СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 11. БАКТЕРИИ
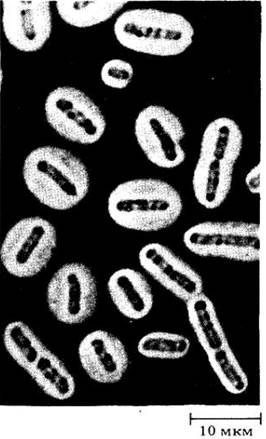
Бактерии относятся к прокариотам; это самые простые, мельчайшие и наиболее распространенные организмы (рис. 11-1). Они были единственной формой жизни на Земле по крайней мере в течение 2 млрд. лет. Бактерии настолько отличаются от других живых организмов, что их выделяют в особое царство — Моnеrа. Они обладают разнообразными типами метаболизма, в том числе некоторые из них способны к фотосинтезу. Одну из групп фотосинтезирующих бактерий, так называемые сине-зеленые водоросли, или цианобактерии, раньше относили к «водорослям», однако в настоящее время их рассматривают как специфическую группу бактерий.
Рис. 11-1. Это клетка обычной кишечной палочки (Escherichia coli); во время приготовления препарата из клетки вьшли нити ДНК. Бактериальная хромосома состоит из одной гигантской молекулы ДНК, содержащей около 3 млрд. пар оснований. Небольшие кольцевые ДНК, видимые слева, — это плазмиды, содержащие 5000 пар оснований. (Черные пятна — это дыры в пленке, с которой была напечатана фотография; рассеянные белые точки — возможно, частички пыли или клетки.)

Бактериальные клетки не имеют ядра, покрытого ядерной оболочкой, и набора хромосом, характерного для эукариот. У бактерий отсутствует половой процесс размножения, хотя случайная рекомбинация генетического материала может иметь место. Бактерии в отличие от эукариот никогда не были многоклеточными организмами, тем не менее некоторые из них образуют нити или скопления из нескольких клеток. Подобные структуры образуются в результате неполного разделения клеточных оболочек после деления клеток; иногда группа клеток окружается общей слизистой капсулой, или оболочкой. Плазмодесмы между такими клетками встречаются чрезвычайно редко (только у нескольких видов цианобактерий).
Бактерии лишены покрытых мембраной органелл, но имеют другие структуры, выполняющие их функции. Плазматические мембраны бактерий часто образуют внутри клеток складки (рис. 11-2), которые увеличивают поверхность прикрепления ферментов и пространственно разделяют ферментативные реакции. У одних фотосинтезирующих бактерий пигменты локализованы на внутренних мембранах, у других они находятся в отдельных округлых тельцах, называемых хроматофорами. Клеточные оболочки большинства бактерий содержат мурамовую кислоту, которая отсутствует у эукариот.
Общая характеристика бактерий
Диаметр большинства бактериальных клеток равен 1 мкм (иногда 0,1 мкм), в длину некоторые бактерии достигают 10 мкм (редко 30 мкм). Хотя каждая бактериальная клетка очень мала, тем не менее общая масса бактерий, обитающих на Земле, превышает массу всех других живых организмов вместе взятых. Сейчас известно около 2500 видов бактерий.
В древнейших осадочных породах Гренландии, имеющих возраст 3,9 млрд, лет, ископаемые бактерии не обнаружены. Они найдены в Западной Австралии (см. рис. 1-2) и в Южной Африке (возраст пород 3,5 млрд. лет). Органические тельца, напоминающие цианобактерии, обнаружены в южноафриканских породах возрастом около 3,2 млрд. лет. Химический анализ ископаемых отложений, а также отпечатки указывают на то, что фотосинтез возник около 3,3 млрд, лет назад. С другой стороны, первые эукариоты появились около 1,5 млрд. лет назад. Ископаемые бактерии были найдены при исследовании ультратонких срезов пород с помощью электронного микроскопа только в 50-х годах.
Рис. 11-2. Одиночная клетка бактерии Рrochloron; хорошо видна обширная мембранная структура. Рrochloron — фотосинтезирующая бактерия, содержащая хлорофиллы а и b и каротиноиды. — пигменты, обнаруженные у зеленых водорослей и высших растений

Местообитания бактерий чрезвычайно разнообразны; в силу специфики обмена веществ некоторые группы способны выжить там, где не может существовать ни один другой организм. Некоторые бактерии являются облигатными анаэробами, т. е. они живут только в отсутствие кислорода. Другие — факультативные анаэробы — могут существовать без кислорода, но лучше развиваются в его присутствии. При этом в процессе дыхания выделяется больше энергии, чем в процессе брожения (см. гл. 6).
Бактерии обнаружены в самых удивительных местах; они действительно распространены по всему земному шару. Например, бактерия Thermoanaerobacter ethanolicus, найденная в горячих источниках Йеллоустонского национального парка (США), может выдержать температуру, превышающую 78°С. Известны бактерии, обитающие при высоком давлении около глубоководных вулканических кратеров при температуре выше 360°С. Бактерии, живущие там, можно культивировать в лаборатории только при высоком давлении; при этом некоторые формы активно растут и удваиваются в числе при температуре 250°С в течение 40 мин! (Столь высокая температура водной среды может поддерживаться длительное время только при высоком давлении, т. е. на дне океана.) Другие глубоководные бактерии, например, обитающие в кишечнике морских животных, для роста и развития тоже нуждаются в высоком давлении.
Разнообразные бактерии обнаружены в Антарктиде, например, в образцах пород и льда, взятых с глубины более 430 м. Эти бактерии имеют возраст не менее 10 000 лет (а возможно, миллион лет). От — 7 °С до — 14°С они пребывают в состоянии покоя, но становятся активными при повышении температуры. Подобные примеры свидетельствуют о том, что некоторые бактерии способны чрезвычайно долго пребывать в состоянии, при котором приостановлены процессы жизнедеятельности.
Недавно ученые исследовали пределы выносливости бактерий, чтобы понять, может ли существовать жизнь на других планетах с различной атмосферой. Например, полагали, что в щелочной атмосфере Юпитера жизнь невозможна, однако некоторые бактерии из долины Ливермор в Калифорнии способны расти и размножаться в щелочной среде при рН = 11,5. Другие бактерии могут выдержать не только высокие концентрации щелочи, но и аммония, т. е. условия, напоминающие атмосферу Венеры. С этой точки зрения можно допустить существование бактерий на других планетах. Однако большинство бактерий не выживает в кислой среде, вот почему уксус, т. е. разбавленная уксусная кислота, хорошо защищает пищу от бактериального заражения.
Бактерии играют немаловажную роль в функционировании экосистем земного шара. Некоторые бактерии — автотрофы и поэтому вносят большой вклад в круговорот углерода; некоторые из них, выращиваемые экспериментально, могут быть коммерческими источниками белка. Способность бактерий к фиксации атмосферного азота имеет решающее экологическое значение. Гетеротрофные бактерии, подобно грибам, являются редуцентами. Благодаря им вещества, утилизированные когда-то живыми организмами, высвобождаются и становятся доступными для последующих поколений. В 1 г плодородной почвы сельскохозяйственных угодий может содержаться 2,5 млрд, бактерий, 400 000 грибов, 50 000 водорослей и 300 000 простейших. На всех этапах круговорота элементов в экосистемах земного шара бактерии играют существенную роль.
Бактерии в изобилии обитают в море; например, установлено, что 90% всей биомассы океана составляют организмы, имеющие размер менее 10 мкм, т. е. бактерии. В зависимости от географического расположения моря от 20 до 60% первичной продукции органического вещества проходит через свободноживущих гетеротрофных бактерий.
Бактерии могут разлагать различные органические вещества. Именно поэтому интенсивно исследуется их способность разлагать нежелательные синтетические вещества, такие, как пестициды, красители, а также нефть (причем не важно, поступили ли эти вещества в окружающую среду). Например, найлон, впервые выпущенный в 1939 г., разлагается бактерией рода Flavobacterium, у которой за такой короткий срок, очевидно, возникли два новых фермента. Некоторые бактерии разлагают пестициды столь быстро, что ограничивают их воздействие на вредителей. Другие бактерии, по-видимому, могут быть использованы для извлечения нефти из различных пород.
Кроме экологической роли бактерий необходимо отметить, что многие из них являются возбудителями очень серьезных заболеваний человека и животных, включая туберкулез, холеру, сибирскую язву, гонорею, дифтерит, столбняк; они вызывают болезни растений, нанося экономический урон. Только в США известно более 200 видов бактерий, которые признаны патогенами растений. Например, бактериальный ожог (рис. 11-3) поразил тысячи грушевых деревьев, распространившись по всем США, начиная с восточного побережья, куда был завезен в 1880 г. К 30-м годам эта болезнь практически уничтожила все грушевые посадки. Сейчас выращивание груш в США ограничено несколькими районами.
Рис. 11-3. Бактериальный ожог, вызываемый Erwinia amylovora (см. рис. 11- 10), —очень распространенное заболевание груш, яблонь и многих плодовых деревьев семейства розоцветных (Rosaсеае). Вначале бактерия поражает ветки этих растений, а затем вызывает некроз ствола (показано больное грушевое дерево). Зараженные деревья довольно быстро погибают. Бактериальная природа данного заболевания была установлена в конце XIX в.

