СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 18. СЕМЕННЫЕ РАСТЕНИЯ
Голосеменные
Существуют четыре отдела голосеменных с современными представителями: Cycadophyta (саговниковые), Ginkgophyta (гинкговые), Coniferophyta (хвойные) и Gnetophyta(гнетовые). Название всей группы указывает на одну из важнейших характеристик всех семенных, за исключением цветковых: семязачатки и семена «голые», т. е. расположены открыто на поверхности спорофиллов или аналогичных структур. Четыре отдела, возможно, отражают определенные стадии эволюционного развития, достигнутые различными потомками прогимноспермов.
За несколькими исключениями, каждый женский гаметофит голосеменных образует несколько архегониев. В результате может быть оплодотворено несколько яйцеклеток, и в одном семязачатке иногда начинают развиваться несколько зародышей. Это явление называется полиэмбрионией. В большинстве случаев выживает только один зародыш; лишь немногие полностью развитые семена отклоняются от этого правила.
Чтобы подвижные, снабженные жгутиками спермин споровых сосудистых растений могли достичь яйцеклеток и оплодотворить их, необходима вода. У голосеменных она для этого не нужна. Их не до конца развитый мужской гаметофит, пыльцевое зерно, переносится целиком (обычно пассивно, с помощью ветра) к женскому гаметофиту, находящемуся внутри семязачатка. Этот процесс называется опылением. Затем мужской гаметофит образует вырост, пыльцевую трубку. У хвойных и гнетовых сами спермин неподвижны, и эти трубки доставляют их прямо к архегониям. У саговниковых и у гинкговых спермин многожгутиковые, и их пыльцевые трубки, по-видимому, функционируют как гаустории (аналогичные гаусториям паразитических грибов), проникающие в семязачатки и поглощающие из них питательные вещества. Пыльцевая трубка может несколько месяцев расти в ткани нуцеллуса, или мегаспорангия, пока достигнет полости над женским гаметофитом, после чего лопается, выпуская в нее два спермия. Спермин плывут к архегонию, и один из них оплодотворяет яйцеклетку. С развитием переносящей спермин пыльцевой трубки размножение сосудистых растений перестало зависеть от присутствия жидкой воды, необходимой для оплодотворения всем споровым.
Хвойные
Эта группа, намного превосходящая по численности и распространению другие отделы современных голосеменных, включает около 50 родов с примерно 550 видами, в том числе самое высокое сосудистое растение — секвойю вечнозеленую (Sequoia sempervirens), растущую на побережье Калифорнии и юго-западе Орегона (см. рис. 31-1), которая достигает 117 м в высоту при диаметре ствола более 11 м. Хвойные, к которым принадлежат также сосны, пихты и ели, имеют большое хозяйственное значение; их высокоствольные леса — одно из основных богатств обширных областей севера умеренной зоны. В начале третичного периода некоторые роды этого отдела были распространены шире, чем сейчас, и господствовали на огромных пространствах всех континентов Северного полушария.
История хвойных начинается по крайней мере в позднем карбоне, около 290 млн. лет назад; их примитивными представителями были жившие в то время кордаитовые (см. приложение к гл. 17). Листья современных хвойных имеют много черт, свидетельствующих об их засухоустойчивости, что может быть связано с формированием разнообразия этого отдела на протяжении относительно сухого и холодного пермского периода (286 — 248 млн. лет назад). В то время повсеместное возрастание аридности, вероятно, благоприятствовало такого рода структурным адаптациям.
Сосны
Сосны (род Pinus) — наверное, самые известные из голосеменных (рис. 18-9); они доминируют на огромных площадях Северной Америки и Евразии и широко культивируются даже в Южном полушарии. Около 90 их видов характеризуются уникальным средисовременных голосеменных листорасположением. Листья сосен игловидные. У проростков они располагаются спирально и появляются на стеблях по одному (рис. 18-10). Через год или два у сосны начинают появляться пучки длинных игл (хвоинок), каждый из которых содержит строго определенное их число (от одного до восьми в зависимости от вида). Эти пучки, одетые у основания рядами коротких чешуевидных листьев, представляют собой укороченные побеги, у которых подавлена активность апикальной меристемы (рис. 18-11). Таким образом, пучок хвоинок у сосны в морфологическом отношении является детерминированной(ограниченной в росте) веткой. При необычных условиях его апикальная меристема может возобновить свою активность и дать новый побег с неограниченным (недетерминированным) ростом, способный иногда образовать корни, вырастая в целое дерево (рис. 18-12).
