ГЕТЕРОФІЛІЯ У РОСЛИН - О.М. НЕДУХА - 2011
РОЗДІЛ ПЕРШИЙ. МОРФОЛОГІЧНА ТА СТРУКТУРНО-ФУНКЦІОНАЛЬНА ХАРАКТЕРИСТИКИ ВЕГЕТАТИВНИХ ОРГАНІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
1.2. ПОХОДЖЕННЯ ГЕТЕРОФІЛІЇ
Уявлення про виникнення гетерофілії у рослин неоднозначні. Є кілька точок зору щодо походження гетерофілії: еволюційне та екологічно- генетичне (це так звана пластична гіпотеза), які не є протилежними, а розглядаються на різних рівнях організації (філогенетичному, онтогенетичному, тканинному, клітинному і ін.). Розглянемо кожну з існуючих гіпотез.
Еволюційне походження гетерофілії базується на принципі, що гетерофілія у рослин виникла внаслідок конвергентної або паралельної еволюції, як відповідь на водне оточення (Bradshaw, 1965). Згідно з даними інших авторів (Cook, Johnson, 1968; Smith, Hake, 1992) дуже мало відомо про еволюційне походження гетерофільних рослин, тобто про умови, в яких виникає гетерофілійний габітус (Wells, Pigliucci, 2000). Вважають, що більшість водних таксонів належить до однодольних порівняно лише з кількома представниками дводольних гідрофітів. Повернення до водного існування у дводольних не виявлено (Wells, Pigliucci, 2000). За гіпотезою Дж. Хатчінсона пластичні гетерофільні види виникли з непластичних гетерофільних однодольних суходільних предків (Hutchinson, 1975; Goliber, Feldman, 1990) у процесі еволюції під час змін оточуючого середовища та клімату. І як вважає С.Л. Велс та М. Піглусі (Wells, Pigliucci, 2000), саме на це вказують численні експериментальні роботи, коли гідрофіти переносили в акваріуми, в яких створювалися умови виходу пагона з водного середовища.
А.Д. Бредшоу, вивчаючи еволюційне значення пластичності для рослин, відзначив, що численні родичі гетерофільних таксонів стали «слабкішими» до експресії пластичності за зміни форми листків. Він показав, що не всі види одного й того самого роду є гетерофільними. Так, серед однодольних роду Potamogeton є три гетерофільні види: Potamogetonnodous, P. natans і P. polygonifolius, тимчасом як інші види цього роду, зокрема P. lucens і P. perfoliatus, не є гетерофільними (Bradshaw, 1965). Подібні відмінності характерні й для видів роду Juncuss. Так, J. heterophylla - гетеро- фільний вид, а J. obtusiflorus - негетерофільний. Серед дводольних також не всі види певної родини мають ознаки гетерофілії:
1) Callitriche intermedia - гетерофільний вид,
C. heterophylla - гетерофільний вид,
C. stagnalis - негетерофільний вид,
2) Proserpinaca intermedia - гетерофільний вид,
P. palustris - гетерофільний вид,
P. pectinata - негетерофільний вид,
3) Ranunculus aquatilis - гетерофільний вид,
R. flabellaris - гетерофільний вид,
R. flammula - гетерофільний вид,
R. hederaceus - негетерофільний вид (Bradshaw, 1965).
Наявність певних філогенетичних зв'язків у гетерофільних і гомо- фільних видів [heterophyllous and homophyllous species (Iida et al., 2009)] роду Potamogeton також відмітили Д. Лес і Д. Шрейдер (Les, Sherider, 1990). Ці дослідники, вивчаючи сімнадцять Північно-Американських видів роду Potamogeton (P. foliosus, P. pectinatus, P. perfoliatus, P. pusillus, P. richardsonii, P. robbinsii, P. crispus, P. vaseyi, P. zosterifolius, P. alpinus, P. amplifolius, P. epihydrus, P. gramineus, P. illinoensis, P. natans, P. nodosus, P. spirillus), серед яких були гетерофільні й гомофільні види (перші дев'ять видів) (рис. 1.2.1), показали, що надводні й підводні листки гетерофільних видів рдесника сильно відрізняються за синтезом флавоноїдів. Тоді як вміст флавоноїдів у надводних листках гетерофільних видів був подібний до вмісту у гомофільних видів рдеста.
