СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 2. ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
Клеточная оболочка
Наличие клеточной оболочки более чем все другие признаки отличает растительные клетки от животных. Клеточная оболочка ограничивает размер протопласта и предотвращает его разрыв за счет поглощения воды вакуолью.
Клеточную оболочку не так давно считали внешним, неактивным продуктом протопласта. Теперь установлено, что она имеет специфические функции, которые весьма важны не только для клетки и ткани, в которой она находится, но и для всего растения. Клеточные оболочки играют существенную роль в поглощении, транспорте и выделении веществ, а кроме того, в них может быть сосредоточена лизосомальная, или переваривающая, активность.
Компоненты клеточной оболочки
Наиболее типичным компонентом клеточной оболочки является целлюлоза, которая в значительной степени определяет ее архитектуру. Молекулы целлюлозы состоят из повторяющихся молекул глюкозы, соединенных конец к концу (см. гл. 3). Длинные тонкие молекулы целлюлозы объединены в микрофибриллы толщиной примерно от 10 до 25 нм (рис. 2-28). Целлюлоза обладает кристаллическими свойствами (рис. 2-29) благодаря упорядоченному расположению молекул в отдельных участках микрофибрилл — мицеллах (рис. 2-30). Микрофибриллы перевиваются и образуют тонкие нити, которые в свою очередь могут обматываться одна вокруг другой, как пряди в канате. Каждый такой «канат», или макрофибрилла, имеет толщину около 0,5 мкм и может достигать в длину 4 мкм. Макрофибриллы так же прочны, как равная по толщине стальная проволока.
Рис. 2-28. Поверхность оболочки клетки водоросли Chaetomorpha. Видны целлюлозные микрофибриллы, каждая из которых состоит из сотен молекул целлюлозы

Рис. 2-29. Каменистые клетки (склереиды) из мякоти плода груши (Руrus communis), видимые в поляризованном свете. Скопления этих клеток обусловливают зернистую текстуру плода. Каменистые клетки имеют очень толстые вторичные оболочки, пересеченные многочисленными простыми порами (они видны как линии на оболочках). В поляризованном свете оболочки выглядят светлыми из-за кристаллических свойств их основного компонента — целлюлозы
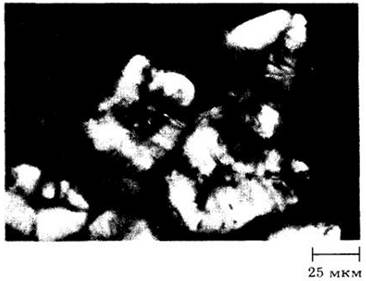
Рис. 2-30. Детальная структура клеточной оболочки. A. Участок оболочки, на котором показаны срединная пластинка, первичная оболочка и три слоя вторичной оболочки. Целлюлоза, основной компонент клеточной оболочки, представлена системой фибрилл разного размера. Б. Самые крупные фибриллы, макрофибриллы, можно увидеть под световым микроскопом. В. С помощью электронного микроскопа можно различить, что макрофибриллы состоят из микрофибрилл толщиной около 100 А Г. Участки микрофибрилл, мицеллы, имеют упорядоченное строение и придают оболочке кристаллические свойства. Д. Фрагмент мицеллы. Показаны участки похожих на цепочки молекул целлюлозы, образующих решетку

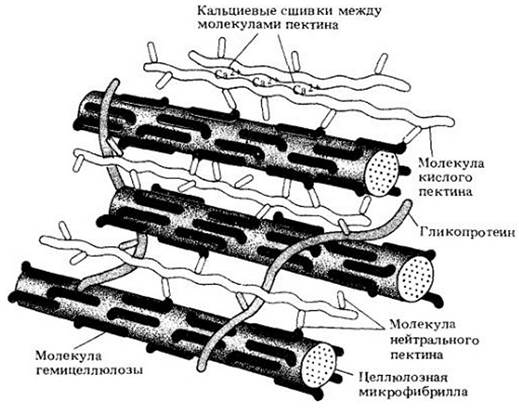
Целлюлозный каркас клеточной оболочки заполнен переплетающимися с ним нецеллюлозными молекулами матрикса. В его состав входят полисахариды, называемые гемицеллюлозами, и пектиновые вещества, или пектины, химически очень близкие к гемицеллюлозам (рис. 2-31).
Рис. 2-31. Схема возможного соединения микрофибрилл и компонентов матрикса (пектины, гемицеллюлозы и гликопротеины) в первичной клеточной оболочке. Молекулы гемицеллюлозы прикреплены к поверхности целлюлозных микрофибрилл водородными связями. Некоторые молекулы гемицеллюлозы связаны с молекулами кислого пектина через молекулы нейтрального пектина. Гликопротеины, вероятно, присоединены к молекулам пектина
Другой компонент оболочки — лигнин — является самым распространенным после целлюлозы полимером растительных клеток. Лигнин увеличивает жесткость оболочки и обычно содержится в клетках, выполняющих опорную, или механическую, функцию.