Рис. 18-10. А. Проростки сосны болотной (шт. Джорджия) с ювенильными листьями (длинные одиночные хвоинки) и первыми взрослыми листьями, собранными у этого вида в пучки по три. Б. Проросток сосны съедобной (Pinus edulis) с ювенильньми листьями и молодым главным корнем. Взрослые листья этого вида собраны в пучки по два

Рис. 18-11. А. Сосна остистая (Рinus longaeva) в Брайс-Каньоне, шт. Юта. На ветке видны пучки из пяти хвоинок и зрелая женская шишка. Отдельные иглы могут функционировать до 45 лет; эта сосна — самое долгоживущее дерево (см. также рис. 23-27, А). Б. Ветка сосны смолистой (Pinus resinosa) с молодыми женскими шишками и пучками игловидных листьев, характерных для взрослого растения

Рис. 18-12. Однолетние экземпляры сосны замечательной (Рinus radiata), выросшие из укорененных пучков хвои, являющихся, следовательно, укороченными побегами с подавленной активностью апикальной меристемы, которая может быть восстановлена

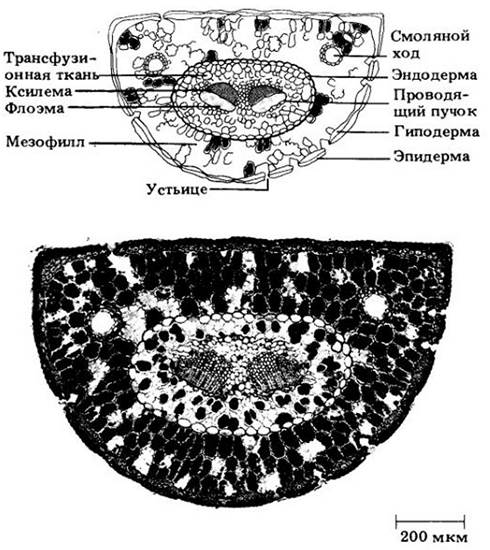
Листья сосен, как и у многих других хвойных, прекрасно приспособлены к произрастанию в аридных условиях (рис. 18-13). Их эпидерма покрыта толстой кутикулой, под которой компактно располагаются один или более слоев толстостенных клеток — гиподерма. Устьица погружены в углубления на поверхности листьев. Мезофилл, или основная ткань листа, состоит из паренхимных клеток с извилистыми стенками, причем выступы на поверхности одной клетки заходят в углубления другой. Обычно мезофилл пронизан двумя или более смоляными ходами. В центре листа одни или два расположенных рядом проводящих пучка окружены трансфузионной тканью, состоящей из живых паренхимных клеток и коротких мертвых трахеид. Полагают, что эта ткань обеспечивает обмен веществами между мезофиллом и пучками. Она окружена эндодермой, так что не находится в прямом контакте с мезофиллом.
Рис. 18-13. Поперечный разрез сосновой хвоинки, иллюстрирующий зрелые ткани
Большинство сосен не меняет свою хвою 2 — 4 года, и общий фотосинтетический баланс растения зависит от состояния всех образовавшихся за несколько лет хвоинок. У сосны остистой (Pinus longaeva), самого долговечного дерева (см. рис. 18-11, А и 23-27, А), они сохраняются до 45 лет, оставаясь при этом фотосинтетически активными. Поскольку у сосны и других вечнозеленых растений листья функционируют более одного вегетационного периода, они испытывают действие засухи, холодов или загрязнения воздуха гораздо дольше, чем у листопадных растений, и чаще оказываются поврежденными.
