СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 3. МОЛЕКУЛЯРНЫЙ СОСТАВ КЛЕТОК
Как указывалось, в предыдущей главе, клетки, несмотря на разнообразие, имеют сходную структуру. И сходство это становится еще более поразительным, когда мы начинаем исследовать клетки на молекулярном уровне.
Земная кора и атмосфера содержат 92 природных химических элемента. Из всех этих элементов лишь ограниченное количество было отобрано в ходе эволюции для формирования сложного, высокоорганизованного живого вещества. Фактически, как показано в табл. 3-1, около 99% (по массе) протоплазмы построено всего из шести элементов, которые входят в состав органических соединений или растворены в воде.
Рис. 3-1. Крахмальные зерна в амилопластах клубня картофеля, сфотографированные в поляризованном свете. Крахмал, состоящий из углерода, водорода и кислорода, является главным запасным полисахаридом растений

Таблица 3-1. Атомный состав различных организмов (в процентах сухой массы)
Элемент С |
Человек 19,37 |
Люцерна 11,34 |
Бактерии 12,14 |
Н |
9,31 |
8,72 |
9,94 |
N |
5,14 |
0,83 |
3,04 |
О |
62,81 |
77,90 |
73,68 |
Р |
0,63 |
0,71 |
0,60 |
S |
0,64 |
0,10 |
0,32 |
CHNOPS вместе: |
97,90 |
99,60 |
99,72 |
Вода составляет более половины массы живых тканей, в частности более 90% массы большинства тканей растений. Напротив, на долю ионов, например, калия (К+), натрия (Na+) и кальция (Са2+), приходится не более одного процента. Оставшаяся часть клетки почти полностью состоит из органических молекул.
Органические соединения
По определению органические соединения — это соединения, содержащие углерод. Помимо углерода почти все органические соединения содержат водород и большинство из них — кислород (рис. 3-1). Азот и сера встречаются реже, чем кислород, и только малая доля органических соединений содержит другие элементы. Поскольку протоплазма построена из немногих элементов, основную сухую массу живых клеток составляют четыре типа органических соединений. К ним относятся углеводы, липиды, белки и нуклеиновые кислоты (табл. 3-2).
Таблица 3-2. Некоторые основные классы органических соединений
Органические соединения |
Функции |
Компоненты |
Химический состав |
Углеводы |
Источник энергии, структурный материал, строительные блоки для других молекул |
Простые сахара |
Углерод, водород и кислород |
Липиды |
Запасание энергии, структурный материал, препятствие для потери влаги |
Жирные кислоты, глицерол |
Углерод, водород и кислород |
Белки |
Структурный материал, ферменты |
Аминокислоты |
Углерод. водород, кислород, азот и сера |
Нуклеиновые кислоты |
Синтез белка |
Нуклеотиды (азотистые основания, сахара и фосфаты) |
Углерод, водород, кислород, азот и фосфор |
Углеводы
Углеводы (сахара, крахмал и др.) — это соединения, содержащие углерод в сочетании с водородом и кислородом. Углеводы — самые распространенные в природе органические вещества. Крупные молекулы, построенные из сходных или идентичных субъединиц (мономеров), называются полимерами. В случае углеводов мономерами являются моносахариды, а полимерами — полисахариды.