Кутин, суберин и воска — жировые вещества, которые обычно откладываются в оболочках защитных тканей растения. Кутин, например, содержится в клеточных оболочках эпидермы, а суберин — вторичной защитной ткани, пробки. Оба вещества встречаются в комбинации с восками и предотвращают чрезмерную потерю воды растением (см. с. 55).
Слои клеточной оболочки
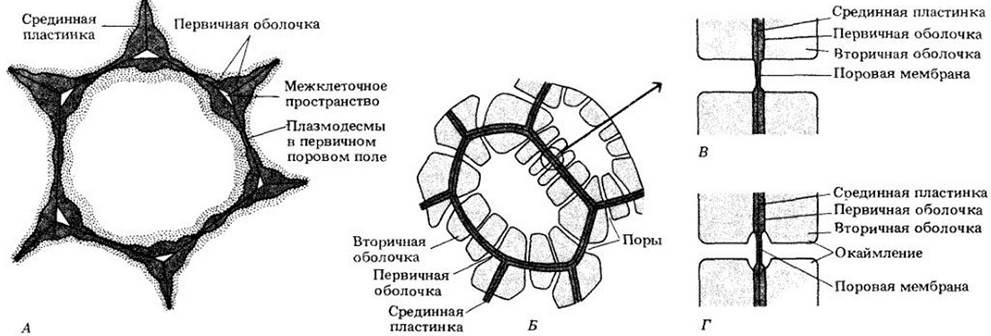
Толщина оболочки растительных клеток варьирует в широких пределах в зависимости от роли клеток в структуре растения и возраста индивидуальной клетки. Исследования, проведенные с помощью электронного микроскопа, поляризованного света и рентгеновских лучей, показали наличие двух слоев в оболочке растительных клеток: срединной пластинки (называемой также межклеточным веществом) и первичной клеточной оболочки. Кроме того, многие клетки откладывают еще один слой — вторичную клеточную оболочку. Срединная пластинка располагается между первичными оболочками соседних клеток. Вторичная оболочка, если она есть, откладывается протопластом клетки на внутреннюю поверхность первичной клеточной оболочки (рис. 2-32).
Рис. 2-32. Первичные поровые поля, поры и плазмодесмы. А. Паренхимная клетка с первичной оболочкой и первичными поровыми полями — тонкими участками оболочки. Видно, что плазмодесмы обычно пронизывают оболочку в первичных поровых полях. Б. Клетки со вторичными оболочками и многочисленными простыми порами. В. Пара простых пор. Г. Пара окаймленных пор
Срединная пластинка
Срединная пластинка состоит в основном из пектиновых веществ. Как правило, трудно различить срединную пластинку и первичную оболочку, особенно в клетках с хорошо развитой, толстой вторичной оболочкой. Когда клеточная оболочка лигнифицируется, этот процесс начинается в срединной пластинке, затем распространяется на первичную и, наконец, на вторичную оболочку.
Первичная клеточная оболочка
Слой целлюлозной оболочки, который откладывается до начала или во время роста клетки, называется первичной клеточной оболочкой. Помимо целлюлозы, гемицеллюлоз и пектина первичные оболочки содержат гликопротеин (рис. 2-31). Первичные оболочки могут и лигнифицироваться. Пектиновый компонент придает пластичность, которая позволяет первичной оболочке растягиваться по мере удлинения корня, стебля или листа.
Активно делящиеся клетки (как и большинство зрелых клеток, вовлеченных в процессы фотосинтеза, дыхания и секреции) имеют только первичные оболочки. Такие клетки — т. е. клетки с первичной оболочкой и живым протопластом — способны утрачивать характерную форму, делиться и дифференцироваться в новый тип клеток. По этой причине именно они участвуют в заживлении ран и регенерации у растений.
Первичные клеточные оболочки не одинаковы по толщине на всем своем протяжении, а имеют тонкие участки, которые называются первичными поровыми полями (рис. 2-32). Тяжи цитоплазмы, или плазмодесмы, соединяющие протопласты соседних клеток, обычно, проходят через первичные поровые поля.
Вторичная клеточная оболочка
Хотя многие растительные клетки имеют только первичную оболочку, у некоторых протопласт откладывает к центру клетки вторичную оболочку. Обычно это происходит после того, как клетка прекращает расти и площадь первичной оболочки более не увеличивается. Отчасти по этой причине вторичная оболочка по структуре отличается от первичной. Вторичные оболочки особенно нужны специализированным клеткам, укрепляющим растение и проводящим воду. Протопласт этих клеток, как правило, отмирает после отложения вторичной оболочки. Во вторичных оболочках больше целлюлозы, чем в первичных, а пектиновые вещества и гликопротеины в них отсутствуют. Вторичная оболочка поэтому жестка и нелегко растягивается, ее матрикс состоит из гемицеллюлозы.