В стеблях сосен и других хвойных рано начинается вторичный рост, ведущий к образованию значительного количества вторичной ксилемы (рис. 18-14). Она откладывается внутрь от камбия, а вторичная флоэма снаружи от него. Ксилема хвойных состоит главным образом из трахеид, а флоэма — из ситовидных клеток, типичных, проводящих питательные вещества элементов голосеменных и споровых сосудистых растений (см. гл. 20). Оба типа тканей пересечены по радиусам узкими лучами. В начале вторичного роста эпидерма замещается перидермой, закладывающейся во внешнем слое коры. По мере этого роста следующие слои перидермы возникают за счет активного деления клеток, расположенных глубже в коре.
Рис. 18-14. Поперечный разрез стебля сосны, показывающий вторичную ксилему и вторичную флоэму, разделенные камбием. Все ткани снаружи от камбия, включая флоэму, составляют кору

Размножение сосен. Микро- и мегаспорангии сосен и большинства других хвойных образуются в отдельных шишках на одном и том же дереве. Обычно микроспоронгиатные (мужские) шишки формируются на нижних ветвях дерева, а мегаспорангиатные (женские) — на верхних; у некоторых видов они располагаются на одной и той же ветви, но при этом женские шишки ближе к ее концам. Так как разносимая ветром пыльца обычно не летит прямо вверх, семязачатки, как правило, опыляются пыльцой с другого дерева, за счет чего происходит перекрестное опыление.
Мужские шишки у сосен относительно мелкие, обычно 1 — 2 см длиной (рис. 18-15). Более или менее пленчатые микроспорофиллы (рис. 18-16) располагаются спирально; каждый из них несет два микроспорангия. В молодом микроспорангии множество микроспороцитов, или материнских клеток микроспор; ранней весной они делятся мейотически, образуя четыре гаплоидные микроспоры каждый. Каждая микроспора развивается в пыльцевое зерно с воздушными мешками, состоящее из двух проталлиальных клеток, генеративной клеткии клетки трубки (рис. 18-17). Эта четырехклеточная структура и есть незрелый мужской гаметофит. Именно на этой стадии происходит рассеивание огромных количеств пыльцевых зерен, некоторые из которых переносятся ветром к женским шишкам.
Рис. 18-15. Сосна замечательная. Микроспорангиатные (мужские) шишки, рассеивающие пыльцу, которая уносится ветром. Часть пыльцы оказывается поблизости от семязачатков в женских шишках и прорастает, образуя пыльцевые трубки. В результате происходит оплодотворение

Рис. 18-16. Продольный разрез мужской шишки сосны; видны микроспорофиллы и микроспорангии, содержащие зрелые пыльцевые зерна

Рис. 18-17. Сосна. А. Пыльцевые зерна с незрелыми мужскими гаметофитами внутри. Каждый гаме то фит состоит из двух проталлиальных клеток, относительно мелкой генеративной клетки и относительно крупной клетки трубки. Б. Пыльцевое зерно на несколько более поздней стадии; проталлиальные клетки, не имеющие никакой явной функции, уже дегенерировали. В. Пыльцевое зерно с двумя воздушными мешками под сканирующим электронным микроскопом. Когда оно прорастает, пыльцевая трубка выходит снизу между воздушными мешками

Последние у сосен намного крупнее и устроены сложнее, чем мужские (рис. 18-18). Их чешуи не являются мегаспорофиллами, а представляют собой целые видоизмененные укороченные побеги, включающие кроме семенной чешуи с двумя семязачатками на верхней поверхности кроющую стерильную чешую (рис. 18-19). Они располагаются спирально вокруг оси шишки. Таким образом, женская шишка — сложная структура, а мужская — простая, поскольку в ней микроспорангии непосредственно прикреплены к микроспорофиллам. Каждый семязачаток содержит многоклеточный нуцеллус (мегаспорангий), окруженный массивным интегументом с отверстием (микропиле), обращенным к оси шишки (рис. 18-19). В мегаспорангии находится единственный мегаспороцит, т. е. материнская клетка мегаспоры, дающая в результате мейоза линейный ряд из четырех мегаспор. Функциональна только одна из них, а три другие, расположенные ближе к микроииле, вскоре отмирают.
