МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 7. ЦВЕТОК КАК ОСОБЫЙ РЕПРОДУКТИВНЫЙ ОРГАН ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
7.6. Развитие тычинки в онтогенезе. Характеристика процессов, происходящих в пыльнике
7.6.1. Формирование тычинки, развитие и строение пыльника
Как было отмечено ранее, тычинки закладываются на цветоложе после заложения элементов околоцветника. У большинства покрытосеменных растений развитие тычинок в цветке, если они расположены в несколько кругов или по спирали, идет от периферии к центру, но, у крестоцветных, тычинки формируются в центробежном направлении — от центра к периферии.
Первоначально бугорок меристемы заложившегося пыльника разрастается равномерно, клетки поверхностного слоя меристемы делятся антиклинально (перпендикулярно поверхности) и дают начало эпидермису. Эпидермальные клетки имеют типичное для этой ткани строение. Наружная стенка клеток эпидермиса сильно утолщена, на поверхности ее образуется слой кутикулы, а иногда и слой воска. Особенно сильно утолщается оболочка клеток эпидермиса в местах вскрывания пыльника. Все остальные клетки длительное время сохраняют меристематическую функцию, что приводит к увеличению меристематического бугорка, а затем и к его дифференцировке. В процессе дифференцировки бугорок приобретает четырехлопастную форму. В каждой лопасти впоследствии сформируется по одному микроспорангию (пыльцевой камере) (рис. 165).
Рис. 165. Заложение и развитие пыльника тычинки: А—меристематический бугорок с выделившейся субэпидермальной клеткой; Б—появление первичного археспория; В —образование вторичного археспория и париетальных клеток; Г—начало образования микроспорангиев со спорогенными клетками и дифференциация стенки пыльника; Д—сформированные микроспорангии со спорогенной тканью: 1—эпидермис; 2—субэпидермальная клетка; 3—первичный археспорий; 4—вторичный археспорий; 5— париетальные клетки; 6—образующаяся спорогенная ткань; 7—клетки, формирующие стенку пыльника; 8—сосудисто - волокнистый пучок; 9—микроспорангий со спорогенной тканью; 10—стенка молодого пыльника
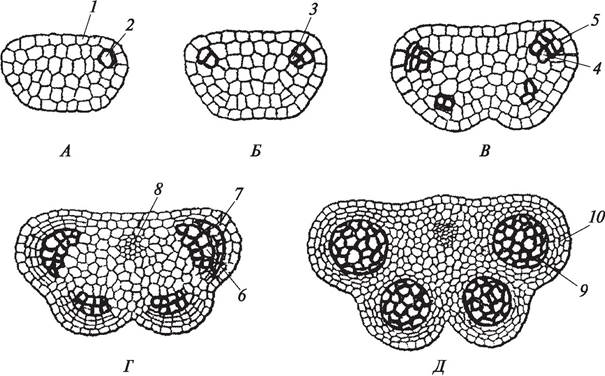
На ранних этапах развития в каждой из лопастей пыльника под эпидер мисом начинает выделяться по одной клетке, которые представляют собой клетки первичного археспория. От остальных меристематических клеток они отличаются более крупными размерами, большим ядром и, несомненно, био химическими особенностями. В частности, в клетках первичного археспория выявлено высокое содержание РНК, основных и кислых белков, некоторых ферментов (пероксидазы, фосфатазы), однако в биохимическом плане археспорий изучен очень слабо. Это в полной мере относится к большинству эмбриологических структур.
Первичные археспориальные клетки путем митоза делятся тангентально (периклинально — параллельно поверхности органа) и дают начало двум типам клеток: париетальным (постенным) клеткам и клеткам вторичного археспория. Из париетальных клеток в результате серии митотических делений и последую - щей дифференцировки образуется стенка пыльника. У разных видов покрыто - семенных растений она имеет разную толщину и характеризуется определенной специфичностью, но общий план ее строения в принципе однотипен (рис. 166).