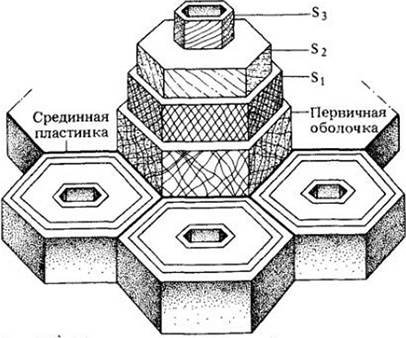
Как правило, во вторичной оболочке можно выделить три слоя — наружный, средний и внутренний, которые обозначаются соответственно S1, S2 и S3 (рис. 2-33). Эти слои отличаются друг от друга по ориентации целлюлозных микрофибрилл. Такие слоистые оболочки характерны, например, для клеток вторичной ксилемы, или древесины. Слоистая структура вторичных оболочек, — подобная той, что видна в фанере, — значительно увеличивает их прочность. Микрофибриллы целлюлозы во вторичной оболочке откладываются плотнее, чем в первичной. Лигнин — обычный компонент вторичных оболочек клеток древесины.
Рис. 2-33. Схема расположения микрофибрилл в первичной оболочке и в трех слоях (S1, S2, S3) вторичной клеточной оболочки
Вторичная оболочка не откладывается на первичных поровых полях. Соответственно во вторичной оболочке образуются характерные углубления, или поры (рис. 2-32). В некоторых случаях поры образуются в местах, где нет первичных поровых полей. Поры в оболочках контактирующих клеток расположены напротив друг друга. Срединная пластинка и две первичные оболочки между двумя порами называются поровой мембраной. Две лежащие друг против друга поры и поровая мембрана образуют пару пор. В клетках, имеющих вторичные оболочки, существуют два основных типа пор: простые и окаймленные. В окаймленных порах вторичная оболочка нависает над полостью поры. В простых порах этого нет.
Рост клеточной оболочки
По мере роста клетки увеличиваются и толщина, и площадь клеточной оболочки. Растяжение оболочки — это сложный процесс, находящийся под строгим биохимическим контролем протопласта; он регулируется гормоном ауксином (см. гл. 24) и требует усиления синтеза белка, дыхания и поглощения воды клеткой. Большая часть новых микрофибрилл располагается поверх образовавшихся ранее (слой на слой), однако некоторые могут включаться и в уже существующую структуру.
В клетках, растущих во всех направлениях более или менее равномерно, отложение микрофибрилл носит случайный характер, и они образуют неправильную сеть. Такие клетки обнаружены в сердцевине стебля, запасающих тканях и при культивировании растительных клеток in vitro. Напротив, в удлиняющихся клетках микрофибриллы боковых оболочек откладываются под прямым углом к оси удлинения. По мере увеличения поверхности оболочки ориентация микрофибрилл наружного слоя становится почти продольной, или параллельной длинной оси клетки.
Вещества матрикса — пектины и гемицеллюлозы — так же, как и гликопротеины, переносятся к оболочке в пузырьках диктиосом. При этом пектины более характерны для растущих клеток, а гемицеллюлозы преобладают в клетках, прекративших рост.
Синтез целлюлозы изучен слабо, однако сейчас совершенно ясно, что, во-первых, целлюлозные микрофибриллы синтезируются на поверхности клетки с помощью ферментного комплекса, связанного с плазматической мембраной, и, во-вторых, ориентация микрофибрилл контролируется микротрубочками, расположенными у внутренней поверхности плазматической мембраны.
Плазмодесмы
Протопласты соседних клеток связаны между собой тонкими нитями цитоплазмы, называемыми плазмодесмами. Хотя такие структуры уже давно наблюдали под световым микроскопом (рис. 2-34), более подробно их изучили только с помощью электронного микроскопа (рис. 2-35).