Моносахариды и дисахариды
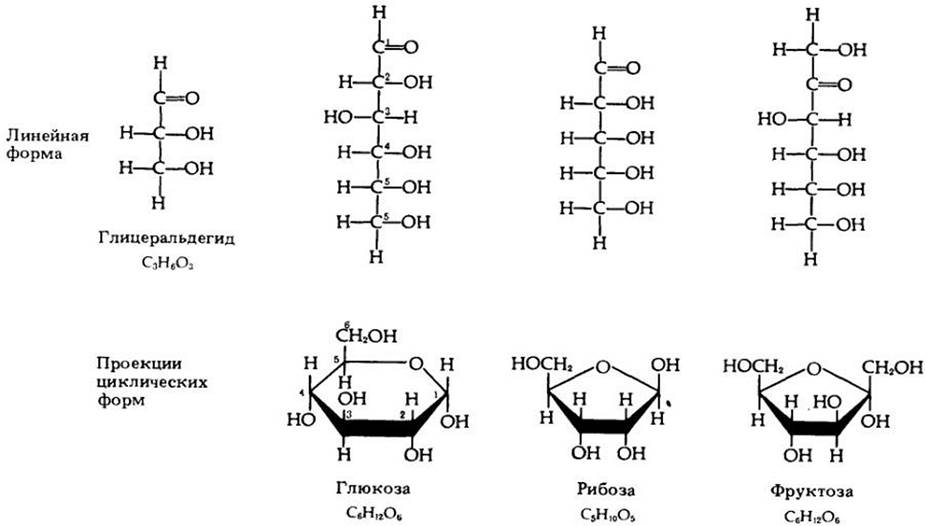
Моносахариды — самые простые углеводы. Они состоят из цепочки атомов углерода, к которым присоединены атомы водорода и кислорода в соотношении 1:2:1 (СН2О). Примеры нескольких моносахаридов приведены на рис. 3-2. Как показано на рисунке, молекулы пятиуглеродных и шестиуглеродных сахаров могут существовать и в кольцевой форме (например, при растворении в воде).
Рис. 3-2. Примеры биологически важных моносахаридов. Пятиуглеродные сахара (пентозы) и шестиуглеродные сахара (гексозы) существуют в линейной и циклической формах. Атомы углерода нумеруются. Нумерация углеродных атомов показана для двух форм молекулы глюкозы. При этом в циклических формах атомы углерода не обозначаются; их присутствие подразумевается в каждом углу кольца
Дисахариды построены из двух моносахаридов. Рис. 3-3 иллюстрирует соединение двух молекул моносахаридов в молекулу дисахарида. Реакция сопровождается выделением молекулы воды. Дисахарид в свою очередь может быть подвергнут гидролизу — расщеплению на исходные моносахариды при добавлении молекулы воды (одной на каждую связь). Гидролиз представляет собой экзергоническую реакцию — энергия химических связей ее продуктов меньше, чем исходных молекул. Поэтому в такой реакции энергия выделяется. Напротив, соединение двух моносахаридов с образованием дисахарида требует притока энергии.
Рис. 3-3. Моносахариды глюкоза и фруктоза могут соединяться с образованием дисахарида сахарозы, основного транспортируемого растений. Связи между моносахаридами формируются путем удаления молекулы воды. Образование сахарозы требует поглощения энергии из расчета 5,5 ккал/моль
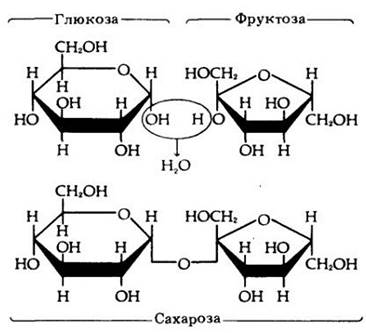
В организме животных транспортируемые сахара чаще всего представлены глюкозой. Глюкоза и фруктоза образуют сахарозу — дисахарид, характерный для растений. Сахароза — это обыкновенный сахар (тростниковый или свекловичный), который мы едим. Лактоза (молочный сахар) — дисахарид, построенный из глюкозы и галактозы.
Первичным источником сахара для всех растительных клеток служит фотосинтез. В процессе фотосинтеза солнечная энергия преобразуется в энергию химических связей, необходимую для формирования молекулы глюкозы. Когда молекула сахара расщепляется, энергия химических связей высвобождается.