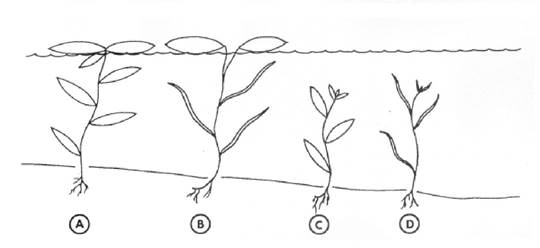
Рис. 1.2.1. Основні морфологічні типи Potamogeton.
А - гомофільний вид, у якого підводні й плаваючі листки за формою подібні (ланцетоподібні); B - гетерофільний вид, у якого підводні лінійні листки відрізняються від ланцетоподібних плаваючих листків; C - вид гомофільний, із широкими ланцетоподібними видовженими підводними листками; D - вид гомофільний, із лінійними видовженими підводними листками (Les, Sherider, 1990)
Дослідники визначили наявність у плаваючих листках гетерофільних видів Potamogeton наступних фенольних речовин: лютеоліну, 7-ОСН3 лю- теноліну, апігенін 6-С-глюкозиду, арігенін 7-О-глюкоуроніду, лютеолін 7-О-глюкозиду, лютеолін 6-С-глюкозиду, хризоеріол 6-С-глюкозиду, лютеолін 7-О-глюкоуроніду, апігеніну й хризоеріолу. У підводних листках перші шість глюкозидів були відсутні, тоді як сьомий глюкозид - лютеолін 7-О-глюкоуронід, виявили як у підводних листках, так і в черешках і пластинках надводних листків. Авторами показано, що в гомофільних видів, для яких характерні широкі підводні листки, набір фенолів є ближчим до такого в надводних листків гетерофільних видів. Аналіз цих речовин показав наявність філогенетичних зв'язків у цього роду незалежно від морфологічних і хромосомних результатів. Глікофлавони, які виявлені в надводних листках, очевидно, є не тільки фільтрами ультрафіолетової радіації (McClure, 1975), а й метаболітами, синтез яких можна віднести до адаптивного механізму на функціональному рівні при прояві гетерофілії. Інгібування синтезу глікофлавонів підводними листками гетерофільних видів Potamogeton, а також і лінійними листками в гомо- фільних видів Potamogeton, очевидно, є наслідком зниження УФ радіації у воді, яка абсорбує її промені. На основі визначення й розподілу дев'яти вище зазначених флавоноїдів автори прийшли до висновку, що гетерофільні види це цілісна група рослин; тоді як гомофільні види (із широкими овальновидовженими листками) за хімічними ознаками більш подібні до плаваючих (овальновидовжених) листків гетерофільних листків, що було представлено ними схематично (рис. 1.2.2) (Les, Sherider, 1990).
Крім цього, автори представили еволюційні зв'язки між різними морфологічними групами видів роду Potamogeton (рис. 1.2.3). Ці дані забезпечують пряму підтримку ранніх гіпотез, згідно яких флавоноїди відіграли суттєву роль в еволюційному розвитку при завоюванні рослинами суходільного габітусу (Miller, 1974; McClure, 1975).
Згідно С. Iida і співав. (2004, 2006), ген хлоропласта rbcL, який кодує каталітичну субодиницю Рубіско, залучений у позитивний відбір більшості наземних рослин. Дослідження адаптивного еволюційного процесу, який зв'язаний із гетерофілією чи гомофілією, дало можливість проаналізувати позитивний відбір послідовностей гена rbcL у екологічно різних водних рослин Японії, а саме у видів роду Potamogeton, які є досить різноманітними за морфологією й ростовими показниками (Wiegleb, 1988). Автори прийшли до висновку, що гомофілія у видів цього роду є спадковою, а гетерофілія, яка передається спадково, можливо, існує, як паралельна еволюція (Iida et al., 2004; 2006). Продовжуючи вивчати взаємодію генотипу й оточуючого середовища в прояві гетерофілії у численних видів роду Potamogeton, серед яких були і природні гібриди, що зростали в Японії, ці дослідники протестували амінокислотний склад гена rbcL у гетерофільних і гомофільних видів рдесника, використовуючи філогенетичний аналіз та аналіз кодона (Iida et al., 2009). Три гени (rbcL, atpB, petA) були послідовно секвеновані із 18 видів роду Potamogeton. Усі нуклеотидні послідовності визначали за даними GenBank. Згідно з даними попередніх праць, філогенетичні дерева, засновані на окремих хлоропластих генах, сумісні (Iida et al., 2004).
