СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ III. ГЕНЕТИКА
ГЛАВА 8. ХИМИЯ И НАСЛЕДСТВЕННОСТЬ
Природа ДНК
В 1951 г. американский генетик Джеймс Д. Уотсон приехал в Англию, где договорился о совместной работе с Фрэнсисом Криком в Лаборатории Кавендиша в Кембридже. Уотсон и Крик были из числа ученых, убежденных в том, что ДНК, а не белок представляет собой основное вещество наследственности. По словам Уотсона, ДНК — «самая золотая из всех молекул».
Свои исследования организации молекулы ДНК Уотсон и Крик частично основывали на представлении о генах, хромосомных единицах наследственности, которое уже сложилось к этому времени. Они знали: чтобы ДНК могла быть генетическим материалом, она должна отвечать по крайней мере четырем требованиям.
1. Переносить генетическую информацию от клетки к клетке и от поколения к поколению; более того, она должна нести большое количество информации. (Подумать только, как много должно содержаться инструкций в наборе генов, которые управляют развитием слона, дерева или даже бактерии.)
2. Удваиваться перед каждым клеточным делением подобно тому, как это делают хромосомы; более того, она должна реплицироваться с большой точностью. (Из накопленных данных о частоте мутаций, например, у человека, мы знаем, что любой человеческий ген должен копироваться без малейшей ошибки в среднем в течение миллионов лет.)
3. С другой стороны, ген должен иногда изменяться, или мутировать. (Когда ген меняется, т. е. совершается ошибка, то должна копироваться «ошибка», а не первоначальная последовательность. Это наиболее важное свойство, вероятно, уникальное для всех живых существ, поскольку без способности копировать «ошибки» не может быть эволюции путем естественного отбора.)
4. Должен существовать некий механизм «считывания» накопленной информации и преобразования ее в живом организме.
Уотсон и Крик хорошо понимали, что молекула ДНК может быть генетическим материалом только в том случае, если будет доказано, что ее размер, конфигурация и сложность достаточны, чтобы кодировать и копировать огромное количество информации, в которой нуждаются живые организмы.
В своей работе Уотсон и Крик использовали информацию, полученную ранее на основе биохимических исследований ДНК, в том числе следующие данные:
1. Молекула ДНК очень крупная, кроме того, длинная и тонкая.
2. Три ее компонента (азотистое основание, сахар и фосфат) образуют нуклеотиды, как показано на рис. 3-19.
3. Розалинд Франклин и Морис Уилкинс из Королевского колледжа в Лондоне исследовали дифракцию рентгеновских лучей, проходящих через препараты ДНК, и обнаружили, что ее длинные молекулы состоят из регулярно повторяющихся единиц, которые, по-видимому, расположены по спирали.
4. ак показано в табл. 8-1, отношение нуклеотидов, содержащих аденин и тимин, составляет 1:1, т. е. точно такое же, как между нуклеотидами, содержащими гуанин и цитозин.
Уотсон и Крик не проводили экспериментальных исследований, но собрали воедино все известные сведения о структуре ДНК. В своей работе они использовали все указанные выше экспериментальные данные и попытались построить металлическую модель ДНК, соответствующую физическим и химическим данным о ее структуре (рис. 8-4, А).