Рис. 166. Фрагмент формирующегося пыльника. Дифференцировка стенки пыльника и микроспорангия (пыльцевой камеры): 1—эпидермис; 2—эндотеций (фиброзный слой); 3—клетки среднего слоя; 4—тапетум (выстилающий слой); 5—микроспорангий со спорогенными клетками (микроспороцитами)
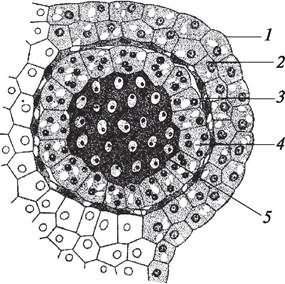
Под эпидермисом образуется эндотеций, или фиброзный слой. У сухопутных растений этот слой имеет своеобразные (фиброзные) утолщения на внутренней стороне клеточной оболочки и принимает активное участие в процессе вскрывания пыльника. При созревании пыльника, которое сопровождается усиленной потерей воды во всех клетках и особенно в поверхностных, оболочки клеток фиброзного слоя в более тонких местах начинают спадаться, что и приводит к активному вскрыванию пыльника в результате разрыва его стенки. У водных растений в клетках эндотеция фиброзных утолщений не образуется и вскрывание пыльника происходит пассивно путем разрушения клеток.
За эндотецием расположен средний слой, характеризующий его положение в стенке пыльника. Термин введен в 1898 г. Дж. М. Коултером. Клетки среднего слоя мелкие, их тонкие оболочки имеют первичное строение. Средний слой может иметь различную толщину. Это кратковременно существующий слой пыльника. По имеющимся данным, для него характерно наличие крахмала, каротина, липидных глобул, что свидетельствует о том, что этот слой выполняет трофическую функцию. Полагают, что клетки среднего слоя рано разрушаются и содержимое их идет на питание развивающихся микроспороцитов и микроспор.
За средним слоем формируется тапетум, или выстилающий слой. Он ограничивает (выстилает) микроспорангий и непосредственно примыкает к микроспороцитам. Тапетум — это полифункциональная ткань, которая обеспечивает нормальное протекание мейоза при образовании микроспор, участвует в раз - витии мужского гаметофита (пыльцы) и формировании оболочки пыльцевых зерен. Выполнению различных функций способствует как морфологическое строение клеток тапетума, так и его биохимические особенности. Клетки тапетума крупные, с большими ядрами, имеющими несколько ядрышек. Довольно часто наряду с клетками, содержащими одно ядро, обнаруживаются многоядерные клетки тапетума. Отмечено также, что некоторые клетки тапе тума имеют полиплоидные ядра. В клетках тапетума обнаружены нуклеино вые кислоты, большое количество ферментов, гетероауксин, аскорбиновая кислота и другие компоненты. Однако биохимические исследования тапетума отрывочны и выполнены на разных культурах. Несмотря на это, эмбриологи считают, что наличие вышеуказанных особенностей позволяет судить о высокой физиологической активности клеток тапетума. Об этом свидетельствуют и данные электронно микроскопических исследований. В клетках тапетума обнаружено большое количество рибосом, митохондрий, сформирован хорошо развитый эндоплазматический ретикулум. На внутренней стороне оболочек клеток тапетума выявлены особые образования — «тельца Убиша», или орбикулы. В орбикулах содержится спорополленин — особое вещество, которое входит в состав наружной оболочки пыльцы. Различают два типа тапетума — секреторный, или железистый, и амебоидный. Клетки железистого тапетума длительное время не разрушаются и функционируют как секреторная ткань. У амебоидного тапетума оболочки клеток к началу формирования мужского гаметофита разрушаются, содержимое клеток сливается в общую массу.
Имеет ли какие-нибудь преимущества один тип тапетума перед другим, каково их эволюционное соотношение (какой тип первичен) и таксономическое значение, до сих пор не ясно и остается предметом спора и научных дискуссий.