Бактерии используются как коммерческие источники разнообразных антибиотиков, таких, как тиротрицин, бацитрацин, субтилин и полимиксин В. Многие бактерии необходимы для получения лекарств и других химических соединений, например, уксусной кислоты, аминокислот и ферментов (см. гл. 30). Кроме того, производство большинства сыров предполагает бактериальное сбраживание лактозы до молочной кислоты, которая осаждает белки молока. Те же самые бактерии используются и для получения простокваши; кроме того, молочная кислота играет защитную роль в процессе квашения капусты и огурцов.
На рис. 11-4 изображена Spirulina — цианобактерия, выращиваемая во многих странах для получения белковой добавки к пище человека и животных.
Такая группа бактерий, как актиномицеты, играет особенно важную роль в производстве антибиотиков, а именно: стрептомицина, ауреомицина, неомицина, тетрациклина. Из нескольких тысяч известных на сегодняшний день антибиотиков две трети продуцируется актиномицетами. Другие актиномицеты, живущие на корнях некоторых растений, фиксируют азот и тем самым играют важную экологическую роль.
Рис. 11-4. А. Спиралеобразная нитчатая цианобактерия спирулина (Spirulina), широко культивируемая как источник белка. Б. Водоемы для культивации спирулины около озера Тескоко близ Мехико. В. Сбор влажной биомассы спирулины. Впервые спирулину стали использовать в пищу испанские конкистадоры в Мехико около 1521 г.; паста из нее содержит больше белка, чем соя, причем этот белок сбалансирован по содержанию аминокислот. Ежедневный выход белка в 10 раз выше, чем белка пшеницы или сои; продуктивностью спирулины и вызвано ее культивирование

Форма бактерий
Бактерии отличаются разнообразием форм и клеточной организации (рис. 11-5). Прямые, палочковидные бактерии называют бациллами, сферические — кокками, спиралевидные — спириллами (рис. 11-6). Сферические бактерии после деления могут образовывать пары (диплококки), цепочки (стрептококки) или собираться в грозди (стафилококки). Диплококки вызывают пневмонию, а заражение стафилококком приводит к сепсису и нагноениям.
Рис. 11-5. Некоторые типы бактерий. А. Миксобактерии, или слизистые бактерии, организованы подобно слизевикам (гл. 14). Здесь представлена микрофотография (сканирующий электронный микроскоп) «плодовых тел» Chondromyces crocatus; каждое состоит примерно из 1 млн. клеток. В норме миксобактерии имеют палочковидную форму, они способны передвигаться и образовывать плодовые тела. Б. Спирохеты — спиралевидные бактерии длиной около 500 мкм (для бактерий это внушительный размер). Они способны к волнообразному движению. Treponema pallidum, показанная здесь, — возбудитель сифилиса. В. Актиномицеты обитают в почве и разлагают органический материал. Особым запахом плесени влажная почва обязана именно им. Streptomyces fradiae, представленный на фотографии, является коммерческим источником антибиотика неомицина. Г. Крупные скользящие бактерии имеют нитевидную форму; они способны к ритмическим сокращениям, которые приводят к скользящим движениям и периодическим изменениям формы клеток. В клетках Beggiatoa, показанной здесь, хорошо заметны гранулы серы, образовавшейся в результате окисления сероводорода (при этом запасается энергия)

Рис. 11-6. Три основные формы бактерий: А — бациллы (палочковидные); Б — кокки (сферические); В — спириллы (изогнутые). Морфология клетки — относительно постоянный признак большинства видов бактерий. К бациллам относятся, например, Clostridium tetani — возбудитель столбняка, а также обыкновенная кишечная палочка
— Escherichia соli. Многие бациллы вызывают болезни растений, например, бактериальный ожог груш и яблонь (возбудитель — Erwinia amylovora), бактериальный вилт картофеля, томатов и бананов (возбудитель — Pseudomonas solanacearum). К коккам относятся Diplococcus pneumoniae, вызывающий бактериальную форму пневмонии; Streptococcus lactis — бактерия для квашения молока; Nitrosococcus nitrosus, почвенная бактерия, окисляющая аммиак до нитритов. Спириллы менее распространены; они имеют форму cnupajtu

Бациллы, как правило, — это одноклеточные формы; но иногда они образуют нити, поскольку делятся поперек. Нити напоминают грибные, поэтому к наименованию бацилл часто добавляют префикс мико- (от греч. «гриб»). Например, Mycobacterium tuberculosis — палочковидная бактерия — образует нити в культуре, но не в организме хозяина; другие актиномицеты имеют аналогичную морфологию (рис. 11-5, Г). Цианобактерии, разделившись, тоже способны образовывать нитевидные структуры (рис. 10-4, Б, В, 11-7).
Рис. 11-7. Три распространенных рода цианобактерий и родственная форма. A. Oscillatoria размножается только участками нити. Б. Calothrix (Gloeotric hia), нитевидная форма с базальными гетероцистами. Gloeotrichia образует акинеты — крупные клетки с прочной внешней оболочкой. В. Желеобразные «шары» Nostoc commune; каждый шар содержит сотни нитей, обитает обычно в пресной воде. Г. Thiothrix — род бактерий, не имеющих хлорофилла, но близких к цианобактериям. Тhiothrix получает энергию, окисляя H2S. Цепочки клеток, содержащих гранулы серы, прикреплены к субстрату (в центре) и образуют характерную розетку