Рис. 18-18. Относительные размеры некоторых зрелых шишек сосен. А. Сосна Сабина (Pinus sabiniana). Б. Сосна съедобная (Pinus edulis), вид сверху и сбоку; съедобные семена этой и некоторых других сосен называют «сосновыми орешками», они бескрылые и разносятся главным образом кедровками, птицами из семейства вороновых. В. Сосна Ламберта (Pinus lambertiana). Г. Сосна желтая (Pinus ponderosa). Д. Сосна веймутова (Pinus strobús). Е. Сосна смолистая (Pinus resinosa)

Рис. 18-9. Сосна болотная (Pinus palustris) в Северной Каролине

Опыление у сосен происходит весной; пыльца прилипает к капле клейкой жидкости, выделяющейся в области микропиле. На этой стадии чешуи женской шишки широко раздвинуты. При испарении микропилярной жидкости пыльцевое зерно втягивается в микропиле и попадает на нуцеллус. После опыления чешуи сближаются, что способствует защите развивающихся семязачатков. Вскоре после соприкосновения пыльцевого зерна с нуцеллусом оно прорастает, образуя пыльцевую трубку. В это время мейоз в мегаспорангии еще не произошел. Примерно через месяц после опыления появляются четыре мегаспоры, одна из которых дает мегагаметофит. Его развитие замедленное, часто начинается только через шесть месяцев после опыления и может потребовать еще полгода для своего завершения. На ранних стадиях формирования мегагаметофита митоз не сопровождается немедленным образованием клеточных стенок, которое начинается примерно через 13 месяцев после опыления, когда уже имеется около 2000 свободных ядер. Затем (примерно через 15 месяцев после опыления) у микропилярного конца мегагаметофита дифференцируются архегонии (два-три); теперь условия для оплодотворения созданы.
Вис. 18-19. Сосна. А. Продольный разрез молодой женской шишки, демонстрирующий ее сложную структуру. Б. Увеличенный фрагмент разреза. Виден мега- спороцит (материнская клетка мегаспоры), окруженный нуцеллусом

Приблизительно за 12 месяцев до этого пыльцевое зерно проросло в пыльцевую трубку, которая медленно прокладывала себе путь через ткани нуцеллуса к развивающемуся мегагаметофиту. Через год после опыления генеративная клетка четырехклеточного мужского гаметофита разделилась на две дочерние — стерильную клетку (клетку ножки) и сперматогенную клетку (клетку тела). Затем, прежде чем пыльцевая трубка достигает женского гаметофита, сперматогенная клетка делится, образуя два спермия. Мужской гаметофит, или прорастающее пыльцевое зерно, теперь достиг зрелости. У семенных растений антеридиев нет.
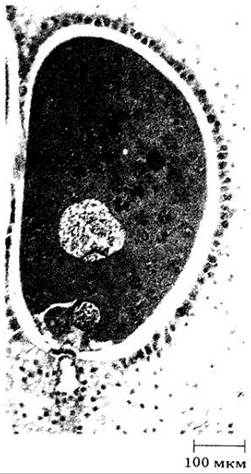
Приблизительно через 15 месяцев после опыления пыльцевая трубка достигает яйцеклетки архегония и впрыскивает в нее большое количество своей цитоплазмы и оба спермия (рис. 18-20). У одного из спермиев ядро соединяется с ядром яйцеклетки, а у второго дегенерирует. Обычно оплодотворяются и начинают развиваться в зародыши яйцеклетки всех архегониев (полиэмбриония), однако, как правило, только один зародыш формируется полностью.