Одновременно и параллельно с формированием стенки пыльника из клеток первичного археспория в результате митотического деления образуются клетки вторичного археспория. Эти клетки или сами становятся материнскими клетками микроспор или претерпевают ряд митотических делений и дают начало материнским клеткам микроспор, или микроспороцитам. Многочисленные микроспороциты заполняют микроспорангии, расположенные по два в каждой половинке пыльника (см. рис. 166).
На определенном этапе развития в микроспорангиях начинают осуществляться репродуктивные процессы: микроспорогенез, образование мужского гаметофита, формирование мужских половых клеток (гамет) — спермиев.
7.6.2. Строение микроспороцитов. Микроспорогенез
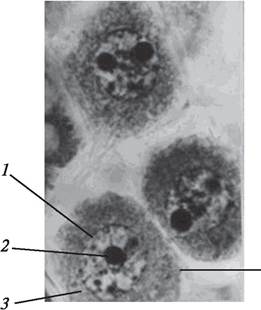
Микроспороциты являются диплоидными клетками, имеющими каллозную оболочку, невакуолизированную цитоплазму и крупные ядра. Молодые микроспороциты плотно прилегают друг к другу, но по мере развития и увеличения размеров клетки микроспороцита они расходятся и свободно располагаются в полости микроспорангия (рис. 167). Обособление микроспороцитов приводит к их разобщению и свидетельствует о подготовке к процессу микроспорогенеза — процессу образования микроспор.
Рис. 167. Микроспороциты бобов обыкновенных (Viciafaba) в микроспорангиях пыльника перед началом редукционного деления: 1—ядро; 2—ядрышко; 3—цитоплазма; 4 —оболочка микроспороцита
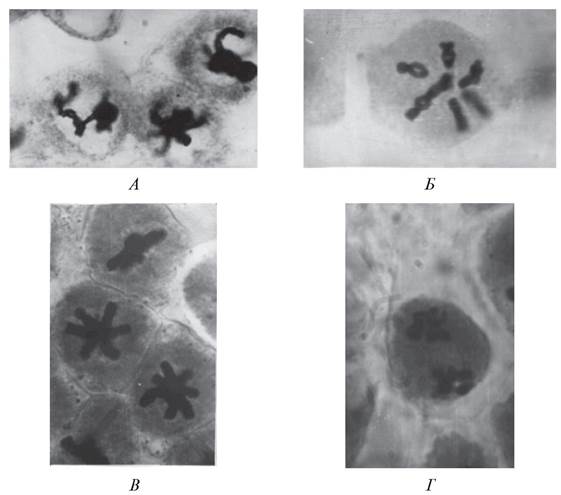
Процесс микроспорогенеза начинается с редукционного (мейотического) деления ядра микроспороцита (рис. 168), вслед за которым происходит цитокинез, т. е. формирование клеточных перегородок. В результате редукционного деления из каждого диплоидного микроспороцита образуется тетрада гаплоидных микроспор.
Заложение клеточных перегородок при образовании микроспор может быть сукцессивным (последовательным) и симультанным (одновременным).
Рис. 168. Мейоз в ядрах микроспороцитов бобов обыкновенных (Vicia faba): А, Б—стадии профазы I; В—метафаза I; Г—метафаза II
При сукцессивном типе развития образование клеточных перегородок в процессе мейоза идет в два этапа. В конце мейоза-1 формируется клеточная перегородка между дочерними ядрами разделившегося ядра микроспороцита и возникает диада. В мейозе-2 делятся ядра диады, после чего между вновь образовавшимися ядрами формируется перегородка и образуется типичная тетрада.
