверхонь стрілоподібних листків показало, що листки цієї форми належать до амфістоматичного типу (рис. 1.7.5.2.1, а, в). Спільні ознаки для обох епідерм: тип продихів (парацитний), їхня щільність, форма продихів (овальновидовжена), наявність кутикуляр- ного гребеня по периметру всіх клітин і наявність воскового нальоту на зовнішній поверхні кожної клітини (рис. 1.7.5.2.1, б, г). Відмінності в структурних ознаках верхнього та нижнього епідермісів були виявлені в розмірах довгої осі епідермальних клітин і горбків воскового нальоту. Клітини нижнього епідермісу були довші порівняно з верхніми, а розмір воскових формувань був меншим на нижній поверхні (табл. 1.7.5.2.1). Воскові горбки - овальної чи округлої форми, середній розмір яких становить 1,49 ± 0,13 мкм, щільність воскового нальоту на одну клітину - 30,7 ± 2,4. Продиховий індекс на верхній стороні листка становив 13,6%, на нижній - 16%.
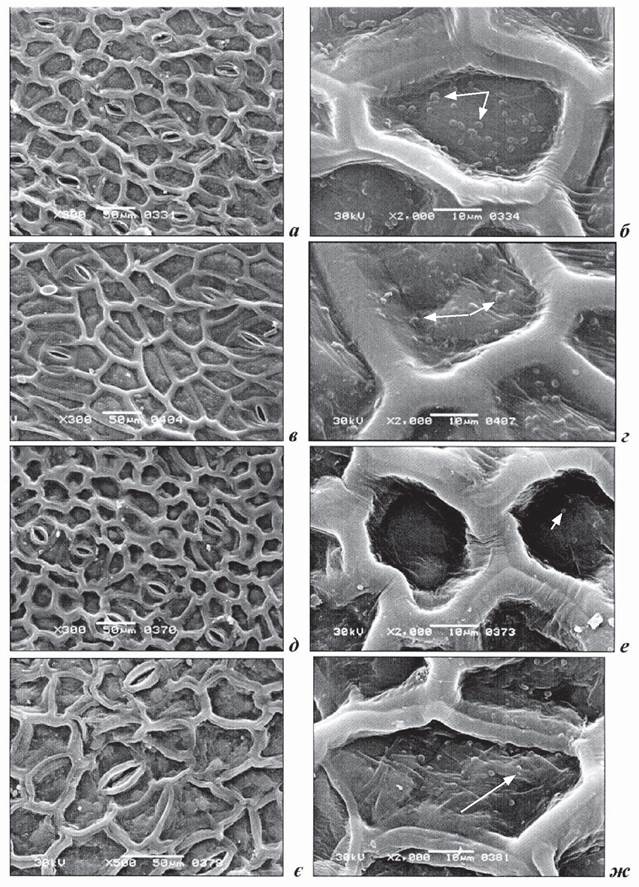
Рис. 1.7.5.2.1, Структура верхньої (а, б, д, е) та нижньої (в, г, є, ж) поверхні надводних листків Sagittaria sagittifolia з різною формою пластинок: a-г - листок з стрілоподібною пластинкою; д-ж - листок з продовгуватою пластинкою.
Восковий наліт вказано стрілками (б, г, е, ж).
Листки продовгуватої форми. Дослідження ультраструктури верхньої та нижньої поверхонь продовгуватих листків показало, що ці листки, як і листки стрілоподібної форми належать до амфістоматичного типу (рис. 1.7.5.2.1, д, є). Продихи на обох епідермах парацитного типу. На верхній епідермі вони видовженоовальні, розташовуються частіше вздовж пластинки, іноді - хаотично, середня кількість продихів на 1 мм2 нараховує 74 ± 3 шт. Контури антиклінальних оболонок основних епідермальних клітин верхньої епідерми майже прямі, форма клітин різна: від округлої чи овальної до п'ятикутної (рис. 1.7.5.2, е, ж), вкриті високим кутикулярним гребенем заввишки 5,4 ± 0,2 мкм. Поверхня частини клітин (~ 43%) епідермісу вкрита восковим нальотом овальної або округлої форми. Продиховий індекс на верхній поверхні становить 9,3%. Продихи на нижній епідермі поздовжньо-овальні, продихові щілини орієнтовані майже в одному напрямку. Показники структури клітин верхньої й нижньої поверхні епідерми продовгуватих листків наведені в табл. 1.7.5.2.1.