Рис. 18-20. Оплодотворение у сосны: слияние ядер спермин и яйцеклетки. Второе ядро спермия (внизу) не функционально и в конце концов отмирает
На ранней стадии эмбриогенеза вблизи нижнего конца архегония образуются четыре яруса клеток. Каждая из четырех клеток самого верхнего (т. е. наиболее удаленного от микропилярного конца семязачатка) яруса начинает развиваться в зародыш. Одновременно четыре клетки следующего ряда, клетки суспензора (подвеска), сильно вытягиваются и проталкивают четыре формирующихся зародыша вглубь женского гаметофита. Таким образом, в жизненном цикле сосны имеется и второй тип полиэмбрионии, но опять же только один из зародышей развивается полностью. В ходе эмбриогенеза интегумент превращается в семенную оболочку.
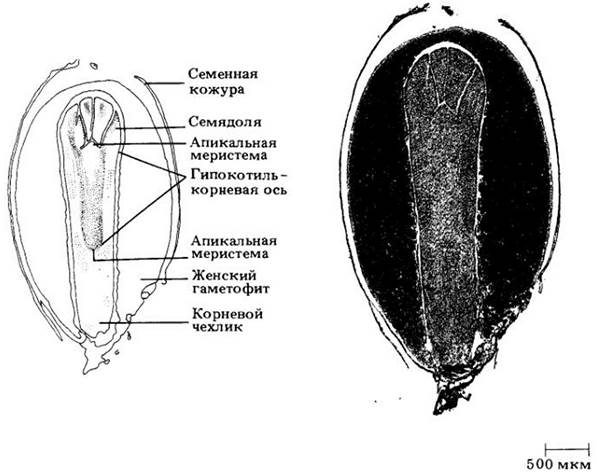
Можно видеть, что семя хвойных состоит из комбинации двух спорофитных поколений — семенной кожуры (и остатков нуцеллуса) и зародыша — и одного гаметофитного, служащего запасом питательных веществ (рис. 18-21). Семенная кожура и зародыш диплоидны, а женский гаметофит гаплоидный. Зародыш представляет собой гипокотиль-корневую ось с чехликом и апикальной меристемой на одном конце и апикальной меристемой и несколькими (обычно восемью) семядолями на другом. Интегумент состоит из трех слоев, из которых средний становится твердым и служит семенной кожурой.
Рис. 18-21. Продольный разрез соснового семени
Семена сосен часто высыпаются осенью второго года после появления шишек и опыления. Чешуи зрелых шишек раздвигаются; крылатые семена большинства видов планируют в воздухе и иногда переносятся ветром на значительные расстояния. Иногда, например, у сосны Банкса (Pinus banksiana), чешуи не раздвигаются до тех пор, пока шишки сильно не прогреются. Когда по сосняку быстро проносится лесной пожар, сжигающий родительские деревья, большинство огнестойких шишек только слегка опаляется, после чего, раскрываясь, высвобождает накопившийся за многие годы урожай семян. За счет этого лес на недавно обгоревшей почве возобновляется. У других видов, включая сосны гибкую (Pinus flexilis) и белоствольную (Pinus albicaulis), а также однохвойные сосны запада Северной Америки и некоторые сходные с ними виды Евразии, крупные бескрылые семена собираются, переносятся и запасаются впрок как корм кедровками, крупными птицами из семейства вороновых.
Жизненный цикл сосны представлен на рис. 18-25.
Другие хвойные
Хотя у других хвойных (см. рис. 18-22 — 18-29) нет характерных для сосен пучков хвои и существует также ряд относительно мелких отличий в строении репродуктивных систем, современные представители этого отдела образуют достаточно однородную группу. Важнейшие их роды — пихта (Abies; рис. 18-22), ель (Picea), тсуга (Tsuga), лжетсуга (Pseudotsuga),кипарис (Cupressus; рис. 18-23) и можжевельник (Juniperus; рис. 18-24). У тиссов (семейство Тахасеае) семязачатки формируются не в шишках, а по одиночке и окружены мясистым чашевидным образованием — ариллусом (рис. 18- 26, А). Ряд других интересных родов хвойных встречается главным образом в Южном полушарии. Некоторые из них. подобно араукарии разнолистной (Araucaria heterophylla) с острова Норфолк и араукарии чилийской (А. агаuсаnа), часто разводятся в областях с достаточно мягким климатом, а другие виды пересекают экватор и естественным путем.