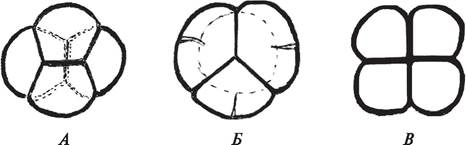
При симультанном типе развития микроспор цитокинез происходит в конце мейоза-2. Развитие перегородок в этом случае идет от периферии к центру и четыре клетки образуются одновременно. Таким образом, каждая микроспора в тетраде одета собственной оболочкой, а вся тетрада окружена каллозной оболочкой материнской клетки (рис. 169). Микроспоры в тетраде расположены различно, что обусловлено направлением веретена деления (ахроматинового веретена) и определяет тип тетрад. Наиболее часто при микроспорогенезе формируются тетраэдрические и изобилатеральные, реже крестообразные, тетрады (рис. 170). Очень редко образуются Т-образные и линейные тетрады.
Рис. 169. Тетраэдрическая тетрада микроспор бобов обыкновенных (Vicia faba): 1—микроспора; 2—ядро микроспоры с ядрышком; 3—оболочка микроспоры; 4—клетка тапетума; 5—ядро клетки тапетума; 6—оболочка тетрады микроспор
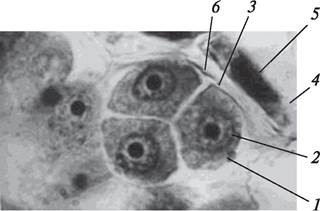
Рис. 170. Типы тетрад микроспор: А—крестообразная тетрада вереска обыкновенного (Calluna vulgaris); Б—тетраэдрическая брусники (Vaccinium vitis-idaea); В— изолатеральная гороха посевного (Pisum sativum)
У большинства растений тетрады существуют недолго. Вскоре после образования тетрады оболочка микроспороцита (материнской клетки микроспор), окружавшая тетраду, разбухает, ослизняется и разрушается, а тетрада распадается на отдельные микроспоры.
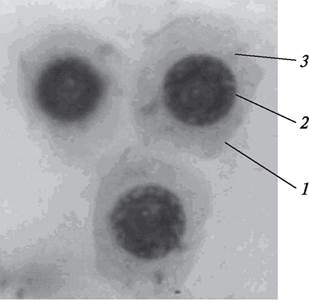
Каждая микроспора является гаплоидной клеткой с типичным для клетки строением. Микроспора имеет округлую форму, одета первичной клеточной оболочкой, содержит густую невакуолизированную цитоплазму, ядро, расположенное в центре, и все органоиды (рис. 171). С обособлением микроспор процесс микроспорогенеза заканчивается и здесь же в микроспорангии начинается новый процесс — прорастание микроспор и образование мужского гаметофита — пыльцевых зерен (пыльцы).
Рис. 171. Микроспоры бобов после распадения тетрады: 1—оболочка микроспоры; 2—ядро; 3—цитоплазма
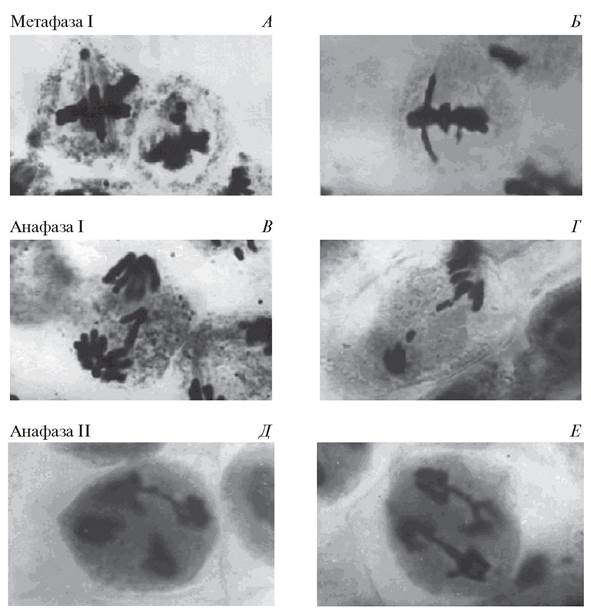
Каждая микроспора потенциально способна развиваться в мужской гаметофит только в том случае, если она образовалась в результате нормально осуществившегося процесса редукционного деления. Однако иногда в мейозе наблюдаются различного рода нарушения. Наиболее часто нарушения связаны с неправильным расхождением хромосом в метафазе и анафазе мейоза-1 и мейоза- 2 (рис. 172). Нарушения в мейозе приводят к образованию микро - спор с более или менее выраженными дефектами. Такие микроспоры или во - обще не способны к дальнейшему развитию, или из них формируется деформированная и стерильная пыльца.