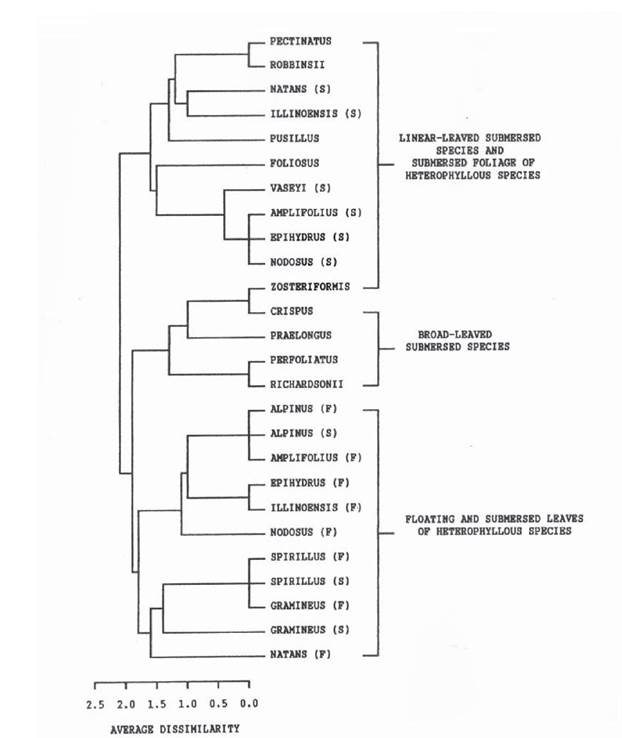
Рис. 1.2.2. UPGMA дендрограма евклівдовських відстаней, які відповідають біохімічним даним, отриманим за флавоноїдними ознаками на 17 видах роду Potamogeton. Позначення: F - дані по плаваючим листкам, S - дані по підводним листкам (Les, Sherider, 1990).
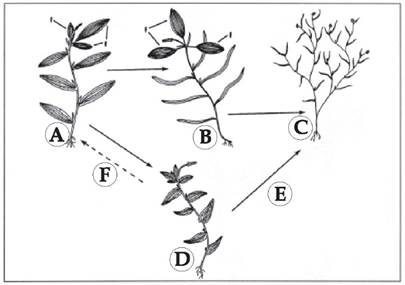
Рис. 1.2.3. Гіпотетичні зміни морфологічних і біохімічних ознак у процесі еволюції видів роду Potamogeton. А - нечітка гетерофілія й відмінності по флавоноїдам характеризують предків Potamogeton (наприклад, у P. alpines); B - морфологічна і біохімічна спеціалізація призвела до чіткої гетерофілії із редукцією флавоноїдів в підводних листках (наприклад, у P. epihydrus); C - гомофільні види із лінійними листками, в яких вміст флавоноїдів знижений (наприклад, у P. foliosus), можуть походити від ге- терофільних предків шляхом втрати плаваючих листків; D - гомофільні види із ланцетоподібними листками із різним вмістом флавоноїдів (наприклад, у P. richardsonii) можуть походити від гетерофільних предків, які втратили плаваючі листки; E - гомофільні види із лінійними листками, очевидно, виникли із гомофільних видів (з ланцетоподібними листками) шляхом морфологічної спеціалізації і зниженням синтезу флавоноїдів; F - почергово широколистяні (із ланцетоподібними листками) гомофільні види були предками гетерофільних видів шляхом морфологічної спеціалізації плаваючих і підводних листків. Позначення: f - плаваючі листки (Les, Sherider, 1990)
Дослідники (Iida et al., 2009) скомбінували дані по чотирьох генах (1349 rbcL, 1467 atpB, 918 petA, 660 trnT-trnL) i отримали філогенетичне дерево Potamogeton за хлоропластною ДНК (це - cpDNA tree; рис. 1.2.4; див. вставку І).