Полисахариды
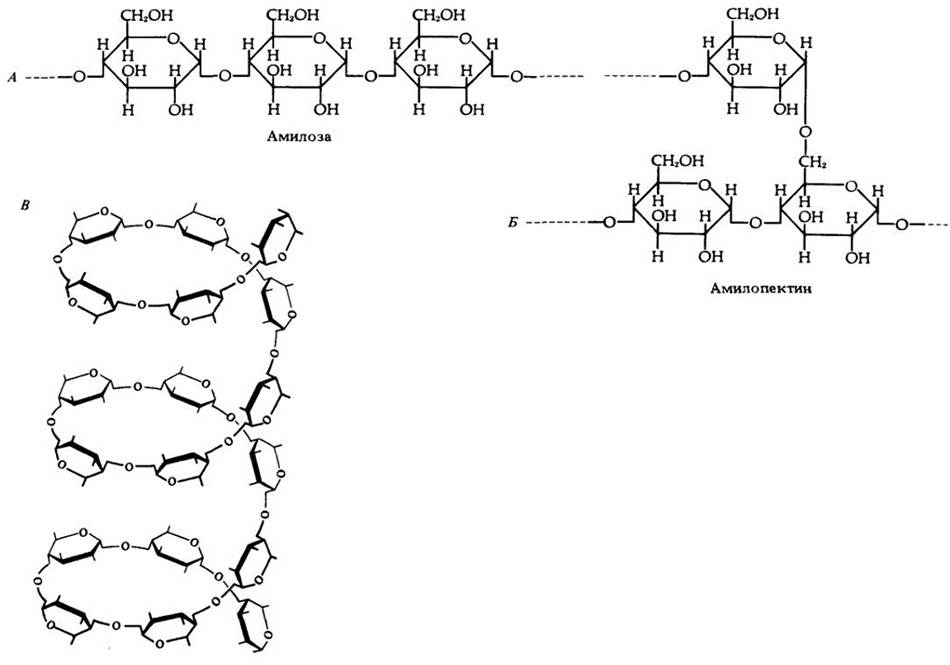
Полисахариды состоят из моносахаридов, соединенных друг с другом в длинные цепи. Некоторые полисахариды являются запасными веществами. Крахмал, построенный из молекул глюкозы (точнее, ее остатков) — основной запасный полисахарид растений (рис. 3-1 и 3-4), а гликоген — грибов, бактерий и животных. Запасные углеводы гидролизуются с высвобождением энергии.
Рис. 3-4. Сахара запасаются в растениях в форме крахмала. Крахмал построен из двух полисахаридов: амилозы (А) и амилопектина (Б). Одна молекула амилозы содержит 1000 и более остатков глюкозы, образующих длинную неразветвленную цепь, которая закручивается в форме спирали (В). Молекула амилопектина имеет молекулярную массу от 1 до 6 млн. и ветвится примерно через каждые 20 — 25 остатков глюкозы, представляющих собой мономеры альфа-глюкозы (см. рис. 3-5).
Возможно, спиральная структура молекул крахмала способствует откладыванию этого полимера в виде зерен (см. рис. 2-10 и 3-1). Обычный запасной полисахарид животных, грибов и бактерий — это гликоген, который похож на амилопектин по общему строению и отличается тем, что ветвление происходит через каждые 6 — 12 остатков глюкозы. Молекулярная масса гликогена достигает 100 млн
Полисахариды — это и важнейшие структурные компоненты клеток. У растений основной структурный полисахарид — целлюлоза (рис. 3-5). Более того, целлюлоза — самый распространенный в природе полисахарид. Хотя она, как и крахмал, построена из остатков глюкозы, ее нитевидные молекулы соединены в длинные жесткие цени, что определяет и иную биологическую роль. К тому же из-за различий в способах соединения глюкозных остатков у крахмала и целлюлозы последняя нелегко гидролизуется. Помимо целлюлозы оболочки растительных клеток могут содержать ряд других полисахаридов (рис. 3-6), пектиновые вещества (пектины) (рис. 3-7) и лигнин.