Рис. 172. Нарушения мейоза при микроспорогенезе бобов: А—выброс хромосом из метафазной пластинки; Б—«забегание» хромосом; В—отставание хромосом; Г— разрыв хромосомного моста и образование фрагментов; Д, Е—образование хромосомных мостов
7.6.3. Образование мужского гаметофита (пыльцы). Морфологические и физиолого-биохимические особенности пыльцы
Первым признаком начала развития мужского гаметофита является изменение формы микроспоры.
Первоначально округлая микроспора становится эллиптической, но ядро в ней, как и прежде, расположено в центре. Постепенно развивающаяся микроспора увеличивается в размерах, в ее цитоплазме возникает крупная вакуоль, вследствие чего ядро смещается к одному из полюсов.
Мужской гаметофит образуется в результате митотического деления ядра микроспоры с последующим своеобразным формированием клеток. Митотическое деление ядра развивающейся микроспоры происходит типично с прохожденим всех фаз (рис. 173), однако нетипичным является ахроматиновое веретено — оно асимметричное. Ахроматиновые нити веретена, направленные к экваториальной зоне развивающегося пыльцевого зерна, значительно длиннее ахроматиновых нитей, направленных к полюсу. Следствием формирования асимметричного ахроматинового веретена является образование двух различных в функциональном и морфологическом смысле ядер. Ядро, которое расположено в центральной части разросшейся клетки, называется вегетативным, ядро, образовавшееся вблизи оболочки, — генеративным. Несмотря на то что эти ядра формируются в результате митотического деления, и, казалось бы, должны были обладать одинаковыми свойствами, они различаются по морфологическим, физиологическим и функциональным признакам. Вегетативное ядро крупнее генеративного, оно слабее окрашено реактивом Фельгена (специфическая реакция на ДНК), чем генеративное, что свидетельствует о различном содержании в них нуклеиновых кислот. Генеративное ядро более плотное, чем вегетативное.
Рис. 173. Образование мужского гаметофита (пыльцевого зерна, пыльцы) бобов обыкновенных (Vicia faba): А—Д — последовательные стадии митоза при делении ядра микроспоры: А—микроспора перед началом митоза (вид с экватора); Б—профаза; В—метафаза (вид с полюса); Г—анафаза; Д—телофаза; Е—образование вегетативного и генеративного ядер; Ж—образование вегетативной и генеративной клеток; З—двуклеточная пыльца бобов: 1—вегетативное ядро; 2—генеративное ядро; 3—вегетативная клетка; 4—генеративная клетка
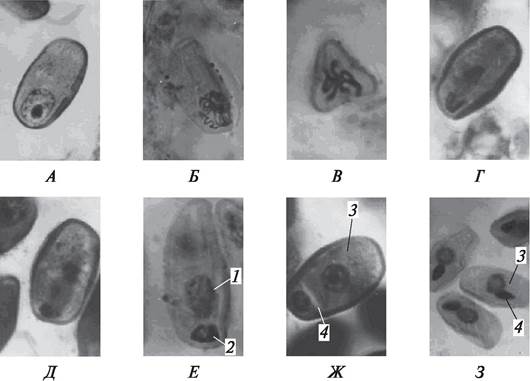
Вскоре после образования между вегетативным и генеративным ядрами происходит расслоение цитоплазмы или, как предполагают некоторые эмбрио логи, формируется каллозная перегородка, и в результате возникают две неравные по размеру клетки: крупная — вегетативная и меньшая — генеративная.