Головне галуження в цьому філогенетичному ДНК-дереві адекватне такому, яке це описано в попередній роботі при вивченні міжгенної заміни (підставки trnT-trnL) ( Iida et al., 2004). Японські види Potamogeton були виділені в групу Ш, із широкими підводними листками, а в групі П - були види з лінійними підводними листками. А пізніше вони були розділені на дві підгрупи П а й П б (рис. 1.2.4). Додатково було ідентифіковано нове бокове галуження, як група із кількох видів (рис. 1.2.4). Перевірку гіпотези позитивного відбору в гена rbcL перевіряли на співпаданні послідовностей у гена Potamogeton rbcL, базуючись на cpDNA-дереві. Авторами встановлено розподіл 12 змінних амінокислотних сайтів у гені RbcLs у гетерофільних і гомофільних видів із роду Potamogeton і у початкових (вихідних) видів (таблиця 1.2.1). Амінокислотні заміни при 225 і 281 залишках були ідентифіковані, як потенціальні залишки при позитивному відборі. Найбільші заміни відбувалися на термінальних галуженнях філогенетичного дерева, у порівнянні з основою (рис. 1.2.4; див. вставку І).
Заміна залишка 225 (Ile225 → Leu225, та обмін у кодоні ATT → CTT) відбувалася двічі: перший раз у підгрупі Ia, а потім - у підгрупі IIa. Тоді як залишок 281 замінювався п'ять разів: тричі відбувалися паралельні заміни Ser281→ Ala281 (TCT → GCT), і двічі Ala281→ Ser281 (GCT → RTCT). Численні амінокислотні заміни в Potamogeton відбуваються між субодиницями та біля активних сайтів. Сім амінокислот (76, 230, 245, 249, 279, 282, 309) були локалізовані на поверхні молекули RbcL. Дві з них (76 і 230) були на ітер- фейсі між RbcL і RbcS, тоді як п'ять амінокислот (245, 249, 279, 282 і 309) розміщувалися на інтерфейсі димера RbcL. Останні чотири амінокислоти (101, 225, 281 і 328) були заглиблені в молекулу. Залишок 270 локалізувався біля молекули, хоча він не повністю був у неї занурений. Тоді як залишки 270 і 328 розміщувалися в субстрат-зв'язуючих зонах і були закриті активними сайтами (Arg295, His298 і His327) (Knight et al., 1990). Тобто проведений молекулярно-еволюційний аналіз показав, що ген rbcL у видів роду Potamogeton залучений до позитивного відбору (таблиця 1.2.1).
ТАБЛИЦЯ 1.2.1. Сайти заміни амінокислот в гені RbcL у 18 видів Potamogeton і Stuckenia pectinata (Iida et al., 2009)
Вид / ознака |
Амінокислотний сайт |
|||||||||||
76 |
101 |
225 |
230 |
245 |
249 |
270 |
279 |
281 |
282 |
309 |
328 |
|
P. crispus (-a) |
S |
V |
I |
A |
G |
E |
L |
T |
S |
H |
M |
S |
P. maackianus (-a) |
S |
V |
I |
A |
G |
D |
L |
S |
A |
H |
M |
S |
P. gramineus (++) |
N |
V |
L |
G |
G |
E |
L |
S |
A |
H |
M |
S |
P. dentatus (-) |
N |
V |
L |
G |
G |
E |
L |
S |
S |
H |
M |
S |
P. distinctus (++) |
S |
V |
I |
A |
G |
E |
L |
S |
A |
H |
M |
A |
P. malaianus (++) |
S |
V |
I |
A |
A |
E |
L |
S |
A |
H |
M |
A |
P. perfoliatus (-a) |
S |
V |
I |
A |
G |
E |
I |
S |
A |
H |
M |
S |
P.alpinus (+), P. cristatus (+) |
S |
V |
I |
A |
G |
E |
L |
S |
A |
H |
M |
S |
P. fryeri (++), P. natans (++) |
S |
V |
L |
A |
G |
E |
L |
S |
A |
N |
M |
S |
P. praelongus (-), P. oxyphyllus, P. octandrus (+) |
S |
V |
I |
A |
G |
E |
L |
S |
S |
H |
M |
S |
P. compressus (-), P. obtusifolius (-), P. panormitanus (-a), P. pusillus (-) |
S |
I |
I |
A |
A |
E |
L |
S |
S |
H |
I |
S |
Stuckenia pectinata (-a) |
S |
V |