Рис. 3-5. В живых системах циклические формы альфа- и бета-глюкозы находятся в равновесии. Переход из одной циклической формы в другую осуществляется через линейную форму (А). В то время как крахмал состоит из мономеров альфа-глюкозы, целлюлоза (Б) построена из мономеров бета-глюкозы. Группы -ОН (окрашены), отходящие от обеих сторон цепи, образуют водородные связи с соседними группами -ОН, и в результате этого формируются тяжи из параллельно соединенных цепей (В). В молекуле крахмала большинство групп -ОН способно к образованию водородных связей снаружи спирали
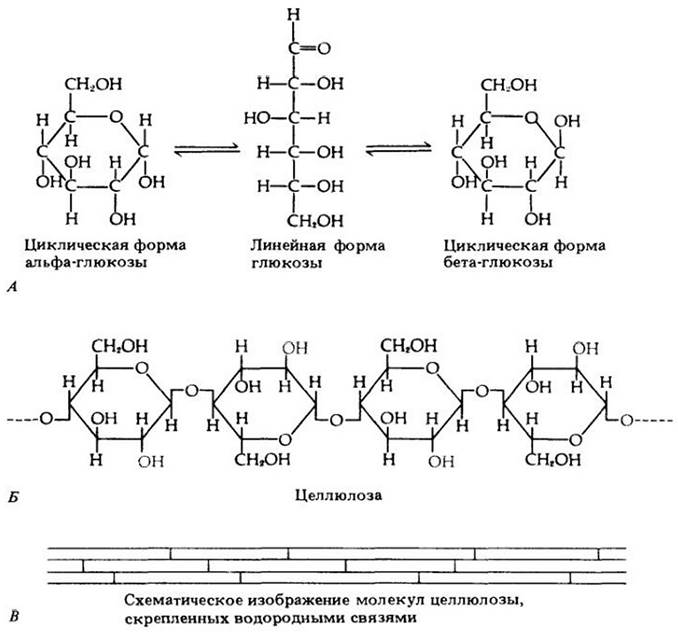
Рис. 3-6. Строительные блоки некоторых полисахаридов, наряду с целлюлозой входящих в состав клеточных оболочек растений. Показанные здесь моносахариды являются мономерами гилактана, маннана, ксилана и арабинана
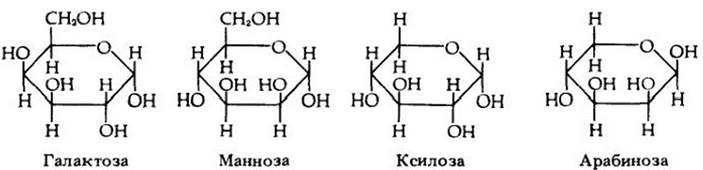
Рис. 3-7. Пектиновые соединения построены из остатков альфа-галактуроновой кислоты (А), которая является производным глюкозы. Пектиновая кислота показана внизу (Б). Кальциевые и магниевые соли пектиновой кислоты образуют основу срединной пластинки, скрепляющей соседние растительные клетки. В пектине и протопектине разное число водородных ионов (выделены цветом) замещается метильными группами (-CH¿). Протопектин — обычный компонент клеточной стенки. Он менее растворим, чем пектин, который содержится в растительных соках
Когда целлюлоза включается в клеточную оболочку, она не может служить растению в качестве источника энергии. Только некоторые бактерии, грибы и простейшие, а также очень немногие животные (тарпон1), например) имеют ферментные системы, способные расщеплять целлюлозу. Жвачные животные, термиты и тараканы могут использовать целлюлозу как источник энергии только благодаря микроорганизмам (которые обладают необходимыми ферментными системами), населяющим их пищеварительные тракты.