Первое описание двух неравных клеток пыльцевого зерна было дано немецким ботаником К. Негели (1818—1891) в 1842 г. Название меньшей клетке — «генеративная» — дал Э. Страсбургер, он же определил ее функцию.
Вегетативная и генеративная клетки отличаются не только по размерам. Согласно имеющимся данным, вегетативная клетка является запасающей с высокой метаболической активностью. В ней содержится большое количество липидов, крахмала, белков, которые необходимы при дальнейшем развитии пыльцы, в первую очередь при росте пыльцевой трубки. По содержанию белков вегетативная клетка богаче генеративной, ее белки характеризуются большей кислотностью, причем преобладает аргинин. Синтез ДНК в ядре вегетативной клетки происходит значительно позднее, чем в ядре генеративной, чаще всего перед созреванием пыльцы. В вегетативной клетке наблюдается интенсивный синтез цитоплазмы, что приводит к исчезновению вакуоли. Перед созреванием вегетативная клетка обезвоживается.
Генеративная клетка содержит небольшое количество цитоплазмы, но имеет все клеточные органоиды.
В цитоплазме генеративной клетки отсутствуют запасные вещества, в частности крахмал. В ядре генеративной клетки быстро удваивается количество ДНК, а накопление белков происходит медленно. При формировании генеративной клетки сначала резко снижается содержание РНК, но затем происходит интенсивный синтез ее в цитоплазме и ядре и одновременно снижается содержание РНК в ядрышке.
Ультраструктуру вегетативной и генеративной клеток начали изучать с 60 - х гг. ХХ в. Как установлено, обе клетки имеют все клеточные органоиды, однако есть и некоторые отличия. В цитоплазме вегетативной клетки особенно много пектиновых пузырьков, которые принимают участие в формировании оболочки пыльцевой трубки. В цитоплазме генеративной клетки иногда нет пластид.
Вскоре после образования вегетативной и генеративной клеток ядро генеративной клетки претерпевает сильное сжатие, так что различить его внутреннее строение невозможно, форма ядра становится эллиптической, а цитоплазма располагается вокруг него тонким слоем. Вследствие сжатия генеративной клетки увеличиваются размеры вегетативной. С образованием вегетативной и генеративной клеток процесс формирования мужского гаметофита не заканчивается. Генеративная клетка начинает перемещаться в экваториальную область развивающегося мужского гаметофита, внедряется в цитоплазму вегетативной и располагается вблизи ядра вегетативной клетки, принимая серповидную или линзовидную форму (см. рис. 173, З).
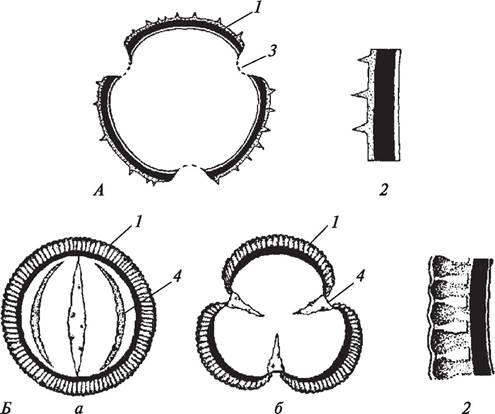
Параллельно с преобразованием внутренней структуры микроспоры, превращающейся в мужской гаметофит, претерпевает изменение и ее оболочка. Постепенно над цитоплазматической мембраной образуется целлюлозная оболочка, пропитанная спорополленином. Таким образом, пыльцевое зерно оказывается «одетым» двумя оболочками: наружной — экзиной и внутренней — интиной. Экзина — оболочка не сплошная, в ней имеются апертуры, представляющие перерывы в экзине (рис. 174). Экзина очень прочная оболочка, благодаря ей пыльца способна не только поддерживать постоянство своей формы, но и длительное время сохранять жизнеспособность (способность к прорастанию).