Клеточная оболочка
В начале 80-х годов прошлого века датский микробиолог Ганс Кристиан Грам, работая в берлинском морге, окрасил инфицированные ткани красителем, сходным с применяемым сейчас кристаллическим фиолетовым, и обнаружил в них бактерии, вызывающие пневмонию. Многие другие бактерии также становились пурпурными при окрашивании этим красителем, однако Грам был разочарован, что не все. Впоследствии открытие Грама во многом способствовало пониманию структуры бактериальных клеточных оболочек, различающихся у разных групп бактерий. Сейчас используют следующую процедуру: клетки окрашивают кристаллическим фиолетовым, затем обрабатывают разбавленным раствором йода (который образует с красителем нерастворимый комплекс), далее промывают спиртом. Спирт удаляет краситель из оболочек грамотрицательных бактерий, грамположительные бактерии, наоборот, удерживают краситель.
Позднее обнаружилось, что матрикс бактериальных оболочек состоит из дисахаридов, связанных короткими цепочками аминокислот (пептидами). Оболочка грамположительных бактерий, толщина которой колеблется от 15 до 80 нм в зависимости от вида бактерии, содержит только такие макромолекулы пептидогликана. В оболочках грамотрицательных бактерий слой пептидогликана окружают липополисахариды (полисахаридные цепи с прикрепленными к ним липидами). Оболочки грамотрицательных бактерий имеют толщину не более 10 нм.
Грамположительные и грамотрицательные бактерии различаются по реакции на антибиотики. Например, пенициллин блокирует образование пептидных мостиков в оболочке грамположительных бактерий, и потому клетка по мере роста разрушается. Действию пенициллина на оболочки грамотрицательных бактерий препятствует слой липополисахаридов. Широко используемый антибиотик актиномицин нарушает синтез белка, связываясь с двойной спиралью ДНК. Молекула актиномицина легко проходит через оболочку грамположительных, но не грамотрицательных бактерий. Таким образом, для последних следует использовать другие антибиотики, например, эритромицин.
Бактериальные клетки обладают способностью к адгезии за счет особого слоя переплетенных полисахаридных нитей, называемого гликокаликсом. Хотя в лабораторных культурах бактерии не образуют гликокаликса, в природных условиях он играет ключевую роль на начальном этапе, а затем и в процессе распространения бактериальной инфекции. Именно с помощью гликокаликса бактерии прикрепляются к различным субстратам.
Благодаря ферментативным реакциям, протекающим в гликокаликсе, некоторые бактерии могут поселиться на неповрежденной поверхности зубной эмали или других, кажущихся устойчивыми субстратах.
Многие бактерии образуют желатинообразную капсулу, которая, вероятно, откладывается протопластом (рис. 11-8). Подобно оболочке, капсула состоит из полисахаридов, однако она непрочно связана с бактерией и может быть удалена отмыванием.
Рис. 11-8. Микрофотография отдельных клеток Bacillus megaterium, диспергированных в китайской туши. Видны капсулы (прозрачный ободок)
Цитоплазма
Цитоплазма бактерий, как и других клеток, окружена плазматической мембраной, на внутренней поверхности которой локализованы многочисленные ферменты. В цитоплазме обнаруживают большое количество рибосом и гранулярных включений, а также один или два участка, где концентрируется ДНК (как правило, два, поскольку деление клетки отстает от деления генетического материала). Каждый из них содержит одну молекулу кольцевой двухцепочечной ДНК, длина которой в 700 или 1000 раз превышает длину самой клетки.
Жгутики и фимбрии
Некоторые бактерии имеют очень тонкие прочные спиральные жгутики, в несколько раз более длинные, чем сами клетки. Жгутики совершают быстрые вращательные движения и способствуют движению бактерий. Бактерии для своих размеров передвигаются очень быстро, за одну секунду они преодолевают расстояние, равное примерно 20 диаметрам самой бактериальной клетки. Вращательное движение нехарактерно для эукариот, и поэтому постоянное вращение бактериальных жгутиков представляет собой уникальное явление.
Жгутики бактерий напоминают (9+2)-жгутики эукариот только внешне. Они длинные (3 — 12 мкм), тонкие и волнообразно изогнутые. Диаметр жгутиков обычно равен 10 — 12 нм, поэтому их нельзя увидеть в световой микроскоп. У некоторых бактерий жгутики равномерно распределены по всей поверхности клетки, у других — прикреплены к одному или обоим концам (рис. 11-9, 11-10). Каждый бактериальный жгутик состоит из одной жесткой молекулы белка флагеллина, которая выходит из «муфты» в клеточной оболочке и связана со сложным вращающим механизмом. Механизм состоит из кольца, вращаемого за счет электрического потенциала, и лежащего внутри другого, большего по размеру неподвижного кольца.
Рис. 11-9. Жгутики Pseudomonas marginalis — бактерии, широко распространенной в почве. Вызывает мягкую гниль корнеплодов и других овощей
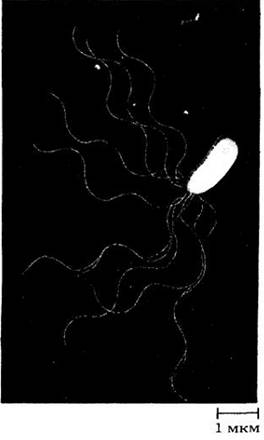
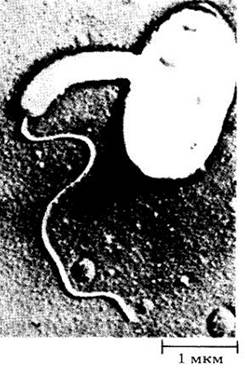
Рис. 11-10. Бактерия Bdellovibrio bacteriovorus паразитирует на других бактериях. Часто встречается в почве, сточных водах; двигается с помощью заднего жгутика. Показано, как паразитическая клетка атакует палочковидную Erwinia amylovora (справа на фотографии) — возбудителя бактериального ожога груш и яблонь
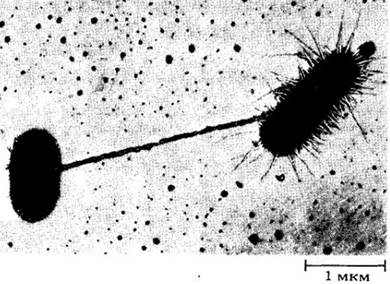
Фимбрии (ворсинки) короче (до нескольких микрометров) и прямее, чем жгутики; их диаметр составляет около 7,5 — 10 нм (рис. 11-11). Фимбрии характерны главным образом для грамотрицательных бактерий. Они состоят из белка, отличного от белка жгутиков, и имеют иную структуру. Полые ворсинки образуются на бактериальных клетках во время конъюгации, однако их точная функция пока неизвестна. Возможно, они переносят ДНК или способствуют соединению клеток. В большинстве случаев ворсинки помогают бактериям прикрепляться к определенным мембранам. Лучшее понимание роли фимбрий может помочь в борьбе с бактериальными болезнями.
Рис. 11-11. Фимбрии Escherichia coli. Полая фимбрия связывает две клетки; у правой видны многочисленные фимбрии
Специфические особенности цианобактерий
Некоторые фотосинтезирующие бактерии, известные как цианобактерии, образуют нити длиной до одного метра и более и способны нарастить большую биомассу. Некоторые цианобактерии являются одноклеточными, другие соединены в цепочки, и очень немногие образуют округлые или неправильной формы колонии (рис. 10-4, Б, В; 11-7, 11-12,11- 13). Любая клетка, кроме гетероцисты, может разделиться; образующиеся при этом клетки способны дать начало новым колониям. Подобно другим нитчатым и колониальным формам бактерий, цианобактерии прикреплены друг к другу только оболочками или желатиновыми капсулами, поэтому каждая клетка действительно ведет независимую жизнь; однако у некоторых колониальных цианобактерий обнаружены плазмодесмы.
Рис. 11-12. А. Клетка цианобактерии Anabaena azollae. Электронная микрофотография демонстрирует внутреннее строение. Желатиновая капсула разрушилась в процессе приготовления препарата. Данный микроорганизма тесно связан с плавающим водным папоротником A zolla и фиксирует азот (см. рис. 10-8, А и гл. 26). Б. Китайский рабочий собирает Azolla из водоема, чтобы культивировать его на рисовых полях