Рис. 2-34. Световая микрофотография плазмодесм в толстых первичных оболочках клеток эндосперма (питательной ткани семени) хурмы (Diospyros). Плазмодесмы выглядят как тонкие нити, тянущиеся от клетки к клетке и проходящие через оболочки
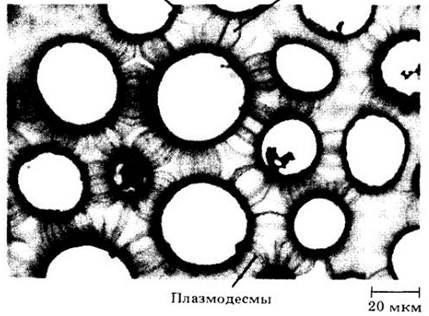
Рис. 2-35. Электронная микрофотография плазмодесм. А. Продольный срез плазмодесм, соединяющих протопласты двух клеток листа кукурузы (Zea mays). Б. Поперечный срез плазмодесм на первичном поровом поле клетки листа кукурузы
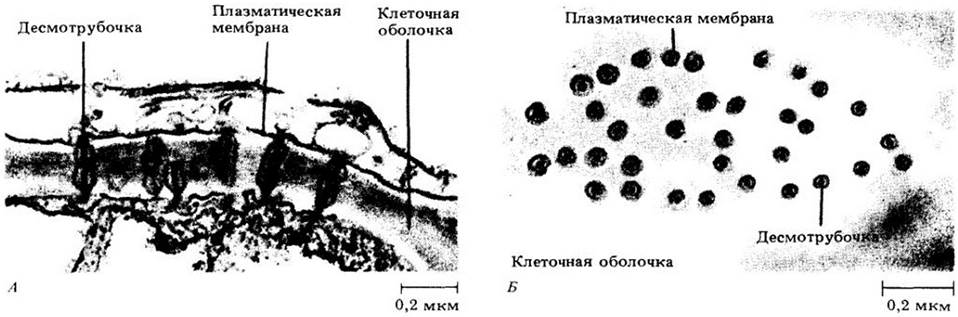
Плазмодесмы либо проходят сквозь клеточную оболочку в любом месте, либо сосредоточены на первичных поровых полях или в мембранах между парами пор. Под электронным микроскопом плазмодесмы выглядят как узкие каналы (диаметром примерно от 30 до 60 нм), выстланные плазматической мембраной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера — десмотрубочка, которая сообщается с эндоплазматическим ретикулумом обеих смежных клеток. Многие плазмодесмы формируются во время клеточного деления, когда трубчатый эндоплазматический ретикулум захватывается развивающейся клеточной пластинкой (см. рис. 2-44). Плазмодесмы могут образоваться и в оболочках неделящихся клеток. Считается, что эти структуры обеспечивают эффективный перенос некоторых веществ от клетки к клетке (см. с.70).
Деление клеток
Одноклеточные организмы растут, поглощая вещества из окружающей среды и используя их для синтеза новых структурных и функциональных молекул. Когда клетка достигает определенного размера, она делится. Две дочерние клетки, каждая из которых примерно вполовину меньше исходной материнской, снова начинают расти. Одноклеточные организмы могут делиться каждый день или даже каждые несколько часов, образуя непрерывный ряд идентичных организмов. У многоклеточных организмов деление клеток наряду с увеличением их размеров является способом роста организма. Во всех случаях новые клетки, образовавшиеся путем деления, сходны по структуре и функциям как с родительской клеткой, так и между собой. Процесс деления у эукариот можно подразделить на две частично перекрывающиеся стадии: митоз и цитокинез. Митоз — это образование из одного ядра двух дочерних ядер, морфологически и генетически эквивалентных друг другу. Цитокинез включает деление цитоплазматической части клетки с образованием дочерних клеток.
Клеточный цикл
Живая клетка проходит ряд последовательных событий, составляющих клеточный цикл (рис. 2-36). Продолжительность цикла варьирует в зависимости от типа клетки и внешних факторов, например, температуры или обеспеченности питательными веществами. Обычно цикл делится на интерфазу и четыре фазы митоза.
Интерфаза
Интерфазу, период между двумя последовательными митотическими делениями, когда-то рассматривали как «фазу покоя». Однако это оказалось не так.
Интерфазу можно разделить на три периода, обозначаемые G1, S и G2 (рис. 2-36). Период G1 (от англ. gap — промежуток) начинается после митоза; в это время увеличивается количество цитоплазмы, включая различные органеллы. Кроме того, согласно современной гипотезе, в период G1 синтезируются вещества, которые либо стимулируют, либо ингибируют период S и остальную часть цикла, определяя, таким образом, произойдет ли вообще деление клетки.
Рис. 2-36. Клеточный цикл можно разделить на митоз и интерфазу — период между двумя последовательными митотическими делениями. Обычно клетки проводят большую часть своей жизни в интерфазе, которую можно подразделить на три периода. Первый из них (G1) — период общего роста и репликации цитоплазматических органелл. В течение второго периода (S) происходит синтез ДНК. В третьем периоде (G2) формируются структуры, необходимые для митотического деления, например, нити веретена. После периода G2начинается митоз. Митоз делится на четыре фазы
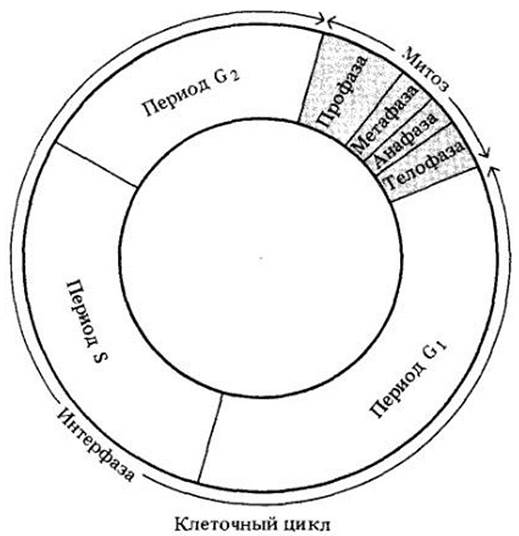
Период S (от англ. synthesis — синтез) следует за периодом G1, в это время происходит удвоение генетического материала (ДНК). В период G2, который следует за периодом S, формируются структуры, непосредственно участвующие в митозе, например, компоненты нитей веретена.