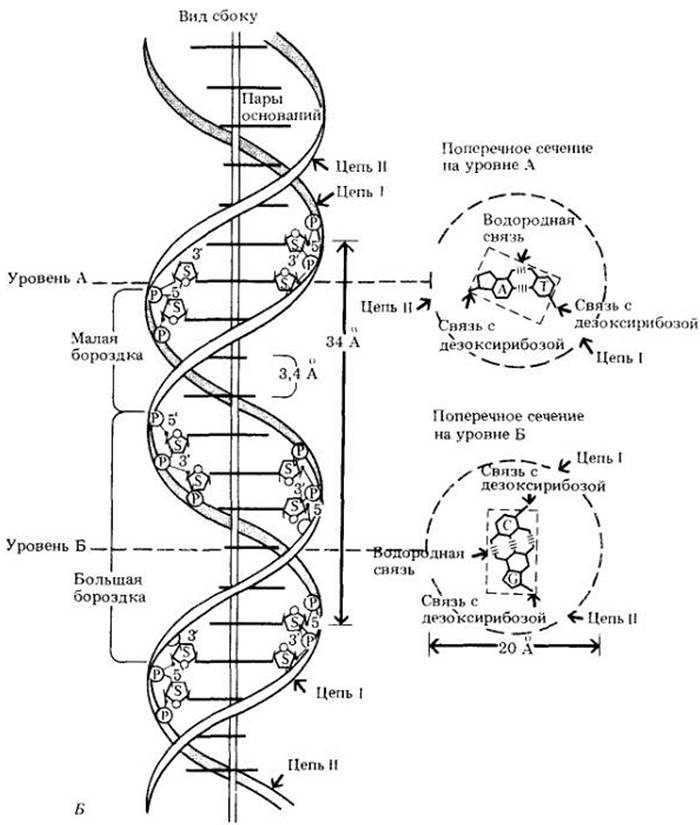
Рис. 8-4. А. Уотсон (слева) и Крик с их металлической моделью ДНК. Б. Схема строения молекулы ДНК, предложенная Уотсоном и Криком. Слева — боковой вид молекулы ДНК (указана вертикальная ось). Молекула состоит из полипуклеотидных цепей, которые закручены в правосторонние спирали и переплетены вместе, образуя двойную спираль. Ее средний диаметр — 2 нм (20 А). Две цепи, образующие спираль, состоят из нуклеотидов, содержащих остатки дезоксирибозы (8). При этом сахар каждого нуклеотида связан через фосфатную группу (Р) с сахаром соседнего. Правильное чередование сахаров и фосфатов образует остов молекулы. Сахара каждой спирали вдаются внутрь цилиндра. Пары азотистых оснований (показанные жирными горизонтальными линиями) занимают центральную область цилиндра (пунктирные прямоугольники на поперечных сечениях справа). Пары оснований образованы тимином (Т) и аденином (А) на уровне А и цитозином (С) и гуанином (G) на уровне Б. Основания соединены водородными связями. Круги, нарисованные штриховыми линиями, указывают внешний край двойной спирали, если смотреть на спираль с ее конца. Основания уложены стопкой с интервалом в 3,4 А и на каждой ступени повернуты на 36°. Таким образом, имеется 10 пар оснований на виток спирали. В результате такого поворота пары оснований, если смотреть сбоку, имеют вид линий различной длины в зависимости от угла зрения

Самым важным, с их точки зрения, был вопрос: каким образом химическая структура ДНК отражает ее биологическую функцию? Вспоминая свои исследования, Уотсон писал: «В моменты пессимистического настроения мы часто опасались, что правильная структура будет скучной, т. е. абсолютно ничего не значащей». На деле же структура ДНК оказалась невероятно «увлекательной».
Двойная спираль
Связывая вместе различные данные, Уотсон и Крик сделали вывод, что ДНК — не одиночная спираль, как у многих белков, но гигантская, тесно переплетенная двойная спираль. Представьте себе перила винтовой лестницы, которые образуют простую спираль. Если лестница заворачивается в виде спирали, а ступеньки сохраняют положение, перпендикулярное ее боковым краям, то получится грубая модель двойной спирали.
В молекуле ДНК «края» состоят из чередующихся молекул дезоксирибозы и фосфата (см. рис. 8-4, Б и 8-5), а «ступени» образованы азотистыми основаниями — аденином (А), тимином (Т), гуанином (G) и цитозином (С) — по одному основанию на каждый сахар-фосфат, причем каждую ступеньку образуют два основания. Основания соединены водородными связями — относительно слабыми химическими связями, которые играют важную роль в образовании вторичной и третичной структур белков (рис. 8-5).