Цианобактерии, обитающие в поверхностном слое воды пресных и морских водоемов, образуют сообщества микроскопических организмов, называемых планктоном; как правило, эти микроорганизмы имеют прозрачные, неправильной формы структуры — газовые вакуоли. Эти вакуоли регулируют плавучесть организмов и позволяют им оставаться в толще воды. Когда цианобактерии теряют способность регулировать свою плотность, например, при резких перепадах температуры или нарушениях кислородного обмена, они всплывают на поверхность и вызывают «цветение» воды. При этом некоторые цианобактерии выделяют химические вещества, токсичные для других организмов и приводящие к их массовой гибели.
Движение бактерий
Несмотря на простоту организации, бактерии могут реагировать на определенные раздражители, двигаясь в направлении увеличивающейся концентрации пищи или кислорода. Подвижные бактерии, такие, как Escherichia coli, способны перемещаться к химическим агентам или от них, вращая свои жгутики; каждая клетка Е. coli имеет от шести до восьми жгутиков. Бактерии обладают специфической чувствительностью к различным питательным веществам, например, сахарам. После того как клетки получат сигнал о «привлекающих» (аттрактантах) или «отталкивающих» (репеллентах) веществах, они могут выбрать нужное им направление движения.
Джулиус Адлер и его коллеги из Висконсинского университета определили, что Е. coli имеет около 20 различных хеморецепторов белковой природы: из них 12 — на аттрактанты и 8 — на репелленты. Белки, ответственные за связывание сахаров, например, галактозы, мальтозы, рибозы, расположены в периплазматическом пространстве между клеточной оболочкой и плазматической мембраной. Другие хеморецепторы (также белки) находятся на плазматической мембране. Информация, получаемая хеморецепторами, неизвестным способом передается жгутикам, и те приходят в движение.
Отдельные клетки Е. coli двигаются быстро, совершая при этом криволинейные «пробежки», каждая из которых длится около секунды. Серия пробежек часто прерывается поворотами на месте, «кувырканиями», которые длятся одну десятую долю секунды. После каждого из них бактерия начинает следующую пробежку в другом направлении. Частота «кувырканий» контролирует длину пробега и, следовательно, общее направление движения. Если бактерия плывет в «верном» направлении, ее перевороты ингибированы, если — в «ошибочном», то частота «кувырканий» увеличивается.
Конечная цель подобных перемещений — миграция бактерий к источнику аттрактантов или от источника репеллентов. Аттрактанты вызывают вращение жгутиков против часовой стрелки, а репелленты — наоборот, по часовой стрелке. Таким образом, «пробежки» совершаются за счет вращения жгутиков против часовой стрелки, а «кувыркания» — за счет вращательного движения по часовой стрелке. При длительном движении жгутики работают вместе и собираются в пучок на заднем конце клетки, несмотря на то что расположены практически по всей поверхности. Когда жгутики вращаются по часовой стрелке, пучок распадается, и клетка переворачивается. В этом случае жгутики вращаются независимо. Реакции таких бактерий на внешние воздействия очень сложны (если иметь в виду прокариотический уровень организации) и предвосхищают гораздо более сложные реакции эукариот.
Некоторые группы бактерий по-другому реагируют на условия внешней среды. Например, некоторые бактерии, обитающие в воде, содержат кристаллы магнетита (Fе3O2). Они способны ориентироваться в магнитном поле земного шара и постоянно плывут в одном направлении. Те микроорганизмы, которые обнаружены в Северном полушарии, движутся на север, следуя линиям магнитного поля, которые направлены под углом к горизонту вглубь водоема. В Южном полушарии аналогичные бактерии плывут к югу. В обоих случаях бактерии углубляются в толщу воды; возможно, магнитотаксис способствует поиску пищи.
Совершенно иной способ движения обнаружен у нитчатых цианобактерий и у бактерий, лишенных жгутиков (см. рис. 11-5). Движение этих микроорганизмов представляет собой скольжение, но может включать и вращение вдоль продольной оси клетки. Короткие сегменты, отчлененные от колонии цианобактерий, могут скользить со скоростью порядка 10 мкм/с. Движению способствуют выделение слизи через поры клеточной оболочки и образование сократительных волн на ее внешней поверхности. Некоторые цианобактерии совершают конвульсивные движения.
Генетика бактерий
Клеточное деление
Основной способ размножения бактерий — бесполый; каждая клетка увеличивается в размерах и делится надвое. Во время деления плазматическая мембрана и клеточная оболочка впячиваются, в конце концов перешнуровывая клетку пополам (рис. 11-13). Вновь образованная клеточная оболочка толще прежней, она быстро расслаивается в центре, обеспечивая каждую дочернюю клетку новой клеточной оболочкой. Цепочки бактерий образуются в том случае, если клеточная оболочка разделилась не полностью; такие цепочки могут разрываться на многоклеточные фрагменты, например, гормогонии у цианобактерий.
Рис. 11-13. Деление Аnabaena. Электронная микрофотография показывает цепочку клеток, удерживаемых вместе не разделившейся клеточной стенкой; слева показано деление отдельной клетки. Левая клетка цепочки — гетероциста. Тип деления, при котором клетки перешнуровываются, характерен для многих организмов, кроме растений и некоторых родов водорослей, у которых при делении образуется клеточная пластинка

Полагают, что единственная кольцевая двухцепочечная молекула ДНК, несущая генетическую информацию, прикреплена в одной точке к внутренней поверхности плазматической мембраны. После того как ДНК удвоится, две идентичные кольцевые молекулы остаются соединенными бок о бок с плазматической мембраной. Новая плазматическая мембрана и клеточная оболочка образуются между двумя точками прикрепления ДНК по мере деления клетки; в итоге мембрана впячивается внутрь между двумя молекулами ДНК, а каждая дочерняя клетка обеспечивается идентичной молекулой ДНК (рис. 11-14).
Рис. 11-14. Схематическое изображение, позволяющее понять способ прикрепления бактериальной ДНК к плазматической мембране, приводящий к распределению ДНК между дочерними клетками. На самом деле (см. рис. 11-1) ДНК намного длиннее

Некоторые бактерии образуют толстостенные эндоспоры, которые устойчивы к нагреванию и обезвоживанию. Эндоспоры образуются путем деления протопласта бактериальной клетки на две или более частей. Вокруг обособившегося участка протопласта, содержащего ДНК, образуется плотная споровая оболочка. Эндоспоры могут прорасти через десятки, даже сотни лет. Стойкие споры цианобактерий называют акинетами. Акинеты принципиально отличаются от эндоспор тем, что представляют собой увеличенные вегетативные клетки, вокруг которых сформировалась плотная оболочка (рис. 11-15). Актиномицеты образуют цепочки эндоспор на концах нитей.
Рис. 11-15. Нити Аnаbаenа — азотфиксирующей цианобактерии, — состоящие из бочкообразных клеток, окруженных студенистым матриксом. Фиксация азота происходит в особых клетках, называемых гетероцистами. Как и Calothrix (см. рис. 11-7, Б), Аnabaena образует окинеты. Электронные микрофотографии представлены на рис. 11-12, 11-13

Генетическая рекомбинация
Генетическая рекомбинация у бактерий происходит в результате переноса участка молекулы ДНК от одной бактериальной клетки к другой. Внесенный фрагмент ДНК может действовать совместно с ДНК клетки-реципиента и участвовать в образовании информационной РНК. В другом случае он встраивается в кольцевую молекулу ДНК и передается при делении дочерней клетке вместе с остальным генетическим материалом. Рекомбинация может происходить в результате конъюгации (обмена фрагментами ДНК при прямом контакте клеток), трансдукции (передачи ДНК от клетки- донора клетке-реципиенту с помощью вируса) или трансформации (передачи свободной растворимой ДНК без всякого межклеточного контакта и каких-либо переносчиков).
Почти все бактерии наряду с большими кольцевыми молекулами ДНК, называемыми бактериальными хромосомами, имеют небольшие кольцевые ДНК — плазмиды (см. рис. 11-1). Некоторые плазмиды встраиваются в хромосомы и удваиваются вместе с ними. У некоторых штаммов бактерий конъюгация и обмен плазмидами происходят довольно часто (см. рис. 11-11), что способствует передаче от одного штамма бактерий к другому таких признаков, как устойчивость к антибиотикам. Процесс рекомбинации широко используется в генной инженерии (см. гл. 30).
Генетический материал может передаваться от одного штамма к другому путем трансдукции. Бактериальные вирусы, называемые бактериофагами (см. рис. 12-1), могут захватывать небольшие участки бактериальных хромосом и переносить их в другую бактерию. ДНК вируса вместе с ДНК бактерии могут встроиться в хромосому нового штамма- хозяина.
Рис. 11-16. Отбор мутантных бактерий методом реплик (отпечатков). А. Исходная чашка. Колонии переносились с чашки А на чашку Б и чашку В, с помощью стерильного бархатного диска, который прижимали к агару; при этом часть клеток приставала к ворсинкам бархата. Затем бархат прижимали к незасеянному агару других чашек, и на них воспроизводилась вся картина колоний исходной части. В первых двух чашках виден рост колоний на полной среде, а в чашке В не хватает ряда факторов роста. При этих условиях колонии, указанные стрелками на чашке Б, не растут на третьей чашке В; они являются мутантами, которые уже возникли, но еще не были идентифицированы в чашке А