Некоторые клетки проходят неограниченный ряд клеточных циклов. К ним относятся одноклеточные организмы и некоторые клетки зон активного роста (меристем). Ряд специализированных клеток после созревания теряет способность к размножению. Третья группа клеток, например, образующих раневую ткань (каллус), сохраняет способность делиться только в специальных условиях.
Митоз
Митоз, или деление ядра, — это непрерывный процесс, обычно подразделяемый на четыре фазы: профазу, метафазу, анафазу и телофазу (рис. 2-37 — 2-39), В результате митоза генетический материал, удвоившийся в интерфазе, делится поровну между двумя дочерними ядрами.
Рис. 2-37. Схема митоза. В качестве примера взята клетка с четырьмя хромосомами. В ранней профазе хромосомы выглядят как длинные тонкие нити, разбросанные внутри ядра. Постепенно хромосомы укорачиваются и утолщаются и наконец можно увидеть, что каждая состоит из двух нитей (хроматид). Затем исчезают ядрышко и ядерная оболочка. Появление веретена означает начало метафазы, на протяжении которой хромосомы мигрируют в экваториальную плоскость веретена. В поздней метафазе (показанной здесь) центромеры хромосом лежат в этой плоскости. Анафаза начинается с деления центромер; в результате каждая из сестринских хроматид (называемых теперь дочерними хромосомами) имеет свою центромеру. Как показано на рисунке, дочерние хромосомы расходятся к противоположным полюсам веретена. Телофаза в какой-то мере противоположна профазе; она начинается после того, как дочерние хромосомы разошлись к полюсам

Рис. 2-38. Митоз, наблюдаемый с помощью фазово-контрастной оптики в живой клетке хемантуса (Haemanthus katherinae). Клетки раздавлены, чтобы можно было более детально рассмотреть все хромосомы, при этом веретено еле различимо. А. Поздняя профаза: хромосомы конденсированы. Вокруг ядра обозначилась светлая зона. Б.
Поздняя профаза — ранняя метафаза: ядерная оболочка исчезла и концы хромосом видны в цитоплазме. В. Метафаза: хромосомы расположились так, что центромеры лежат в экваториальной плоскости. Г. Средняя анафаза: сестринские хроматиды (называемые теперь дочерними хромосомами) отделились друг от друга и движутся к противоположным полюсам веретена. Д. Поздняя анафаза. Е. Телофаза (цитокинез): дочерние хромосомы достигли противоположных полюсов, и начинается формирование двух дочерних ядер. Образование клеточной пластинки практически завершено
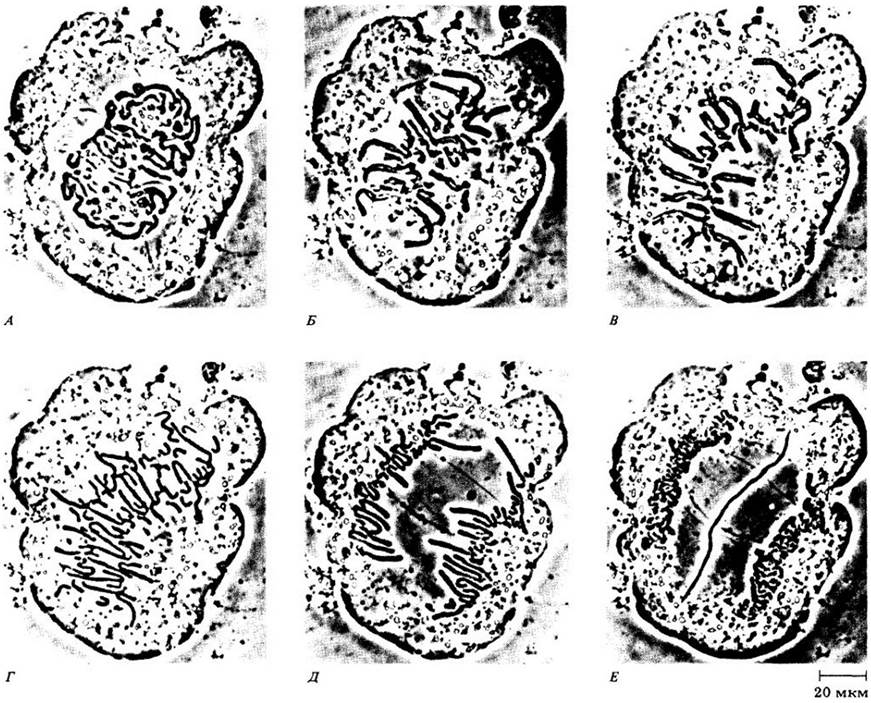
Рис. 2-39. Микрофотография делящихся клеток в кончике корня лука (Allium). Сравнивая рис. 2-37 и 2-38 с этими метками, вы должны определить показанные здесь фазы митоза

Препрофазный поясок
Одним из самых ранних признаков перехода клетки к делению служит появление узкого, кольцеобразного пояска из микротрубочек непосредственно под плазматической мембраной (см. Приложение 1 к этой главе). Этот относительно плотный поясок окружает ядро в экваториальной плоскости будущего митотического веретена (рис. 2-45). Так как он появляется перед профазой, его называют препрофазным пояском. Препрофазный поясок исчезает после формирования митотического веретена, задолго до появления в поздней телофазе клеточной пластинки, которая растет от центра к периферии и сливается с оболочкой материнской клетки в области, ранее занятой препрофазным пояском.