1Тарпон — представитель тарпонообразных, одного из отрядов костистых рыб с многими архаичными особенностями морфологии. Атлантический тарпон (Megalops atlanticus) — крупная (до 180 см) и очень сильная рыба, объект спортивного лова у юго-восточных берегов Северной Америки. — Прим. перев.
Хитин — другой важнейший структурный полисахарид. Он входит в состав клеточных оболочек грибов и формирует твердый наружный покров (экзоскелет) насекомых и ракообразных. Мономер хитина — шестиуглеродный сахар, к которому присоединена азотсодержащая группа (см. рис. 13-5).
Липиды
Липиды — это жиры и жироподобные вещества, имеющие две отличительные особенности: во-первых, они. как правило, гидрофобны («боятся воды») и поэтому нерастворимы в воде (хотя растворяются в других липидах); во-вторых, содержат много углерод-водородных связей, и при их окислении высвобождается больше энергии, чем при окислении других органических соединений. Жиры дают в среднем около 9,3 ккал/г, тогда как углеводы — около 3,8 ккал/г. Указанные особенности липидов определяют их роль как структурных компонентов и запасных источников энергии.
Жиры
Жиры — важнейшие запасные вещества (рис. 3-8). Некоторые растения накапливают жиры (масла) в больших количествах, особенно в семенах и плодах. Клетки синтезируют жиры из сахаров. Молекула жира состоит из трех жирных кислот, соединенных с молекулой глицерола (рис. 3-9). Жирные кислоты представляют собой длинные углеводородные цени, несущие на концах карбоксильную группу, которая придает им свойства слабой кислоты. Глицерол образует связь с карбоксильной группой, высвобождая молекулу воды, и, таким образом, служит как бы «клеем» или «скрепкой» для жирных кислот. Подобно полисахаридам и белкам, жиры расщепляются при гидролизе.
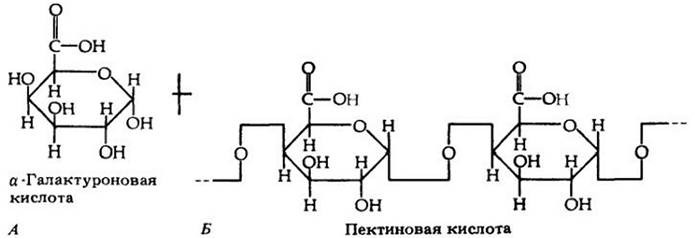
Рис. 3-8. Две камбиальные клетки мясистого подземного стебля (клубнелуковицы) полушника (Isoeles muricatа). Зимой эти клетки запасают большое количество жира (видны липидные капли) и углеводов (видны крахмальные зерна в амилопластах). В каждой клетке находится несколько вакуолей. Темный материал. отложившийся внутри вакуолей, — таннин
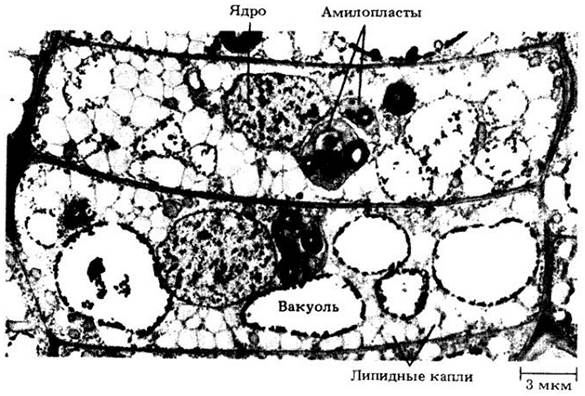
Рис. 3-9. Молекула жира состоит из трех жирных кислот, соединенных молекулой глицерола. Связи образуются путем отщепления молекулы воды (отщепляемые атомы выделены цветом). Жирная кислота, например, стеариповая, у которой каждый атом углерода соединен с предельно возможным числом атомов водорода, называется насыщенной. Жирная кислота, например, олеиновая, у которой некоторые атомы углерода соединены только с одним атомом водорода, является ненасыщенной. Ненасыщенные жирные кислоты чаще встречаются у растений, чем у животных
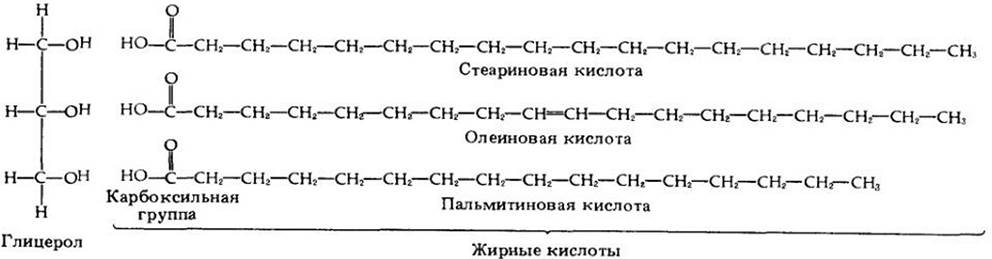