В другом случае генетическая рекомбинация происходит в результате трансформации. Первые опыты по трансформации бактерий были проведены на Diplococcus pneumoniae — возбудителе пневмонии. Эти бактерии образуют два типа колоний: шероховатые (R) и гладкие (S). Бактерии, образующие S-колонии, имеют капсулы и вызывают пневмонию. В 1928 г. Фредерик Гриффит показал, что можно трансформировать безвредный штамм R в вирулентный штамм S, если смешать штамм R с убитыми высокой температурой бактериями штамма S (см. рис. 8-2). При разрушении бактериальных клеток с помощью химических веществ или нагреванием фрагменты ДНК высвобождаются и могут проникнуть в другие клетки. Таким образом, продемонстрировав существование генетически активного материала, участвующего в трансформации, Гриффит, по сути, первым доказал генетическую роль ДНК. Сейчас известно, что процесс трансформации происходит у многих бактерий.
Мутации — это еще более важный источник изменчивости бактерий, чем генетическая рекомбинация (рис. 11-16). Вычислено, что для определенного гена на 107 клеток приходится одна мутантная. Отдельная клетка Escherichia coli имеет приблизительно 5000 генов. Таким образом, в культуре этих бактерий на 2000 клеток имеется одна клетка, которая мутантна по любому гену; в культуре 0,05% клеток будут иметь мутантный фенотип при каждом клеточном делении. В культуре из 109 клеток (потомство 30 делений одной бактерии) частота мутаций составит 30x0,05, т. е. 1,5%. Бактерии размножаются очень быстро. При оптимальных условиях популяция клеток Е. coli может удвоиться за 12,5 мин; таким образом, число мутантных клеток быстро растет. Большая скорость размножения бактерий и высокая частота мутаций обусловливают удивительную способность этих организмов к адаптации.
Метаболизм бактерий
Гетеротрофы
Большинство бактерий — гетеротрофы, они не способны синтезировать органические соединения из простых неорганических, а должны получать их в готовом виде. Самая большая группа гетеротрофных бактерий — это сапробионты. Они «питаются» мертвым органическим материалом. Сапробионты бактерии и грибы ответственны за разложение и кругооборот органического вещества в почве; многие образуемые при этом соединения имеют специфический запах.
Фотосинтезирующие бактерии
Выделяют по крайней мере пять групп фотосинтезирующих бактерий: цианобактерии, зеленые серобактерии, пурпурные серобактерии, пурпурные несерные бактерии и Prochlorоn.Подобно растениям, фотосинтезирующие бактерии содержат хлорофилл. Цианобактерии и Prochloron имеют хлорофилл а аналогично всем фотосинтезирующим эукариотам. Хлорофиллы других фотосинтезирующих бактерий несколько отличаются от хлорофилла а, но структура их в принципе одинакова (см. рис. 7-8).
Окраска перечисленных групп бактерий обусловлена вспомогательными пигментами, необходимыми для фотосинтеза. У двух групп пурпурных бактерий это желтые и красные каротиноиды; у цианобактерий, кроме того, обнаружен синий пигмент фикоцианин и красный — фикоэритрин. Процессы фотосинтеза у цианобактерий и эукариот происходят сходным образом. В цитоплазме цианобактерий находятся многочисленные мембраны, часто расположенные параллельно друг другу, и большое количество рибосом. Эти мембраны являются фотосинтетическими тилакоидами, аналогичными тилакоидам хлоропластов. В клетках цианобактерий, однако, они не организованы в хлоропласты. Основным запасным углеводом цианобактерий, как и других бактерий, является гликоген.
У зеленых и пурпурных серобактерий соединения серы играют в процессе фотосинтеза ту же роль, что и вода у организмов, содержащих хлорофилл а, т. е.
![]()
Как обсуждалось в гл. 7, изучение процесса фотосинтеза у пурпурных серобактерий помогло К. Б. ван Нилю составить общее уравнение этого процесса:
![]()
где Н2А — обобщенный донор водорода.
У пурпурных несерных бактерий донорами водорода для фотосинтетических реакций служат спирты, жирные кислоты и кетокислоты.
Фотосинтезирующие серные бактерии нуждаются в сероводороде или других соединениях серы, поэтому обитают в среде, содержащей большое количество разлагающего органического материала со специфическим запахом серы. Сера может накапливаться в бактериальных клетках про запас.
Halobacterium halobium — пурпурная несерная бактерия — очень своеобразный организм. Она обитает в концентрированном растворе соли на прогретых солнцем местах вблизи соленых водоемов. В плазматической мембране Halo- bacterium содержатся включения пурпурного пигмента, напоминающего родопсин — зрительный пигмент человеческого глаза. Эти включения являются активными светозависимыми «насосами», выкачивающими протоны из бактериальной клетки. Аналогичный процесс происходит при окислительном фосфорилировании в митохондриях; из всех свободноживущих форм бактерий пурпурные несерные бактерии наиболее близки к митохондриям, которые, возможно, и произошли от этой группы бактерий. Этот процесс, вероятно, приводит к образованию АТР с использованием фермента АТРазы, находящегося в тех участках плазматической мембраны, где нет пигмента. Таким образом, Halobacterium — единственный существующий ныне организм, который не имеет хлорофилла, но способен при этом превращать солнечный свет в химическую энергию, т. е. осуществлять фотосинтез. Halobacterium двигается и обнаруживает фототаксис на фиолетовый свет, подтверждая тем самым идею о том, что его пигмент воспринимает свет с помощью того же механизма, что и родопсин.
Пятая группа фотосинтезирующих бактерий состоит из единственного рода Prochloron, который был открыт в начале 70-х годов Ральфом Льюином из Скриппсковского океанографического института. Prochloron содержит хлорофиллы а и b и каротиноиды, т. е. фотосинтстические пигменты зеленых водорослей и растений (рис. 11-2 и 11-17). Насколько известно, он обитает в симбиозе с асцидиями — колониальными морскими животными, населяющими прибрежные зоны тропических и субтропических морей. Prochloron имеет такой набор пигментов, что его можно было бы отнести к бактериям, от которых, по-видимому, произошли хлоропласты зеленых водорослей (см. гл. 15). С другой стороны, по набору нуклеотидов в рибосомальной РНК (рРНК) Рrochloron больше напоминает цианобактерии, чем хлоропласты зеленых водорослей и растений. Возможная эволюционная роль Рrochloron и его связи с другими организмами активно изучаются.
Рис. 11-17. Prochloron — довольно необычная фотосинтезирующая бактерия, обитающая только в симбиозе с асцидиями. A. Стадия личинки у асцидии рода Didemnum. Показано, как происходит распространение бактерий Рrochloron. Б. Взрослая особь Didemnum. В. Внутри вскрытой асцидии Didemnum видна плотная масса Рrochloron
Хемоавтотрофные бактерии
Хемоавтотрофные бактерии получают энергию, необходимую для осуществления синтетических реакций, путем окисления неорганических веществ (азота, серы, соединений железа и газообразного водорода), которые обеспечивают их энергией подобно свету у фотосинтетических организмов. Однако источник углерода тот же — двуокись углерода. Бактерии, обитающие у глубоководных кратеров при температуре выше 360°С, тоже хемосинтетики. Они получают энергию, превращая сульфид водорода в серу, и, кроме того, обеспечивают энергией целое сообщество организмов, существующих в полной темноте океанических глубин.
Архебактерии
Одна группа хемоавтотрофных бактерий, которая привлекла в последние годы большое внимание, относится к метанобразующим бактериям. Эти бактерии — строгие анаэробы; они обитают в желудочно-кишечном тракте жвачных животных, в сточных водах, болотах и в глубине моря. Большинство запасов природного газа образовано в далеком прошлом благодаря метаболической деятельности метанобразующих бактерий. Некоторые из них образуют метан из двуокиси углерода и водорода, при этом получая энергию; другие способны восстанавливать элементарную серу с образованием сероводорода.
Метанобактерии отличаются большим морфологическим разнообразием. Однако недавно Карлом Уозом и его коллегами из Иллинойсского университета было показано, что различные формы метанобактерий имеют гомологичные последовательные рРНК, что свидетельствует об их родстве. Удивительным оказался факт, что эти последовательности оснований резко отличаются от таковых в рРНК других бактерий и эукариот. По набору нуклеотидов рРНК цианобактерий, например, ближе к Escherichia coli и актиномицетам, чем к метанобразующим бактериям. Клеточная оболочка метанобактерией в отличие от всех других не содержит мурамовой кислоты; кроме того, они характеризуются иным типом метаболизма. В 1984 г. было опубликовано сообщение, что один род архебактерий — Methanosarcina — способен фиксировать атмосферный азот. Это довольно интересное открытие, поскольку представители различных родов, обладающие этим свойством, немногочисленны.
На основании изложенных фактов было высказано предположение, что метанобактерии появились на Земле около 3 млрд. лет назад, когда атмосфера была бескислородной, но обогащенной СO2 и Н2. Сейчас они обитают только в определенных специфических условиях. Отличия метанобактерий от других групп настолько велики, что высказано предложение отнести их к отдельному царству — архебактерии. Как уже обсуждалось, Halobacterium вместе с некоторыми другими бактериями, обитающими при высокой температуре и в кислой среде, по ряду признаков напоминает метанобактерии. По этой причине, может быть, все эти организмы стоит отнести к царству архебактерий.
Экология бактерий
Почвенные бактерии
Различные группы микроорганизмов участвуют в отдельных этапах разложения и круговорота веществ, происходящих в почве; эти естественные сообщества организованы довольно сложным путем (рис. 11-18). Например, штамм Streptococcus, неустойчивый к пенициллину, защищается пенициллиноустойчивым штамом рода Bacteroides, который так эффективно разрушает пенициллин, что антибиотик не в состоянии подавить рост Streptococcus. Многие бактерии и грибы разлагают углеродсодержащие соединения и выделяют в атмосферу СO2. Наиболее важные органические вещества растительного происхождения — это целлюлоза, лигнин, пектины, крахмал и сахара. Установлено, что более 90% СO2образуется в биосфере в результате деятельности бактерий и грибов.
Рис. 11-18. Данная фотография иллюстрирует сложные взаимоотношения между почвенными микроорганизмами; показаны бактерии, растущие на агаре с пенициллином. В центре хорошо видна плотная колония Staphylococcus epidermides. Данный штамм устойчив к пенициллину, поскольку образует фермент пенициллиназу и расщепляет антибиотик. По обе стороны колонии стафилококка можно видеть крошечные колонии Neisseria gonorrhoeae, возбудителя гонореи. Данный микроорганизм чувствителен к пенициллину, но может расти на том участке среды, где стафилококк разрушил антибиотик