Расстояние между «краями», согласно измерениям Уилкинса, составляет 2 нм. Если бы связывались два пурина, то расстояние между ними оказалось бы большим, а два пиримидина не «достали» бы друг друга. Но все становится на свои места, если пурин соединяется с пиримидином. Поэтому пара оснований — ступенька винтовой лестницы — всегда представлена пурином и пиримидином (см. рис. 8-5). По этой причине отношение пуринов и пиримидинов в молекуле ДНК всегда равно 1:1.
Уотсон и Крик отметили, что последовательность нуклеотидов каждой цепи двойной спирали может быть любой, например, АТGСGТАСАТТ и т. д. Поскольку длина молекулы ДНК может составлять несколько тысяч нуклеотидов, то возможно ожидать и большого разнообразия в порядке чередования нуклеотидов. Число пар оснований колеблется в пределах от 5000 для простейшего вируса до 5 000 000 000 в 46 хромосомах человека. Если ДНК одной клетки человека расположить линейно, то длина нити достигнет 1,5 м; количество информации, заключенное в ней, уложится на 600 000 печатных страниц в среднем по 500 слов в каждой, что равноценно библиотеке в 1000 книг. Короче говоря, молекула ДНК действительно способна хранить необходимую генетическую информацию.
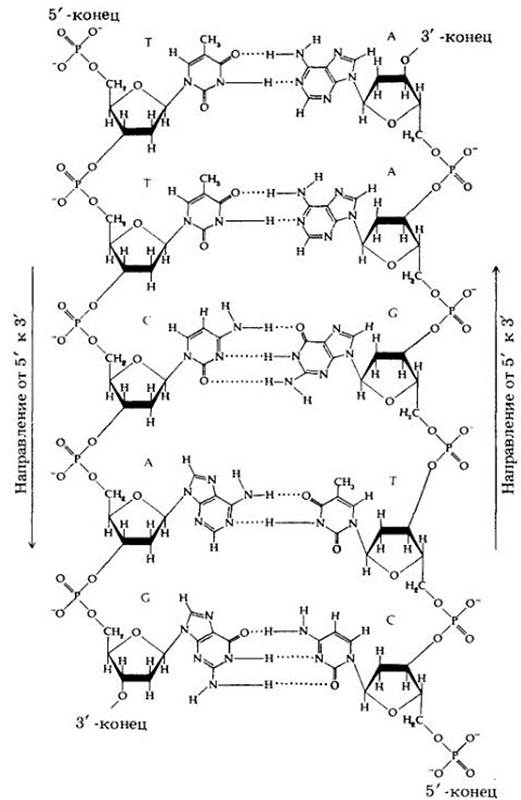
Рис. 8-5. Двухцепочечная структура участка молекулы ДНК. Каждый нуклеотид состоит из сахара (дезокси- рибозы), фосфатной группы и азотистого основания (пурина или пиримидина). Следует обратить внимание на повторяющуюся последовательность сахар-фосфат-сахар-фосфат, которая образует остов молекулы. Каждая фосфатная группа связана с 5'-атомом углерода одного сахара и 3'-атомом углерода сахара соседнего нуклеотида, так что цепь имеет два конца — 5' и 3'. Мостики, образуемые фосфатными группами между нуклеотидами, направлены противоположно, т. е. цепи антипараллельны (сравните этот рис. с рис. 8-4, Б)
Молекула, которая копирует сама себя
Наиболее интересное открытие произошло, когда Уотсон и Крик попытались соединить две цепи ДНК, а именно: они обнаружили, что не только пурины не могут связываться с пуринами, и пиримидины с пиримидинами, но аденин может образовать пару только с тимином, а гуанин — с цитозином. Только эти пары азотистых оснований образуют водородные связи; аденин не может образовать их с цитозином, а гуанин — с тимином.