Физические свойства жира определяются длиной цепей жирных кислот и тем, являются кислоты насыщенными или ненасыщенными. У насыщенных жирных кислот все атомы углерода удерживают столько водородных атомов, сколько вообще возможно. Ненасыщенные кислоты содержат атомы углерода, соединенные двойными связями. Эти атомы углерода способны образовывать дополнительные связи с другими атомами (отсюда термин «ненасыщенные»). Ненасыщенные жиры, как правило, маслянисто-жидкие; они более характерны для растений, чем для животных; примерами служат сафлоровое, арахисовое и кукурузное масла. Животные жиры, такие, как сало и сливочное масло, содержат насыщенные жирные кислоты и имеют обычно более высокую температуру плавления, чем ненасыщенные жиры.
Кутин, суберин и воска
Кутин и суберин — уникальные нерастворимые липидные полимеры, являющиеся важными структурными компонентами клеточных оболочек многих растений. Их главная функция заключается в том, что они образуют основу, в которую погружены воска — длинноцепочечные липидные соединения. Воска совместно с кутином или суберином формируют защитные слои, которые уменьшают потерю влаги и других веществ воздушными или подземными частями растения. Основную барьерную роль при этом играют воска.
Кутин вместе с погруженными в него восками образует кутикулу, которая покрывает наружные стенки эпидермальных клеток. На рис. 3-10 видно, что кутикула имеет сложную структуру и состоит из нескольких слоев. Наружный слой сформирован восками, отложенными на поверхности, и называется эпикутикулярным воском (рис. 3-11). Под ним расположена собственно кутикула, состоящая из кутикулярного воска и кутина. Ниже могут лежать один или более так называемых кутикулярных слоев, образованных целлюлозой, кутином и воском. Наконец, между кутикулой и клеточной оболочкой может лежать еще слой пектина.
Рис. 3-10. Схематическое изображение слоев кутикулы. Первый слой — эпикутикулярный воск — состоит полностью из воска. Второй слой — собственно кутикула — состоит из воска и кутина. Нижележащий кутикулярный слой (или слои) образован целлюлозой и кутином, в нем может присутствовать и воск. Еще ниже находится слой пектина, примыкающий к клеточной оболочке, которая сама построена из целлюлозы и пектина
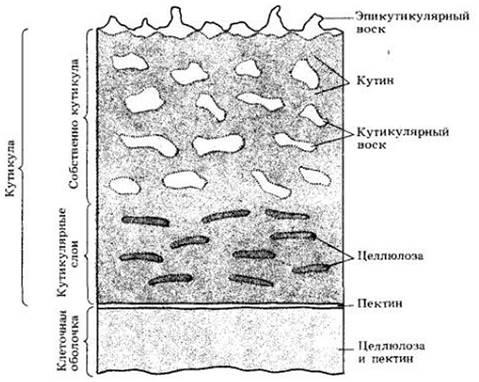
Рис. 3-11. Поверхность листа Eucalyptus cloeziana, на которой видны обложения эпикутикулярного воска. Под ними расположена содержащая воск кутикула, покрывающая наружные стенки эпидермальных клеток. Воска защищают открытые поверхности растения от потери воды

Суберин, основной компонент клеточных оболочек пробки, содержится и в поясках Каспари клеток эндодермы (см. гл. 21), и в клеточных стенках обкладок проводящих пучков в листьях злаков. Под электронным микроскопом содержащие суберин оболочки выглядят ламеллярными (слоистыми), состоящими из чередующихся светлых и темных полос (см. рис. 21-13). Считается, что светлые полосы состоят из восков, темные — из суберина. Некоторые кутикулы также выглядят слоистыми.