I |
A |
A |
E |
L |
S |
S |
H |
M |
S |
Структурна особливість b |
S |
V |
I |
A |
G |
E |
L |
S |
S |
Y |
M |
S |
Достовірність, ω ≥ 1c : |
I |
c |
c |
i |
D |
D |
*c |
D |
c |
D |
D |
*c |
Сайт-специфічна модель (M8) |
.51 |
.38 |
.86 |
.48 |
.85 |
.32 |
.49 |
.48 |
1.0 |
.86 |
.43 |
.38 |
Сайт- розгалужуюча модель (ω2: гетерофілія) |
.73 |
.02 |
.98 |
.74 |
.29 |
.02 |
.72 |
.03 |
.98 |
.29 |
.02 |
.68 |
Позначення: (-a) кількість зв'язків гену RbcL гомофільного виду рдесника (з підводними листками) із амінокислотними сайтами гену RbcL у шпината; (+) кількість зв'язків гену RbcLгетерофільного виду рдесника (з підводними і плаваючими листками) із амінокислотними сайтами гену RbcL у шпината;
(-) кількість зв'язків гену RbcL гетерофільного виду рдесника (з підводними, плаваючими і суходільними листками) із амінокислотними сайтами гену RbcL у шпината.
а - види, які зростали в солончаковій воді; b* - залишки, які були закриті для активних центрів.
C - залишки, які були заглиблені у внутрішню частину RbcL, D - залишки на поверхні димеру RbcL; i - залишки між поверхнями двох генів RbcL і RbcS
Із дванадцяти амінокислотних сайтів два сайти (225 і 281) привели до позитивної селекції; а заміна на сайті 281 ініціює різні філогенетичні покоління в гетерофільних і гомофільних видів (рис. 1.2.4; див. вставку І). Таким чином, було встановлено, що еволюція екологічної дивергенції (розходження) у японського виду Potamogeton залучена в молекулярну адаптацію гена rbcL (Iida et al., 2009).
Згідно з результатами досліджень M.A. Гебера та ін. (Geber et al., 1992) втрата варіацій пластичності виду в гетерогенних умовах його існування допускає невеликі генетичні варіації в ознаках. Очевидно, це є виявом певної фенотипічної пластичності як у водних, так і суходільних рослин, а не результатом стабілізації добору (Cook, Johnson, 1968; Winn, 1999). Втрата певних ознак морфологічної пластичності гомофільними видами ставить перед дослідниками ряд запитань:
— Чому це відбувається в певних видів?
— Чи рівень регуляції в рослинних популяціях гідрофітів до флуктуацій у водному середовищі, які призводять до вияву ознак гетерофілії, є вищим, ніж рівень регуляції в суходільних популяціях, які за умов затоплення зростають у воді?
— Що є показником еволюції гетерофільних таксонів, коли їхнє філогенетичне походження розглядається разом зі змінами навколишнього середовища?
Ці питання потребують відповідей, а отже об'єднання зусиль флористів, ботаніків, систематиків, екологів й генетиків для висунення чіткої концепції походження гетерофільних видів.
Екологічна теорія походження гетерофільних рослин допускає, що популяція, яка зростає за значних змінах рівня води характеризується більшою пластичністю морфології листків порівняно з популяціями, що не зазнають дії таких змін. Мінливість морфології листків зменшується при зниженні рівня води, що прямо корелює з теорією адаптивної пластичності, згідно якої структурно-функціональна пластичність виявляється лише за певних умов оточуючого середовища (Levene, 1953; Schlichting, Pigliucci, 1995; Schlichting, Pigliucci, 1998). Коли ж спостерігається втрата пластичності в популяції за більш однорідних природних умов, то незважаючи на це, у такої популяції все ж лишається здатність до прояву гетерофілії (Wells, Pigliucci, 2000).