Рис. 18-22. Два рода из семейства сосновых (Pinaceae). А. Женские шишки пихты бальзамической (Abies balsamea): они прямостоячие, имеют длину 5 — 10 см и не опадают на землю целиком, как у сосен, а рассыпаются на части, рассеивая крылатые семена. Б. Лиственница европейская (Larix decidua). Ее игловидные, как у сосны, листья появляются по одному и располагаются спирально на укороченных побегах. В отличие от большинства хвойных лиственницы сбрасывают всю хвою в конце каждого вегетационного периода

Рис. 18-23. Кипарис калифорнийский (Cupressus goveniana). Эти невысокие (спелые — всего около 6 м высотой) деревья встречаются только вблизи Монтерея в Калифорнии
Рис. 18-24. У можжевельника обыкновенного (Juniperus communis) женские шишки шаровидные, как у кипарисов, но их мясистые чешуи срастаются между собой в «ягоды», которые используют для придания специфического вкуса и запаха джину

Одно из наиболее интересных семейств хвойных — таксодиевые (Taxodiaceae), появившиеся около 150 млн. лет назад, а сейчас представленные изолированными территориально видами, остатками популяций, гораздо шире распространенных в третичном периоде. Один из наиболее замечательных среди них — секвойя вечнозеленая (Sequoia sempervirens), самое высокое из ныне живущих растений (см. рис. 31-1). К этому же семейству принадлежат знаменитое «мамонтово дерево», Sequoiadendron giganteum, образующее эффектные, изолированные друг от друга рощи по западному склону хребта Сьерра-Невада в Калифорнии, а также болотный кипарис (Taxodium) с юго-востока США и из Мексики (рис. 18-27). Все эти роды были гораздо шире распространены в третичном периоде, чем сейчас (рис. 18-29).
Обильно в третичном периоде в Евразии и Северной Америке произрастала и метасеквойя (Metasequoia) (рис. 18-28), наиболее частое хвойное в западной и арктической Северной Америке с позднего мела до миоцена (другими словами, примерно 90 — 15 млн. лет назад). В Японии и Восточной Сибири она встречалась всего несколько миллионов лет назад. Этот род описан по ископаемому материалу японским палеоботаником Сигеру Мики в 1941 г. (рис. 18-29). Тремя годами позже китайский лесовод Цан Ван из Центрального бюро лесных исследований Китая посетил деревню Мотаочи в провинции Сычуань на юго-западе страны, где увидел огромное дерево, прежде ему неизвестное. Местные жители выстроили вокруг его основания целый храм. Изучение собранных Цан Ваном образцов хвои и шишек показало, что это «ископаемая» метасеквойя. В 1948 г. палеоботаник Р. Чейни из Калифорнийского университета возглавил экспедицию по реке Янцзы и через три горных хребта вывел ее к долинам, где росло множество этих деревьев, последних остатков когда-то обширных лесов из метасеквойи. Впоследствии были собраны тысячи ее семян, и это «живое ископаемое» сейчас можно видеть в парках и садах всего мира.
Другие современные голосеменные
Саговниковые
Другие группы современных голосеменных мало похожи друг на друга вообще. Среди них саговниковые, или цикадофиты (отдел Сусаdophyta), — внешне напоминающие пальмы растения главным образом тропических и субтропических областей. Эти причудливые формы, появившиеся не позднее 285 млн. лет назад, где-то в начале пермского периода, были столь многочисленны в мезозое, что его нередко называют «временем саговников и динозавров». Цикадофиты родственны семенным папоротникам (см. приложение к гл. 17), и среди ископаемых остатков встречаются, по-видимому, переходные между ними формы. Ныне сохранилось 10 родов этого отдела, включающих около 100 видов. Замия карликовая (Zamia pumila), обычная в песчаных редколесьях Флориды, — единственный местный представитель саговников в США (рис. 18-30).