Фосфолипиды
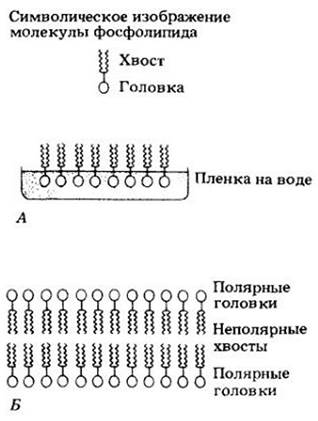
К жирам очень близки фосфолипиды — разнообразные соединения, в которых глицерол связан с двумя жирными кислотами и молекулой, содержащей фосфор (рис. 3-12). Роль фосфолипидов в клетке очень велика, особенно — в построении клеточных мембран.
Рис. 3-12. Молекула фосфолипида состоит из двух жирных кислот, связанных, как в молекуле жира, с молекулой глицерола, и из фосфатной группы (показанной в цвете), присоединенной к третьему атому углерода глицерола. Фосфатная группа содержит еще дополнительную химическую группу, обозначаемую буквой R. Хвосты жирных кислот неполярны (не имеют ни положительных, ни отрицательных зарядов) и поэтому нерастворимы в воде. Полярные головки, содержащие фосфатную и R группы, гидрофильны (растворимы в воде)
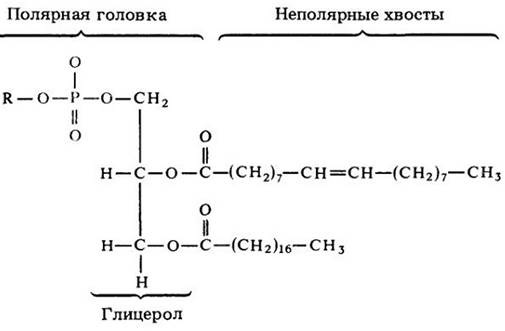
Фосфорный конец молекулы фосфолипида является гидрофильным («любящим воду») и поэтому растворим в воде в отличие от гидрофобных жирных кислот. Добавленные к воде фосфолипиды стремятся образовать пленку на ее поверхности, причем их полярные головки погружаются в воду, а нерастворимые цепи жирных кислот (хвосты) торчат над водой (рис. 3-13, А). В водной среде клеток фосфолипиды самоорганизуются в ряды, в которых нерастворимые жирные кислоты направлены друг к другу, а фосфатные концы — наружу (рис. 3-13, Б). Такие конфигурации занимают важное место в структуре клеточных мембран (см. с. 67).
Рис. 3-13. Глицеролфосфатные участки гидрофильны (растворимы в воде), а жирные кислоты — гидрофобны. Поэтому в воде молекулы фосфолипидов располагаются на ее поверхности таким образом (А), что полярные гидрофильные головки погружены в воду, а неполярные гидрофобные хвосты поднимаются над водой. В клетке эти молекулы самоорганизуются в слои (Б); при этом растворимые головки направлены наружу