Профаза
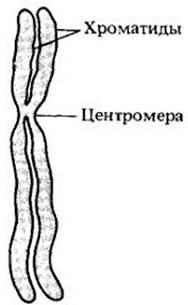
В начале профазы хромосомы напоминают длинные нити, разбросанные внутри ядра. Затем, по мере того как нити укорачиваются и утолщаются, можно увидеть, что каждая хромосома состоит не из одной, а из двух переплетенных нитей, называемых хроматидами. В поздней профазе две укороченные спаренные хроматиды каждой хросомосы лежат рядом почти параллельно и соединены узким участком, который называется центромерой (рис. 2-40). Центромера, имеющая определенное местоположение на каждой хромосоме, делит хромосому на два плеча различной длины.
Рис. 2-40. Схема профазной хромосомы. Хромосомная ДНК реплицируется в интерфазе, и теперь каждая хромосома состоит из двух идентичных хроматид, которые лежат параллельно друг другу. Хроматиды соединяются центромерой
В профазе вокруг ядерной оболочки видна светлая зона, в которой появляются микротрубочки. Вначале микротрубочки имеют случайную ориентацию, а в поздней профазе располагаются параллельно поверхности ядра вдоль оси веретена. Это самое раннее проявление сборки митотического веретена.
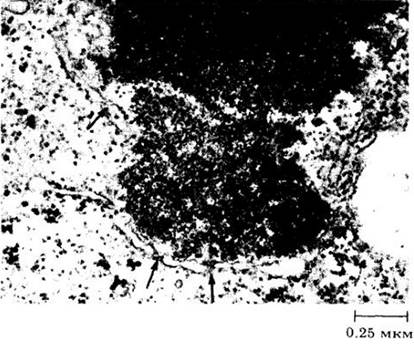
К концу профазы ядрышко постепенно теряет четкие очертания и наконец исчезает. Вскоре после этого распадается и ядерная оболочка (рис. 2-41).
Рис. 2-41. Часть ядра паренхимной клетки стебля хвоща (Еquisetum hyemale) в поздней профазе. Стрелками отмечены поры на фрагментах разрушающейся ядерной оболочки
Метафаза
В начале метафазы веретено (трехмерная структура, наиболее широкая в середине и суживающаяся к полюсам) занимает место, прежде занятое ядром. Нити веретена представляют собой пучки микротрубочек (рис. 2-42). Во время метафазы хромосомы, каждая из которых состоит из двух хроматид, располагаются таким образом, что их центромеры лежат в экваториальной плоскости веретена. Каждая хромосома, по-видимому, прикреплена к нитям веретена своей центромерой (рис. 2-42). Некоторые нити веретена проходят от одного полюса к другому, не прикрепляясь к хромосомам. Располагаются ли хромосомы в экваториальной плоскости случайно, или положение их центромер зафиксировано — до сих пор неизвестно. Этот вопрос представляет значительный интерес для современных исследователей.
Рис. 2-42. Часть метафазной хромосомы в паренхимной клетке стебля хвоща (equisetum hyemale). Некоторые микротрубочки (компоненты нитей веретена) направлены прямо к плотному хромосомному материалу в центромере. Другие микротрубочки (не показаны) идут от полюса к полюсу

Когда все хромосомы расположатся в экваториальной плоскости, метафаза завершается. Хромосомы готовы к делению.
Анафаза
В анафазе (рис. 2-38, Г, Д) хроматиды каждой хромосомы расходятся. Теперь они называются дочерними хромосомами. Прежде всего делится центромера, и две дочерние хромосомы увлекаются к противоположным полюсам. Центромеры, которые еще прикреплены к нитям веретена, движутся впереди, а плечи хромосом тянутся сзади. Когда две дочерние хромосомы отрываются друг от друга, концы длинных плеч разделяются последними.