Так, для видів роду Nuphar (Nymphaeaceae) характерне утворення перших повітряних (надводних) листків, які нагадують їхню підводну форму; це спостерігається у рослин, які зростають як при обмілінні (на суходолі), так і у воді. У видів Nuphar спочатку утворюються недозрілі листки, такими вони лишаються протягом кількох років; за зовнішнім виглядом ці листки нагадують молоді примордії, що розміщуються вздовж горизонтально розміщеного стебла (Cutter, 1957). Такі листки розвиваються кожного року на кореневищі, виявляючи непластичний онтогенез (Aydelotte, Diggle 1997; Diggle 1997, a, b; Wells, Pigliucci, 2000).
Фундаментальні відмінності між більш лабільними видами, які належать до пластичних гетерофільних гідрофітів, та видами, незалежними від зміни умов довкілля (суходіл ↔ вода), очевидно, розкриваються в концепції реверсивності (оборотності). Десятки здійснених досліджень по вивченню гетерофілії у вищих водних рослин дали змогу виявити, що їхні підводні листки, як і повітряні, формуються на одному паростку, у разі затоплення - занурені у воду листки стають подібними до підводних (Cook, 1969; Bodkin et al., 1980; Bruni et al., 1996). Це було також доведено й експериментальними роботами при зміні умов зростання гідрофітів (Goliber, Feldman 1990; Bruni et al., 1996).
Варіабельність форми листків у суходільних рослин може спостерігатися в наступних ситуаціях: 1) у разі прискореного переходу від ювенільної фази розвитку листка до зрілої; 2) у разі зміни вегетативної фази на репродуктивну; 3) коли в багаторічних рослин ранні листки формуються в попередньому сезоні, а пізніші - в наступному (Eckenwalder, 1980; Webb, 1984; Christodoulakis, 1989; Christodoulakis et al, 1990); 4) при утворенні ювенільних листків уздовж осі латерального пагона (Ashby, 1948, а, b). Зміна ж форми листків у водних рослин залежить, як від зміни екзогенних (вода ↔ повітря, вміст газів, освітленість, температура, фотоперіод), так і від ендогенних факторів.
Різна форма листків у гетерофільних рослин - це вияв гетеробластного розвитку, коли ювенільні стебла й пагони відрізняється від стебел і пагонів, що закінчили свій ріст за кількома критеріями (Allsopp, 1967). Ювенільна фаза листків є меншою і простішою, ніж доросла, а, крім того, може відрізнятися й за морфоструктурними ознаками. Наприклад, ювенільні листки кукурудзи значно коротші й вужчі, ніж дорослі, але вони мають епікутикулярний віск і волоски, які відсутні в дорослих листків (Poethig, 1997). Хоча ювенільні листки спочатку затримуються в рості на відміну від розвинутих листків, але порівняння розвитку двох форм листків у кількох видів показало, що примордій ювенільного листка звичайно менший і його морфологія відрізняється від примордію дорослого листа. Перехід від ювенільної до дорослої фази розвитку стебла також маркується трансформацією апікальної меристеми пагона (Abbe et al., 1941; Stein, Fosket, 1969; Kaplan, 1973; Franck, 1976; Greyson et al.,1982). Проте кореляція між розмірами меристеми та листка спостерігається не завжди й не є універсальною в разі вияву гетеробластного розвитку. Наприклад, у Muehlenbeckia plafyclados перехід від ювенільної до зрілої фази розвитку листка супроводжується збільшенням зони меристеми, тоді як зрілі листки мають малу зону меристеми (Bruck, Kaplan, 1980).