ТАБЛИЦЯ 1.6.5.2.1. Показники структури клітин верхньої та нижньої епідерм надводних і підводних листків Sagittaria sagittifolia за даними скануючої електронної мікроскопії
Показник |
Тип листка та його форма |
||
Надводні листки |
Підводні листки, лінійні |
||
стрілоподібні |
продовгуваті |
||
Верхня епідерма Продихи: тип щільність на 1 мм2 довга вісь, мкм коротка вісь, мкм |
Парацитний 85 ± 6 42,8 ± 3,1 20,2 ± 4,4 |
Парацитний 74 ± 3 33,4 ± 2,7 23,8 ± 3,9 |
Відсутні |
Епідермальні клітини: довга вісь, мкм коротка вісь, мкм |
45,5 ± 3,9 35,0 ± 4,1 |
44,3 ± 5,7 30,9 ± 2,7 |
109 ± 7,4* 45 ± 1,7* |
Висота кутикулярних гребенів по периметру основних епідермальних клітин, мкм |
5,5 ± 0,7 |
5,4 ± 0,2 |
0,82 ± 0,2* |
Кількість воскових горбків на клітину епідерми |
30,7 ± 5,1 |
6,2 ± 0,5 |
Не виявлено |
Процент клітин із воском, % |
100 |
43 ± 4,5 |
|
Діаметр воскових горбків, мкм |
1,46 ± 0,13 |
1,0 ± 0,04 |
|
Нижня епідерма Продихи: тип щільність 1 мм2 довга вісь, мкм коротка вісь, мкм |
Парацитний 79 ± 55 39,3 ± 3,1 20,0 ± 2,7 |
Парацитний 65 ± 4 41,8 ± 3,1 23,2 ± 2,7 |
Відсутні |
Епідермальні клітини: довга вісь, мкм коротка вісь, мкм |
62,0 ± 3,9 36,3 ± 5,1 |
62,3 ± 3,1 34,7 ± 2,7 |
106 ± 6,5* 42 ± 2,9 |
Висота кутикулярних гребенів по периметру основних епідермальних клітин, мкм |
4,9 ± 0,3 |
5,4 ± 0,17 |
1,70 ± 0,14* |
Кількість воскових горбиків на клітину епідерми |
29,3 ± 5,1 |
13,8 ± 2,75 |
Не виявлено |
Процент клітин із воском, % |
100 |
24 ± 3 |
|
Діаметр воскових горбків, мкм |
1,24 ± 0,1 |
0,8 ± 0,1 |
_ |
Примітка: * Р ≤ 0,05 (При порівнянні ознак надводних і підводних листків)
Контури антиклінальних оболонок епідермальних клітин нижньої епідерми прямі, форма клітин різна як і у верхній епідермі: від округлої або овальної до п'ятикутної, вкриті високим кутикулярним гребенем. Поверхня епідермальних клітинах (до 24 %) вкрита восковим нальотом. Воскові горбики овальної або округлої форми. Тобто, в цих листків достовірно менше число клітин вкрите восковим нальотом порівняно з стрілоподібними листками. Та й кількість воскових горбиків менша вдвічі, порівняно з клітинами епідерми стрілоподібних листків. Продиховий індекс на нижній поверхні становить 10,9 %.
Підводні листки. Структура верхньої й нижньої поверхонь епідермісу підводних листків стрілолиста звичайного відрізнялася від такої надводних листків, зокрема, відсутністю на обох поверхнях продихів. Клітини мали видовжену форму (рис. 1.7.5.2.2), їхній розмір по довгій осі був більшим, ніж у надводних листків. Контури клітин оточені дуже тонким обідком (гребенем) кутикули; висота й ширина кутикулярного гребеня були утричі менші, ніж у надводних листків (табл. 1.7.5.2.1). Крім того, на поверхні підводних листків також відсутній і восковий наліт. Часто до поверхні клітин прикріплені водорості.