Нити веретена, прикрепленные к хромосомам, укорачиваются, тем самым способствуя расхождению хроматид и движению дочерних хромосом в противоположные стороны. Высказано предположение, что микротрубочки постоянно формируются на одном конце нити веретена и разбираются на другом. При этом нити веретена тянут дочерние хромосомы к полюсам за центромеры.
В конце анафазы две идентичные группы дочерних хромосом разделены и движутся к противоположным полюсам.
Телофаза
В телофазе (рис. 2-38, Е) завершается обособление двух идентичных групп хромосом, при этом вокруг каждой из них формируется ядерная оболочка (рис. 2-43). Мембраны этих оболочек образуются из шероховатого эндоплазматического ретикулума. Аппарат веретена исчезает. В ходе телофазы хромосомы постепенно теряют четкость очертаний, вытягиваются и снова превращаются в тонкие нити. Ядрышки в это время восстанавливаются. Когда хромосомы становятся невидимыми, митоз завершается, и два дочерние ядра вступают в интерфазу.
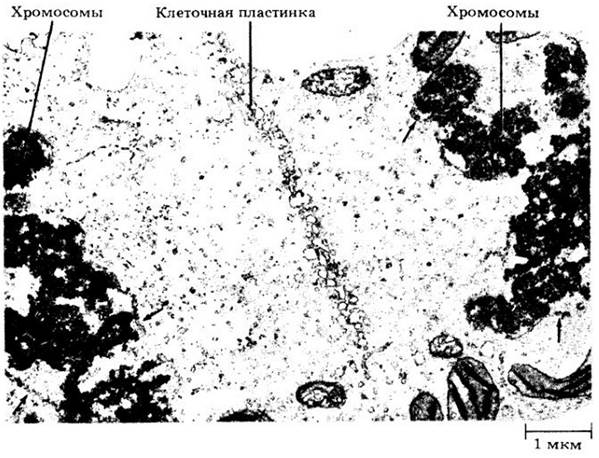
Рис. 2-43. В клетках растений после расхождения дочерних хромосом образуется клеточная пластинка. На этом завершается деление клетки. Здесь можно видеть многочисленные пузырьки Гольджи, сливающиеся на ранней стадии формирования клеточной пластинки. Две группы хромосом по обеим сторонам формирующейся неточной пластинки находятся в телофазе. Стрелками отмечены участки ядерной оболочки, образующейся вокруг разделившихся хромосом
Два дочерних ядра, образовавшихся в процессе митоза, генетически эквивалентны друг другу и материнскому ядру. Это важно, поскольку ядро — контролирующий центр клетки и несет генетическую информацию о синтезе белков (ферментов и структурных компонентов клетки). Эта наследственная программа должна быть в точности передана дочерним клеткам, что в эукариотических организмах обеспечивается путем удвоения хромосом и их последующего деления в процессе митоза.
Продолжительность митоза варьирует у различных организмов, кроме того, она зависит от типа ткани. Однако профаза всегда остается самой длинной фазой, а анафаза — самой короткой. В кончике корня продолжительность профазы может составлять от 1 до 2 ч; метафазы — от 5 до 15 мин; анафазы — от 2 до 10 мин; телофазы — от 10 до 30 мин. И напротив, интерфазы — от 12 до 30 ч.
Организация митотического веретена
Как упоминалось ранее, сборка микротрубочек происходит в особых местах, называемых центрами организации микротрубочек. Во многих эукариотических клетках центры организации микротрубочек, ответственные за формирование митотического веретена, связаны с особыми структурами, называемыми центриолями. До недавнего времени считалось, что именно центриоли служат центрами организации микротрубочек. Сейчас, однако, стало ясно, что эти центры представляют собой плохо очерченные аморфные структуры на полюсах веретена.
Цитокинез
Как, указывалось выше, цитокинез — это процесс деления цитоплазмы. У большинства организмов клетки делятся путем втягивания клеточной оболочки, если она имеется, и образования борозды деления, которая постепенно углубляется и «сжимает» оставшиеся нити митотического веретена. У всех растений (мохообразных и сосудистых растений) и у некоторых водорослей клетки делятся благодаря образованию клеточной пластинки (рис. 2-43 — 2-45).