Некоторые микроорганизмы гидролизуют белки до пептидов, которые затем разлагаются до составляющих их аминокислот. Многие используют процесс аммонификации — разложения аминокислот с выделением ионов аммония (NH+4). Аммоний может окислиться до нитрита (NO2) хемоавтотрофной бактерией Nitrosomonas, а нитрит до нитрата (NO-3) — бактерией Nitrobacter. Окисление аммония в нитриты и нитраты называют нитрификацией. Этот процесс идет с выделением энергии, которая используется хемоавтотрофами для восстановления двуокиси углерода до углеводов. Некоторые бактерии способны превращать нитраты в нитриты и в конечном счете в аммоний.
Денитрификация — превращение нитратов в газообразный азот или оксид азота — приводит к уменьшению количества азота в почве. Процесс, обратный денитрификации, называют фиксацией азота; он чрезвычайно важен с биологической точки зрения. Из всех живых организмов только бактерии нескольких родов способны к фиксации атмосферного азота. Наиболее известная из них — это симбиотическая бактерия Rhizobium (гл. 26), которая образует клубеньки на корнях бобовых и некоторых других растений. Актиномицеты участвуют в образовании клубеньков у многих древесных пород, например, ольхи (Alnus), восковницы (Myrica) и цеанотуса (Ceanothus). Актиномицсты, фиксирующие азот, способствуют его накоплению в почве (рис. 11-19). Кроме того, определенные азотфиксирующие бактерии связаны с листьями, где они используют богатый углеводами экссудат растений, снабжая их в свою очередь азотом в доступной форме. И свободноживущие формы, включая некоторые цианобактерии, играют важную роль в фиксации азота.
Рис. 11-19. Ольха вступает в симбиоз с азотфиксирующими актиномицетами рода Frankia. Эти взаимоотношения хорошо изучены на примере красной ольхи в Гарвардском лесничестве в Массачусетсе Джоном Торри и его коллегами. А. Сеянцы ольхи выращивались на различных почвенных смесях после инокуляции Frankia. Сеянцы, показанные слева, не получили инокулят Frankia. Сеянцы справа выросли с Frankiа, образовав множество клубеньков. Б. Клубенек на корне ольхи. В. Микрофотография участка корня сеянца ольхи, прокрашенного толлуидиновым синим, где видны клетки коры, содержащие Frankiа. Гифы лежат в центре инфицированных клеток, а на периферии видны расширенные концы гиф, называемые везикулами. Полагают, что в везикулах локализован фермент нитрогеназа, фиксирующий

Хемоавтотрофные бактерии, такие, как Thiobacillus, окисляют серу до сульфатов и тем самым делают ее доступной для растений, которые сами не могут усваивать элементарную серу:
2S + 2Н2O + 3O2 —> 4Н+ + 2SO42-.
Сульфаты накапливаются растениями, и содержащаяся в них сера включается в белки. При распаде белков (обсуждение этого вопроса см. в гл. 26) высвобождаются аминокислоты, в том числе и серосодержащие (рис. 11-20). Некоторые бактерии способны разлагать серосодержащие аминокислоты с выделением сероводорода (Н2S). Сульфаты могут восстанавливаться до Н2S некоторыми почвенными микроорганизмами, например, Desulfovibrio.
Рис. 11-20. Круговорот серы. Благодаря деятельности бактерий и грибов многие элементы, например, углерод, азот, сера и другие, не остаются в составе молекул, в которые они включились в ходе реакций. Почвенные гетеротрофы разлагают органические вещества; продукты распада затем могут включиться в другие биологические циклы. Без круговорота органических соединений все организмы вскоре переполнились бы продуктами своего обмена веществ. Выделение в атмосферу громадных количеств серы при сжигании ископаемого топлива нарушает круговорот серы в природе