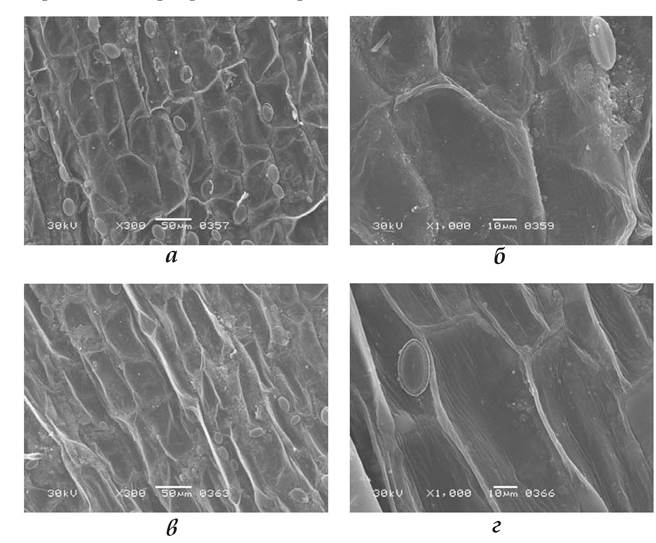
Рис. 1.7.5.2.2. Структура верхньої (а, б) і нижньої (в, г)поверхонь підводних листків Sagittaria sagittifolia.
На епідермі продихи відсутні; одноклітинні водорості прикріплені до верхньої та нижньої епідерм
Отже, порівняльний аналіз структури епідерми підводних і надводних листків стрілолиста показав, що в надводних листках незалежно від форми їхньої пластинки спостерігається наявність воскового нальоту; кількість якого на одну клітину була більша у стрілоподібних пластинках порівняно з продовгуватими. Відсоток клітин із восковим нальотом у продовгуватих пластинках був меншим, ніж у стрілоподібних: в 2,5 раза на верхній епідермі та у чотири рази - на нижній.
Причинами меншого вмісту воскового нальоту у продовгуватих листків у порівнянні із стріловидними листками можуть бути як екзогенні, так і ендогенні фактори. Можливо, що цей феномен є адаптивною ознакою на зниження дії прямого сонячного освітлення на надводні листки продовгуватої форми, оскільки стрілоподібні листки піднімаються вище над водною поверхнею, ніж продовгуваті.
Крім цього, в надводних продовгуватих листків продиховий індекс на обох поверхнях епідерми був нижчий, ніж у надводних стрілоподібних листках. Отже, у стрілоподібних листків, які піднімаються вище над поверхнею води, ніж продовгуваті листки, продиховий індекс підвищений аналогічно, як у листків суходільних рослин (Вовк, 1984).
Відомо, що віск, який утворюється на зовнішній стороні клітин епідерми, гальмує транспірацію та відбиває ультрафіолетові промені світла (Kalattukudy, 1996; Kerstiens, 1996). У стрілолиста звичайного воску було значно більше у стрілоподібних листках порівняно із продовгуватими надводними листками. Враховуючи вищевідмічені дані літератури та отримані нами результати, можна припустити, що у продовгуватих листків стрілолиста, що знаходяться ближче до водної поверхні, ніж стрілоподібні листки, кутикулярна транспірація значно інтенсивніша, ніж у стрілоподібних листках. Наявність воскового нальоту випуклої форми на епідермі двох типів надводних листків стрілолиста, очевидно, сприяє збереженню водного статусу та регуляції поглинання певної частини фотонів світла воском (Лансберг, 1986).
Механізм збільшення кількості та щільності воскового нальоту на поверхні стрілоподібних надводних листків можна також пояснити й дією ендогенних факторів. Відомо, що в епідермальних клітинах, збагачених воском, зокрема, в листках і стеблах суходільних рослин, особливо у листках сукулентів, виявлені ліпід транспортуючі білки, які беруть участь у транспортуванні та секреції фосфоліпідів у периплазматичний простір (Clark et al., 1992; Clark, Bohnert 1993; Clark, Bohnert 1999). Нещодавно були ідентифіковані гени ( Ltp, LTPs та WAX9), які відповідають за транскрипцію цих ліпід транспортуючих білків. У разі експресії відповідних генів у клітинах епідермісу виявлено ефект накопичення відповідної мРНК (Clark, Bohnert 1993; 1999). Як відбувається регуляція синтезу воску в епідермі поки невідомо. Ми можемо лише припустити, що відмінності по наявності воскового нальоту в надводних листках стрілолиста та його відсутності в епідермі підводних листків, можуть бути наслідком генетичної пластичності, в наслідок якої відбувається інгібування синтезу попередників воску (С12-, С14- і С16-ω-гідроксижирних кислот) і віск-синтетази (fatty acyl-coenzyme A: fatty alcogol acyltransferase), субстратами для якої є інтегральні мембранні білки (Hauke, Schreiber, 1998; Lardizabal et al.., 2000).