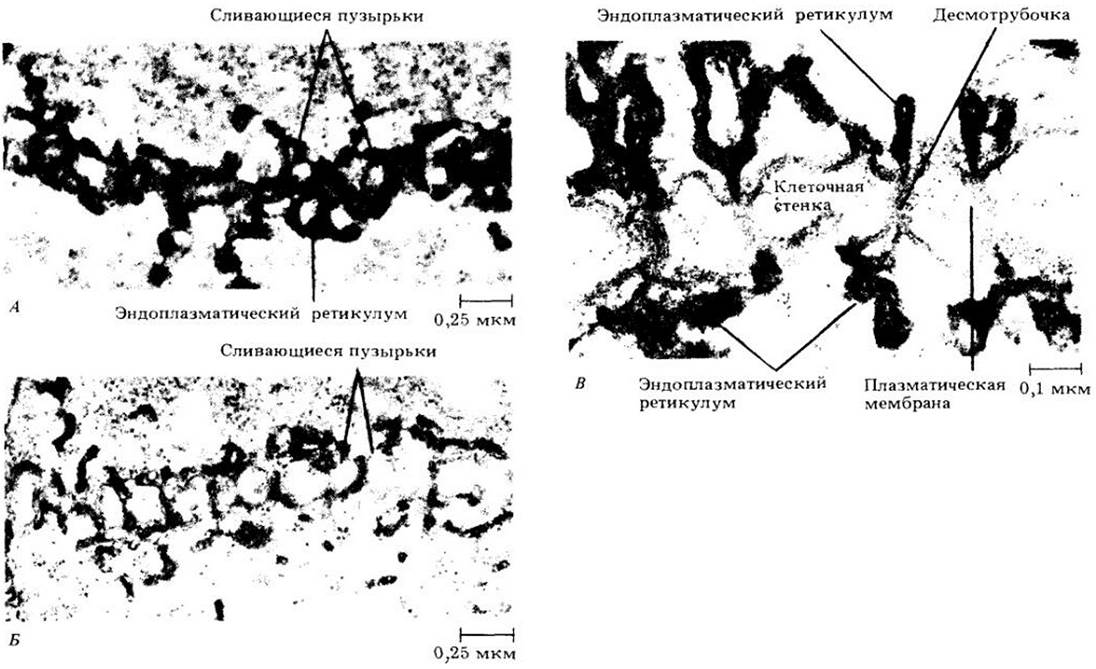
В ранней телофазе между двумя дочерними ядрами формируется бочкообразная система волокон, называемая фрагмопластом. Волокна фрагмопласта, так же, как и волокна митотического веретена, состоят из микротрубочек. В световом микроскопе видно, как в экваториальной плоскости фрагопласта появляются мелкие капли, которые затем сливаются. образуя клеточную пластинку. Клеточная пластинка растет до тех пор, пока не достигнет оболочки делящейся клетки. На этом и завершается разделение двух дочерних клеток. С помощью электронного микроскопа можно видеть, что сливающиеся капельки — это пузырьки, отрывающиеся от аппарата Гольджи. Они в основном содержат пектиновые вещества, из которых формируется срединная пластинка, а мембраны пузырьков участвуют в построении плазматической мембраны по обеим сторонам пластинки. В это время из фрагментов трубчатого эндоплазматического ретикулума образуются плазмодесмы (рис. 2-44).
Рис. 2-44. Последовательные стадии образования клеточной пластинки в метках корня латука (Lactuca sativa), иллюстрирующие связь эндоплазматического ретикулума с развивающейся меточной пластинкой и его участие в образовании плазмодесм. А. Относительно ранняя стадия, на которой сливаются многочисленные мелкие пузырьки, оторвавшиеся от диктиосом. Видны элементы трубчатого эндоплазматического ретикулума. Б. Более поздняя стадия, подтверждающая постоянную тесную взаимосвязь эндоплазматического ретикулума и сливающихся пузырьков. Тяжи трубчатого эндоплазматического ретикулума оказываются включенными в уплотняющуюся клеточную пластинку. В. Зрелые плазмодесмы, состоящие из канала, выстланного плазматической мембраной, и трубочки эндоплазматического ретикулума десмотрубочки
После образования срединной пластинки каждый протопласт откладывает над ней первичную оболочку. Кроме того, каждая дочерняя клетка откладывает новый слой оболочки вокруг всего протопласта, которая продолжает оболочку, возникшую из клеточной пластинки (рис. 2-45). Исходная оболочка родительской клетки разрушается по мере роста дочерних клеток.
Рис. 2-45. Перераспределение микротрубочек в течение клеточного цикла и по мере образования клеточной стенки. А. В интерфазе, во время роста и дифференцировки клеток, микротрубочки лежат непосредственно под плазматической мембраной. Б. Перед профазой кольцевой пояс микротрубочек, или препрофазный поясок, окружает ядро в экваториальной плоскости будущего митотического веретена. В. В метафазе микротрубочки образуют веретено. Г. Когда микротрубочки веретена исчезают, новые микротрубочки образуют между двумя дочерними ядрами фрагмопласт. Клеточная пластинка формируется в экваторе фрагмопласта между двумя ядрами и растет от центра, пока не достигнет оболочки делящейся клетки. Д. Каждая из дочерних клеток формирует собственную первичную оболочку. Е. По мере роста дочерних клеток (здесь показана только верхняя) оболочка материнской клетки разрушается. Д и Е. Микротрубочки опять лежат под плазматической мембраной