Рис. 174. Схема строения пыльцевого зерна: А—трехпоровая пыльца колокольчика персиколистного (Campanula persicifolia); Б—трехбороздная пыльца брюквы (Brassica napus) (а — вид с экватора; б — вид с полюса): 1—экзина; 2—строение экзины; 3—апертуры в виде пор; 4—апертуры в виде борозд
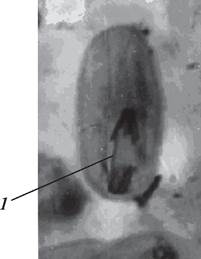
Однако иногда при развитии пыльцевых зерен наблюдаются нарушения митотического деления ядра микроспоры. Наиболее уязвимой стадией митоза является анафаза. Из-за атипичного расхождения хромосом в анафазе могут формироваться так называемые хромосомные мосты (рис. 175). Если происходит их разрыв, то в пыльце вместо двух нормально развитых ядер могут сформироваться микроядра. Пыльца с микроядрами чаще всего нежизнеспособна или с пониженной жизнеспособностью.
Рис. 175. Нарушение митоза при образовании мужского гаметофита бобов обыкновенных (Vicia faba): 1—образование хромосомного моста в анафазе митоза
У двудольных растений развитие пыльцы в пыльнике заканчивается на стадии образования двух клеток: вегетативной и генеративной. Пыльца, которая состоит из вегетативной и генеративной клеток, называется двуклеточной пыльцой, или двуклеточным мужским гаметофитом. Образование гамет — спермиев — у такой пыльцы происходит только после того, как она попадет на рыльце пестика и начнет прорастать. Тогда ядро генеративной клетки разделится митотически и в результате последующего цитокинеза образуются две клетки — спермии.
У однодольных растений генеративная клетка претерпевает митотическое деление, когда пыльца еще находится в пыльцевой камере. В результате в такой пыльце образуется вегетативная клетка и два спермия. Такая пыльца называется трехклеточной пыльцой, или трехклеточным мужским гаметофитом.
Еще в 40-х гг. ХХ в. профессор Киевского университета В. В. Финн (1878— 1957) установил, что спермии представляют собой настоящие клетки и, по его терминологии, могут быть «богатоплазменными» и «бедноплазменными». В настоящее время выявлено, что цитоплазма в спермиях распределена тонким слоем вдоль латеральных стенок и сконцентрирована на полюсах. Хроматин ядра находится в конденсированном состоянии, ядрышко в ядре, как правило, отсутствует. В спермиях содержатся все клеточные органоиды, за исключением пластид. По форме спермии могут быть диморфными (различающимися по морфологическим особенностям) и изоморфными (имеющими одинаковое строение). По представлениям С. Г. Навашина, оба спермия являются энантиоморфными (зеркально подобными).
Пыльца покрытосеменных растений разнообразна по размерам, форме, числу апертур, строению экзины (рис. 176). Размеры пыльцы варьируют от 10 мкм (род фикус — Ficus) до 400 мкм (сем. Тыквенные — Cucurbitaceae, Мальвовые — Malvaceae).
Рис. 176. Строение пыльцы у различных представителей покрытосеменных: А—дуб черешчатый (Quercus robur); Б—клен гиннала (Acer ginnala); В—подмаренник мягкий (Galium mollugo); Г — нивяник обыкновенный (Leucanthemum vulgare); Д—дрема белая (Melandrium album); Е—кубышка малая (Nuphar pumilum); Ж—звездчатка ланцетовидная (Stellaria holostea); З—иван-чай узколистный (Chamaenerion angustifolium); И—береза повислая (Betulapendula); К—ольха черная (Alnusglutinosa): 1—вид с полюса; 2—вид с экватора
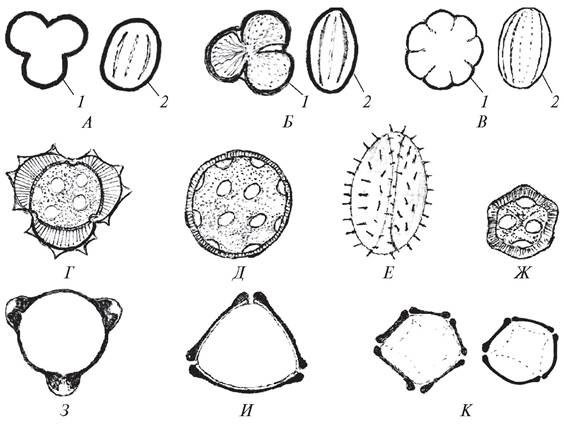
Пыльца может быть округлой (сем. Гвоздичные, Маревые), треугольной (орешник обыкновенный — Corylus avellana), эллиптической (сем. Бобовые). У водных растений пыльца нитевидно вытянутая.