Большинство цикадофитов довольно крупные растения; некоторые из них достигают в высоту более 18 м. У многих четко выражен ствол, густо покрытый основаниями опавших листьев. Функционирующие листья собраны пучком на вершине стебля — этим саговниковые напоминают пальмы (недаром некоторые из них обычно называют «саговые пальмы»), однако в отличие от последних наблюдается настоящий, хотя и медленный, вторичный рост в результате деятельности камбия. Центральная часть их стволов содержит большое количество сердцевины. Репродуктивные органы саговниковых состоят из более или менее редуцированных листьев с прикрепленными к ним спорангиями, рыхло или плотно скученных в шишкоподобных образованиях у верхушки растения. Мужские и женские «шишки» располагаются на разных особях (рис. 18-31).
Гинкго
Гинкго двулопастный (Ginkgo biloba) легко узнается по вееровидным листьям с дихотомически расходящимися жилками (рис. 18-32). Это красивое и крупное, но медленно растущее дерево иногда достигает в высоту более 30 м. Листья на многочисленных укороченных побегах более или менее цельные, а на удлиненных и у проростков — глубоколопастные. В отличие от большинства других голосеменных гинкго листопадный; его листья приобретают осенью перед сбрасыванием красивый золотистый оттенок.
Это единственный доживший до наших дней вид мало изменившегося за 80 с лишним миллионов лет рода и единственный ныне живущий представитель отдела Ginkgophyta. Он имеет общие черты с другими родами голосеменных, жившими в начале пермского периода, около 280 млн. лет назад. Возможно, дикорастущих гинкго уже не осталось нигде в мире, а дошедшие до нас экземпляры сохранены около храмов в Китае и Японии. Завезенные оттуда, они широко разводятся в садах умеренных областей мира уже более 150 лет. Эти деревья очень устойчивы к загрязнению воздуха и поэтому часто выращиваются в городских парках.
Как и у саговников, семязачатки и микроспорангии у гинкго образуются на разных деревьях. Семязачатки появляются парами на конце коротких ножек и осенью, созревая, дают семена с мясистым покровом. Оплодотворения, вероятно, не происходит до тех пор, пока они не опадут с родительских деревьев. Зародыши формируются на поздних стадиях созревания лежащих на земле семян. Последние имеют неприятный запах из-за присутствия в их мясистых покровах масляной кислоты, поэтому в парках и садах обычно разводят только мужские деревья, которые размножаются вегетативно. Микроспорофиллы собраны в сережковидные структуры; каждый из них несет два микроспорангия.
Гнетовые
В этот отдел входят около 70 видов из трех современных родов очень необычных голосеменных: гнетум (Gnetum), эфедра (Ephedra) и вельвичия (Welwitschia). Хотя они, несомненно, родственны между собой и справедливо объединяются вместе, их признаки сильно различаются. Интересно присутствие черт, сближающих их с покрытосеменными; это и сходство их стробилов с некоторыми соцветиями последних, и присутствие почти таких же сосудов в ксилеме, и отсутствие архегониев у гнетума и вельвичии. Десятки лет ученые спорили, родственны ли гнетовые цветковым. В настоящее время появились доказательства в пользу такого родства, хотя, конечно, ни одно из ныне живущих гнетовых нельзя считать предком какого-либо покрытосеменного; каждый их трех современных родов отдела своеобразно специализирован.
Примерно 30 видов гнетума — деревья и лианы с крупными кожистыми листьями, сильно напоминающими листья двудольных (рис. 18-33). Они встречаются во всех влажных тропических областях.
Большинство из приблизительно 35 видов эфедры — сильно разветвленные кустарники со слабозаметными мелкими чешуевидными листьями (рис. 18-34). Растение с внешне членистыми стеблями на первый взгляд напоминает хвощ. Большинство видов рода обитает в аридных или пустынных областях.
Вельвичия, наверное, самое причудливое сосудистое растение (рис. 18-35). Большая его часть погружена в песчаную почву, а над землей выступает массивный деревянистый вогнутый диск, от которого отходят только два ремневидных листа и ветви со стробилами, образуемые меристематической тканью по краю диска. Вельвичия растет в пустынях юго-западной Африки — в Анголе, Намибии и ЮАР.