Паразитические и симбиотические бактерии
Некоторые гетеротрофные бактерии разлагают органику только при внедрении в живые организмы. К этой группе принадлежат болезнетворные формы, а также непатогенные микроорганизмы. Последние могут оказывать незначительное влияние на жизнедеятельность организма-хозяина или способствовать его процветанию. В частности, если устранить все бактерии, обитающие в желудочно-кишечном тракте человека, как это бывает, например, при длительном лечении антибиотиками, то организм становится более восприимчивым к патогенным бактериям и грибкам. У крупного рогатого скота и других жвачных бактерии гидролизуют целлюлозу до сахарозы, необходимой организму-хозяину; в противном случае эти животные не могли бы использовать в пищу траву и листву, богатые целлюлозой. Другой принципиально важный пример — это взаимовыгодный симбиоз бактерий рода Rhizobium и бобовых (см. гл. 26).
В целом микробы и другие паразиты, вмешиваясь в нормальное течение физиологических процессов заражаемых организмов, вызывают серьезные заболевания. Если смерть наступает слишком быстро, то для данного паразита эта ситуация неблагоприятна; поскольку он не успевает найти другой объект, перезаражения не происходит, и паразит погибает сам.
Болезни человека
Некоторые болезни человека передаются воздушно-капельным путем. Среди наиболее известных заболеваний такого рода — бактериальная пневмония (возбудитель Diplococcus pneumoniae); коклюш (возбудитель Bordetella pertussis) и дифтерит (возбудитель Corynebacterium diphtheriae). Последний микроорганизм выделяет сильно действующий токсин, который быстро разносится по всему организму, повреждая сердечную мышцу, нервные ткани и почки. В настоящее время дифтерит встречается довольно редко, поскольку большинство детей вакцинируют против него. Целый ряд других заболеваний, передающихся воздушным путем, вызывается бактериями рода Streptococcus, например, скарлатина, ревматизм и др. Возбудитель туберкулеза Mycobacterium tuberculosis еще остается причиной смерти многих людей, несмотря на усовершенствование методов диагностики и лечения. Однако число зарегистрированных смертельных случаев в США в середине 80-х гг. уменьшилось по сравнению с 70-ми с 30 до 20 тыс. в год.
Целый ряд других болезней бактериального происхождения передается через пищу и воду. Примерами могут служить брюшной тиф, паратиф (возбудители из рода Salmonella) и дизентерия, вызываемая Shigella dysenteriae. Бруцеллез, называемый бактерией из рода Brucella, опасен как для животных, так и для человека, который заражается через молоко от инфицированной коровы. Пастеризация молока разрушает Brucella, и болезнь стала редкой в тех районах земного шара, где регулярно обрабатывают молоко. Широко распространенная «диарея путешественников» вызывается определенным штаммом Escherichia coli, обитающим в толстой кишке.
В 1976 г. была впервые обнаружена «болезнь легионеров», которая передается через питьевую воду. От этой таинственной болезни легких погибли 34 члена Американского легиона на конференции в Филадельфии. Сейчас известно, что данная болезнь вызывается небольшой жгутиковой палочковидной бактерией, которая относится к роду Legionella. Эти бактерии из теплой воды попадают в организм человека и быстро размножаются в моноцитах, белых клетках крови, играющих немаловажную роль в иммунитете. Сейчас установлено, что «болезнь легионеров» охватила в США около 50 тыс. человек, причем 15 — 20% — с летальным исходом. Виды Legionella относятся к грамотрицательным бактериям, болезнь поддается лечению эритромицином.
Бактерии вызывают гниение продуктов питания и других органических материалов, и некоторые из них чрезвычайно опасны для человека. Употребление пищи, зараженной Clostridiumbotulinum, может привести к смертельному исходу. Эндоспоры этого микроорганизма выживают при кратковременном кипячении и могут прорасти даже в герметичных консервных банках (рис. 11-21). Staphylococcus, отравляющий пищу, довольно распространен, но, к счастью, он менее патогенен. Однако в последние годы появился целый ряд вирулентных штаммов Staphylococcus, которые вызывают опасные инфекции. Многие из этих штаммов устойчивы к пенициллину, некоторые образуют фермент пенициллиназу, разрушающую антибиотик. Staphylococcus часто контактируют с продуцирующими пенициллин грибками или растут вместе с ними, т. е. устойчивость к пенициллину обеспечивает им естественное преимущество (см. рис. 11-18).
Рис. 11-21. Колония бактерии Clostridium botulinum. Вызываемое ею пищевое отравление — ботулизм — наступает при использовании в пищу недоброкачественных консервов. Споры Clostridium botulinum обладают высокой устойчивостью к нагреванию и сохраняют жизнеспособность в пище, которая не была тщательно обработана. Clostridium botulinum, растущий только в отсутствие кислорода, выделяет сильный яд, который накапливается в консервах. Яд разрушается при кипячении в течение 15 мин. Это самый сильный яд из всех известных токсических веществ: 1 г достаточно, чтобы отравить 14 млн. человек. Пищевой ботулизм редко встречается в США; в 1980 — 1983 гг. зарегистрировано 114 случаев

Болезни растений
Многие болезни растений, приносящие большой экономический урон и по существу приводящие к потерям 1/8 части урожая всех культурных растений земного шара, вызываются бактериями. Почти все растения подвержены бактериальному заражению (рис. 11-3, 11-22).
Рис. 11-22. Популяция кактуса карнегия гигантская (Carnegiea gigantea), растущего на высоте 1000 м на южных склонах гор Санта-Каталина близ Тусона о Аризоне, была поражена бактерией Ermnia camegieana после того, как кактусы были ослаблены заморозками в январе 1962 г.

Большинство патогенов растений относится к бациллам (палочковидным формам), многие из них паразитируют в растении-хозяине. Симптомы заболеваний, вызванных патогенными бактериями, чрезвычайно разнообразны, в большинстве случаев это пятна на стеблях, листьях, цветках и плодах (рис. 11-23). Почти всегда они свидетельствуют о бактериальном заражении и чаще всего — близкородственными организмами Pseudomonas и Xanthomonas.
Рис. 11-23. Примеры типичных бактериальных патогенов растений и различные заболевания, которые они вызывают