Количество апертур в оболочке пыльцевых зерен варьирует в широких пределах. Среди покрытосеменных преобладает одноапертурная (сем. Лилейные — Liliaceae, Злаки — Gramineae) и трехапертурная (сем. Бобовые — Fabaceaе) пыльца. У гвоздичных, маревых пыльцевые зерна глобально-апертурные, т. е. апертуры расположены по всей поверхности пыльцевого зерна (см. рис. 176). Очертания апертур могут быть округлыми или продолговатыми, поэтому различают пыльцевые зерна порово-апертурные (с округлыми апертурами) и бороздно-апертурные (с продолговатыми апертурами).
Через апертуры происходит прорастание пыльцевого зерна, что связано с образованием пыльцевой трубки. Обычно пыльцевая трубка выходит через одну из апертур, но иногда может наблюдаться и полисифония, однако при образовании нескольких пыльцевых трубок содержимое пыльцевого зерна все равно перемещается только в одну из них (рис. 177).
Рис. 177. Явление полисифонии: А—у бобов обыкновенных (Vicia faba); Б—у гороха посевного (Pisum sativum): 1—пыльцевые трубки; 2—апертуры в экзине

Строение экзины различно, что связано с характером опыления растений. У ветроопыляемых растений экзина гладкая, у насекомоопыляемых — скульптурированная (см. рис. 174). На поверхности экзины перекрестноопыляемых растений формируются различные выросты, шипики, бороздки, что облегчает прикрепление пыльцевых зерен к переносчику пыльцы (насекомым, птицам и т. д.).
Для пыльцы характерна довольно высокая морфологическая видоспецифичность, что находит практическое применение в геологии, а также в палеоботанике (метод спорово-пыльцевого анализа), пищевой промышленности (определение состава меда), в медицине (анализ на аллергию).
В последние годы началось изучение пыльцы различных растений (кукурузы, томата, рапса, мятлика, амброзии, петунии, табака и др.) с целью выявления специфических генов пыльцы и установления их предполагаемой функции. Установлено, что в зрелой пыльце разных видов содержится от 20 тыс. до 24 тыс. генов, т. е. около 60 % генов, содержащихся в побеге.
Еще раньше было установлено, что пыльца содержит различные неорганические и органические вещества. Из неорганических веществ в пыльце выявлены Na, К, Са, Р, Fe, Mg, Сu, Аl, S, Мn. В пыльце обнаружены различные сахара (сахароза, глюкоза, мальтоза, фруктоза), липиды, каротиноиды, около 20 аминокислот. По относительному содержанию витаминов пыльца превосходит все остальные части растения. В пыльце обнаружены витамин С, витамины группы В, Е, РР и другие. О высокой физиологической активности пыльцы свидетельствует наличие 32 ферментов, гетероауксина, соединений, содержащих SH-группы.
Высокое содержание витаминов позволило в некоторых случаях использовать пыльцу растений как витаминоносное средство. В то же время пыльца многих растений (злаков, амброзии) вызывает аллергию, результатом которой являются дерматиты, слезотечение, насморк, затруднение дыхания и другие последствия, требующие медикаментозного лечения.