На органному та клітинному рівнях, згідно з теоретичними уявленнями Е. Синноти (Sinnot, 1940; Синнот, 1963), форма листкової пластинки є кінцевим продуктом зміни трьох компонентів: форми примордію, кількості та розподілу клітинних поділів і кількості та розподілу клітин, які розтягуються. Кожен із цих компонентів може залежати як від спадковості, так і від фізичних факторів оточуючого середовища (Ashby, 1948, a; 1948, b). Наприклад, зв'язок між цими трьома компонентами під час розвитку листків у Gossypium sp., який мав мутацію okra, описаний ще у 1941 р. (Hamrnond, 1941). Листки мутантів були довшими й мали глибші пазухи між дольками (лопатями) листків порівняно із такими в листках дикого типу Gossypium. Крім того, у мутантів листкові примордії були значно вужчими, ніж у листків дикого типу, їхні листки також були довшими, що було спричинено збільшенням кількості клітинних поділів повздовж листка. Три компоненти, які описав Е. Синнот, не завжди взаємодіють зі зміною форм листка. На листках Cyamopsis tetragonoloba, що розвивалися, виявлені зміни клітинного поділу та розтягування по периферії пластинки, що корелювали зі збільшенням або зниженням листкових дольок у трилистих і листках з цілісною пластинкою (Sparks, Postlethwait, 1967, a; 1967, b). Підкреслимо, що примордії цілісного та складного листків не відрізнялися на ранній стадії розвитку. У трилистих листках клітинний поділ у «кишенях» базипетального краю відрізнявся від такого в цілісних листках, а саме, поділ відставав, що призводило до утворення глибоких листкових дольок; у цілісного ж листка клітинний поділ не припинявся, і утворення заокруглених дольок не відбувалося, а утворювався листок з цілісною пластинкою.
На противагу цьому, в іншого виду, Callitriche intermedia, який є гетерофілом (утворює овальні листки над водою та видовжені паскоподібні листки у воді), встановлено, що форма листка значно менше відрізнялась при дослідженні поділу та розтягування клітин; дослідники спостерігали формування лише видовжених дольок у разі утворення паскоподібних листків (Deschamp, Cook, 1983).
Регуляція форми листка може відбуватись й на рівні однієї клітини або органа, або ж шляхом комплексних процесів на обох рівнях - клітини та органа. Ідея, стосовно того, що органогенез керується на рівні клітини, подана в «клітинній теорії» - за аналогією з клітинною теорією, розробленою для тваринних організмів (Jacobs, 1997). З погляду на таку перспективу, форма органів рослин залежить від меристематичних клітин, які діляться, а потім розтягуються до утворення кінцевої морфоструктури.
На противагу цій теорії є «теорія організму» (Kaplan, Hagemann, 1991; Kaplan, 1992), згідно з якою розтягування органу регулюється на надклітинному рівні, можливо, завдяки вияву фізичного натягу через примордії або ж через весь орган, який формується (Francis, 1998).
Порівняння морфогенезу звужених сім'ядолей мутанта Arabidopsis thaliana L. Heynh із широкими сім'ядолями дикого типу цієї рослини, показало, що форма органа обумовлена лише регуляцією клітинного розтягування, а не поділом клітин (Tsukaya et al., 1994). Очевидно є сенс як у першій, так і в другій теорії (клітинній та організменній); згідно з чим у меристемі відбувається певне роз'єднання поділу та розтягування клітин. Зауважимо, що головна роль відводиться клітинам епідермісу, які утворюються з першого шару апікальної меристеми пагона-стебла. Дані, отримані на мутантах, подібних до мутантів okra в рослин роду Gossypium, показали, що саме зміна форми листка відбувається тоді, коли експресія має місце лише в першому епідермальному шарі (Hamrnond, 1941; Dolan, Poethig, 1991; Poethig, 1997). Встановлено, що на найбільш ранніх етапах розвитку, примордії підводних і надводних листків (у Callitriche heterophylla та Proserpinaca palustris) не відрізняються доти, поки їхній розмір по довгій осі не досягне 350-400 мкм (Burns, 2005; Deschamp, Cook. 1983). І лише після цього морфологія повітряних і водних листків корелює зі збільшенням частоти клітинного поділу в епідермісі: у водних листків клітинний поділ був частішим у лобових дольках порівняно з пазушними зонами, тимчасом як у повітряних листках подібна кореляція клітинного поділу по поверхні листка не відмічена (Schmidt, Millington, 1968; Deschamp, Cook. 1983).
Подальше вивчення походження гетерофілії у водних і суходільних рослин може бути направлено на визначення критичних факторів, які обумовлюють прояв гетерофілії в рослині, і механізмів адаптації її онтогенезу у варіабільному середовищі, починаючи від сприйняття зовнішнього сигналу, його передачі, й відповіді рослини на сигнал на всіх рівнях організації (починаючи з молекулярного і клітинного й закінчуючи як видовим, так і популяційним), подібно до такого, який був запропонований видатним ботаніком Єлізаветою Кордюм при вивченні адаптації рослин до дії зовнішніх чинників (Кордюм, 2003, а).