Многие наносящие экономический ущерб заболевания растений, такие, как ожог, мягкая гниль и вилт (увядание), тоже вызываются бактериями. Ожог характеризуется быстро развивающимися некрозами (мертвые бесцветные пятна) стеблей, листьев и цветков. Бактериальный ожог яблонь и груш может привести к гибели молодых деревьев в течение одного сезона. Эта болезнь вызывается бациллами Erwinia amylovora. Бактериальная мягкая гниль обычно поражает мясистые запасающие части овощей, такие, как клубни (картофель), луковицы, а также сочные плоды — томаты, баклажаны и др. Наиболее разрушительный эффект оказывает мягкая гниль, вызываемая бактериями рода Erwinia.
Бактериальные вилты проводящих тканей поражают только травянистые растения. При этом микроорганизмы внедряются в сосуды ксилемы и размножаются в них, распространяясь с током воды и питательных элементов по всему растению и приводя его к гибели. Вначале бактерии разрушают участки клеточных оболочек сосудов и даже могут разорвать сосуд. Затем они проникают в прилегающие к сосудам клетки паренхимы, где продолжают размножаться. Иногда в тканях растения бактерии образуют полости, заполненные клеточными остатками, смолистыми выделениями и собственно бактериальными клетками. Эти полости через трещины сообщаются с поверхностью стеблей и листьев. Однако, как правило, бактерии не выходят на поверхность до тех пор, пока растение не погибнет. Наиболее распространенными примерами такого рода заболеваний являются: бактериальное увядание люцерны и бобов (каждое вызывается своим видом Corynebacterium), бактериальное увядание тыквенных, таких, как тыква и арбуз (вызываемое Erwiniatracheiphila), и черная гниль крестоцветных, например, капусты, вызываемая Xanthomonas campestris).
Представители рода Agrobacterium вызывают у растений образование корончатых галлов. Они имеют большое значение в генной инженерии и будут обсуждаться в гл. 30.
Микоплазмы
Микоплазмы — это группа мельчайших бактерий (около 0,1 мкм в диаметре). Плазматическая мембрана окружает ДНК, РНК, рибосомы, растворимые белки, углеводы и липиды. Микоплазмы лишены ядра, органелл и клеточной оболочки. Микоплазмы, вероятно, содержат немногим более 650 генов, что в 5 раз меньше, чем в среднем у бактерий.
Выделенные в особую группу, микоплазмы разделены на 6 родов, причем 50 или 60 описанных видов относятся к роду Mycoplasma. Изучая нуклеотидные последовательности рРНК, К. Р. Уоз и его коллеги определили, что все роды микоплазм, кроме одного, близки друг другу и, возможно, произошли от одной линии бактерий, включающей Bacillus и Lactobacillus. В то же время род Thermoplasta, очевидно, приобрел особенности микоплазм независимо. Все микоплазмы, вероятно, можно считать бактериями, упростившими свою структуру в процессе эволюции.
Помимо микоплазм, обнаруженных в животных тканях, найдены микоплазмоподобные организмы (MLO, от англ. mycoplasmalike organisms) у 200 видов растений, вызывающие около 50 заболеваний, как правило, характеризующихся пожелтением и низкорослостью. Многочисленные попытки культивировать MLO, выделенные из растений, были безуспешными.
Некоторые спироплазмы — длинные (~ 10 мкм) тонкие (диаметр 0,2 мкм) спиралевидные микоплазмы — подвижны в жидкой среде и культивируются in vitro в искусственных средах. К ним относится Spiroplasma citri, вызывающая мелколистность цитрусовых, и другие патогены. В культуре спироплазмы совершают интенсивные вихревые и волнистые движения. Доказательства патогенности спироплазм, выделенных из больных тканей, получены в отношении небольшого числа видов.
Рис. 11-24. Спироплазмы, выделенные из низкорослого растения кукурузы (Zea mays)
MLO обычно проникают в ситовидные трубки флоэмы. Полагают, что микроорганизмы переносятся с раствором сахаров из одной ситовидной трубки в другую через поры и далее транспортируются по флоэме. Некоторые спироплазмы, однако, способны активно передвигаться по тканям растений.
Цианобактерии
Цианобактерии («сине-зеленые водоросли») заслуживают специального обсуждения благодаря их многообразной экологической роли. Кроме того, цианобактерии часто рассматриваются как растения, а не как одна из групп бактерий. Хотя ранее к цианобактериям относили около 7500 видов, экспериментальные исследования дают основание считать, что только немногим более 200 различных несимбиотических видов действительно относятся к цианобактериям. Эти виды обладают большим разнообразием признаков в зависимости от среды обитания. Например, Microcoleus vaginatus живет во влажной почве, в пресной и соленой воде в различных районах земного шара — от северной Гренландии до Антарктики, от Долины Смерти до горы Пайкс-Пик. При изменении условий окружающей среды бактериальные клетки, составляющие даже одну колонию Microcoleus могут иметь чрезвычайно разнообразную морфологию.
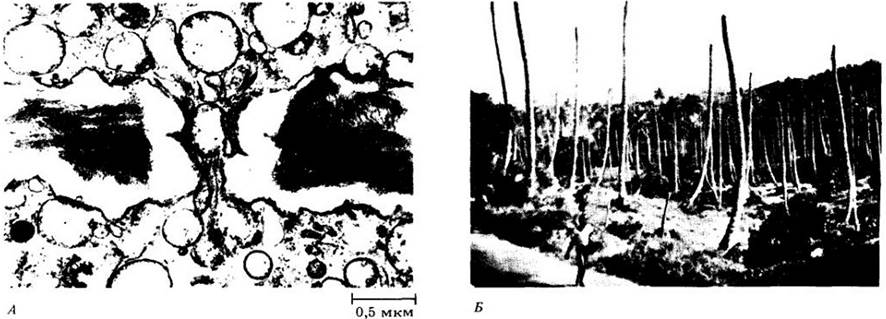
Рис. 11-25. А. Микоплазмоподобные организмы проникают через пору ситовидной пластинки молодого соцветия кокосовой пальмы (Cocos nucífera), вызывая летальное заболевание — желтуху. Б. Опустошенная роща кокосовых пальм на Ямайке, которые сейчас выглядят как телеграфные столбы. Летальная желтуха поражает многие виды пальм, произрастающих на юге Флориды или в других районах
Подобно другим бактериям, цианобактерии могут обитать в крайне неблагоприятных условиях — от горячих источников до замерзших озер Антарктики, где образуют слой толщиной 2— 4 см, лежащий в воде ниже уровня постоянного льда на 5 м. Зеленая окраска некоторых белых медведей в зоопарках обусловлена колониями цианобактерий, которые развиваются в полостях волос, образующих меховой покров.
Цианобактерии оказались первыми поселенцами на новом острове Суртсэй вблизи Исландии, образовавшемся в результате извержения вулкана. С другой стороны, они отсутствуют в кислых водах, где, напротив, эукариотические водоросли довольно многочисленны.
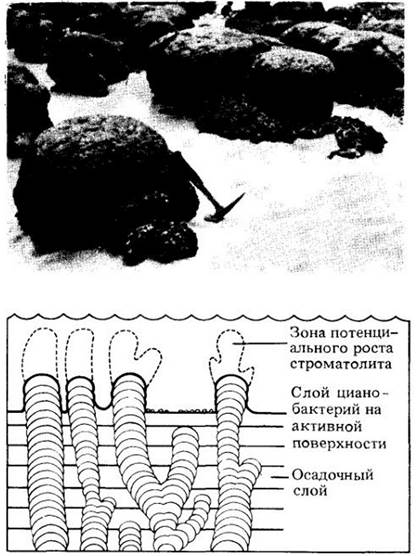
Слоистые меловые отложения, называемые строматолитами (рис. 11-26), имеют огромный геологический возраст — 2,7 млрд, лет — и возникли при захвате кальция цианобактериями. В настоящее время строматолиты образуются только в жарком сухом климате, в местах, где имеются мелкие водоемы. Обилие строматолитов в ископаемых остатках свидетельствует о существовании аналогичных климатических условий в отдаленные геологические эпохи, когда цианобактерии играли решающую роль в повышении уровня свободного кислорода в атмосфере ранней Земли.
Рис. 11-26. Образование строматолитов происходит, когда обширные колонии цианобактерий пропитываются и укрепляются карбонатом кальция. Строматолиты имеют самую разнообразную форму, например, куполообразную, как показано на рисунке и фотографии. Они часто встречаются в ископаемых породах, а в настоящее время образуются только в сугубо специфических условиях, например, на литорали залива Шарк в Западной Австралии
Многие цианобактерии имеют слизистую капсулу, или чехол, часто яркой окраски, особенно у наземных видов. Окраска может быть светло-золотой, желтой, коричневой, красной, изумрудной, голубой, фиолетовой и темно-синей. Кроме того, каротиноиды и фикобилины, присутствующие в клетках цианобактерий, модифицируют их окраску. Несмотря на свое название, только около половины «сине-зеленых водорослей» действительно имеют этот цвет. Красное море получило свое название, вероятно, благодаря «цветению» морских планктонных видов рода Trichodesmium (цианобактерий, содержащих вакуоли), которые часто встречаются на поверхности этого моря.
Многие морские цианобактерии обитают в известняках или на субстратах, богатых известью, таких, как раковины моллюсков или кораллиновые водоросли. Пресноводные виды, особенно живущие в горячих источниках, часто образуют толстые слои известняка в своих колониях. В Йеллоустонском национальном парке нитчатая форма Mastigocladus обитает в горячей воде при 55° С, а одноклеточная Synechococcus выдерживает температуры до 73 — 75° С. Цианобактерии широко распространены в почве, включая и пустыни; насчитывается 20 000 — 50 000 таких организмов на 1 г почвы.
Представители многих родов цианобактерий фиксируют азот. В теплых районах Азии рис может довольно долго расти на одном и том же участке без применения удобрений благодаря азотфиксирующим цианобактериям. Цианобактерии, особенно из рода Anabaena, обитают в полостях листьев водного папоротника Azolla, который в свою очередь имеет почти облигатные связи с этими цианобактериями. Благодаря своей способности фиксировать молекулярный азот цианобактерии могут заселять голые поверхности скал и почву вместе с лишайниками, печеночниками и другими организмами, с которыми вступают в симбиоз. Морские виды Trichodesmium фиксируют около четверти всего поглощаемого морем азота, т. е. довольно значительное количество.
Фиксация азота происходит в гетероцистах — клетках большого размера, которые образуются у нитчатых форм (см. рис. 11-13, 11-15). Оболочка гетероцисты аналогична клеточной оболочке цианобактерий, однако снаружи имеет два дополнительных слоя. Внешний слой состоит из полисахаридов, а внутренний — из гликолипидов. Внутренние мембраны гетероцисты образуют концентрическую и сетчатую структуру. Поскольку в гетероцистах нет фотосистемы II (гл. 7) и происходит только циклическое фосфорилирование, кислород не выделяется, а имеющийся сразу же восстанавливается водородом — побочным продуктом фиксации азота — или удаляется через клеточную стенку гетероцисты. Нитрогеназа чувствительна к кислороду, и фиксация азота — анаэробный процесс. Образование гетероцист у Nostoc и бактерий других родов ингибируется аммиаком и нитратами, однако, когда содержание этих азотистых веществ падает ниже определенного уровня, гетероцисты начинают появляться вновь. Как уже упоминалось, некоторые цианобактерии образуют устойчивые споры — акинеты, которые отличаются по структуре и функции от гетероцист.
Цианобактерии могут вступать в симбиотические отношения с некоторыми губками, амебами, жгутиковыми простейшими, диатомовыми и зелеными водорослями, лишенными хлорофилла, другими цианобактериями, мхами, сосудистыми растениями и оомицетами. При этом они теряют клеточную оболочку и функционируют как хлоропласты. Симбиотическая цианобактерия делится одновременно с клеткой-хозяином аналогично хлоропластам. Эволюционные связи цианобактерий с некоторыми эукариотами будут обсуждены в гл. 15.