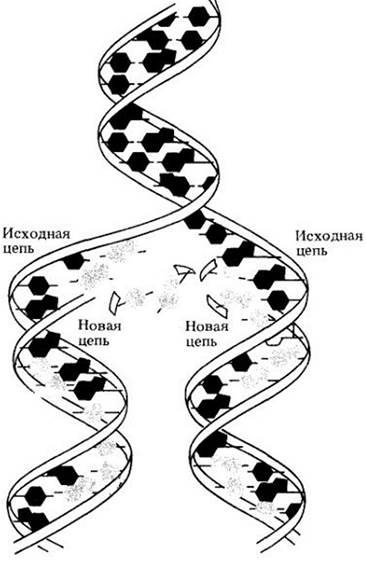
Посмотрим снова на рис. 8-1. Модель Уотсона — Крика просто и логично объясняет состав оснований в ДНК. Может быть, самое важное свойство модели состоит в том, что две цепочки комплементарны, т. е. каждая содержит такую последовательность оснований, которая дополняет другую. Когда молекула ДНК «воспроизводит себя», она просто «расплетается»; водородные связи между азотистыми основаниями рвутся (рис. 8-6). Две цепи расходятся, и вдоль каждой образуются новые. Если в материнской цепи присутствует тимин, то к нему пристраивается только аденин, точно так же, как гуанин образует пару только с цитозином. Таким образом на каждой исходной цепи образуется комплементарная ей цепь, и в результате воспроизводятся две точные копии материнской ДНК. Так открытие структуры ДНК помогло в принципе ответить на старый вопрос: каким образом наследственная информация удваивается и переходит от поколения к поколению.
Рис. 8-6. Изображенная здесь молекула ДНК находится в процессе репликации. Материнские цепи расплетаются по мере того, как рвутся водородные связи между основаниями. (Для большей ясности основания показаны вне плоскости.) Каждая из материнских цепей служит матрицей, на которой из имеющихся в клетке нуклеотидов строится новая комплементарная цепь
По поводу этой, одной из величайших загадок всех времен Уотсон и Крик писали в своей краткой первой публикации: «От нашего внимания не ускользнуло то, что постулированное нами специфическое образование пар сразу предполагает возможный механизм копирования наследственного материала». В 1962 г., через 9 лет после публикации своей оригинальной гипотезы, Уотсон, Крик и Уилкинс были удостоены Нобелевской премии, что свидетельствовало об огромной важности их исследования.
В последствии было показано, что из-за очень большой длины и сложности строения (двойная спираль) процесс репликации ДНК происходит в несколько этапов, катализируемых ферментами. Для расхождения и раскручивания цепей ДНК и правильного расположения комплементарных оснований необходимы специфические ферменты. Механизм работы этих ферментов открывает путь к пониманию процесса репликации ДНК.
По мере того как химическая сторона этого процесса становилась более понятной, появились дополнительные «сложности» — концы двух цепей двойной спирали различны. В каждой цепи фосфатная группа, которая соединяет две молекулы дезоксирибозы, связана с одним сахаром в 5'-положении (пятый углеродный атом кольца дезоксирибозы), а с соседним сахаром — в З'-положении (третий углеродный атом кольца дезоксирибозы). Благодаря подобной структуре связи сахар-фосфат-сахар каждая цепь имеет 5'- и З'-концы (см. рис. 8-5).
Кроме этого, поскольку в одной цепи синтез ДНК направлен от 3'- к 5'-концу, то в другой цепи он имеет противоположное направление. Таким образом, две цепи ДНК антипараллельны (см. рис. 8-5). Синтез цепи в направлении от 3' к 5' непрерывен — последовательно добавляется по одному нуклеотиду, тогда как синтез другой цепи — прерывистый, она строится из ряда коротких фрагментов, которые затем соединяются, образуя непрерывную цепь; при этом каждый фрагмент синтезируется в обратном направлении. В настоящее время у бактерии Escherichia coli обнаружено более 14 различных ферментов, которые участвуют в тех или иных этапах процесса репликации ДНК, а у эукариот, удвоение генетического материала происходит еще сложнее.