ФІЗІОЛОГІЯ РОСЛИН З ОСНОВАМИ БІОХІМІЇ - Притуляк Р. М. - 2016
Опорний конспект лекцій
ТЕМА № 1. ВСТУП. ФІЗІОЛОГІЯ РОСЛИННОЇ КЛІТИНИ
ВСТУП
План
1. Предмет і завдання фізіології рослин. Взаємозв’язок фізіології рослин з іншими біологічними дисциплінами.
2. Методи фізіології рослин.
3. Фізіологія рослин як теоретична основа агрономічних наук.
4. Коротка історія розвитку фізіології рослин як науки і роль вітчизняних вчених у її розвитку.
5. Основні напрямки розвитку сучасної фізіології рослин.
1. Предмет і завдання фізіології рослин.
Сучасна фізіологія рослин є інтегративною дисципліною, яка вивчає головні життєві функції рослинного організму на різних рівнях їх організації, вивчає закономірності життя рослин та шляхи керування ними з метою оптимізації продуктивності культурних рослин. Предметом фізіології рослин є функції живих рослинних організмів, їх органів, тканин, клітин та клітинних компонентів, а також причини тих або інших проявів їхньої життєдіяльності, їх залежність від зовнішніх і внутрішніх чинників. Методологія фізіології рослин заснована на уявленнях про рослинний організм як складну саморегулюючу систему, яка включає ієрархію різних структурних рівнів — від субклітинних, тобто макромолекул до цілісної рослини. Пізнання фізіологічних функцій здійснюється через дослідження простих рівнів організації, з наступною інтеграцією даних при розгляді фізіологічних систем зростаючої складності.
Метою фізіології рослин є пізнання закономірностей життєвих функцій рослин, розкриття їх механізмів, формування уявлення про структурно- функціональну організацію рослинних систем різних рівнів та вироблення шляхів керування рослинним організмом.
Живим системам, в тому числі рослинному організму, притаманна вища форма цілісності, нерозривний зв'язок та взаємозумовленість складових його компонентів. Одним із проявів цілісності є те, що незважаючи на велике значення кожної окремої функції, життєдіяльність організму в цілому залежить від того, яким чином конкретна функція пов'язана з іншими, і як ці співвідношення узгоджені зі зміною умов оточуючого середовища.
Виняткова специфічність хімічного складу, морфологічної будови, тісний взаємозв'язок структури і функції, залежність процесів обміну речовин від стану структур, динамічність останніх - такі специфічні властивості об'єкта досліджень фізіології рослин.
Проблеми та завдання фізіології рослин настільки широкі та складні, що вирішення їх потребує застосування цілого комплексу тонких фізико-хімічних методів, різноманітних експериментальних та теоретичних підходів. Саме тому ця дисципліна входить у ранг точних фундаментальних наук.
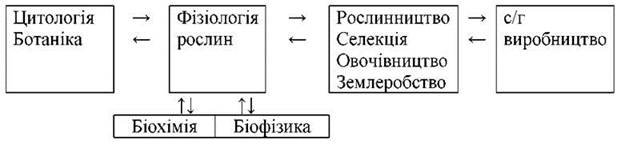
Сучасна фізіологія рослин має тісний взаємозв’язок з ботанікою, цитологією, біохімією, біофізикою, генетикою, імунологією, екологією.
2. Методи фізіології рослин.
Основний метод пізнання процесів, явищ в фізіології - експеримент, дослід. Відповідно, фізіологія рослин - наука експериментальна.
Для дослідження фізико-хімічної суті функцій та процесів у фізіології рослин широко застосовують наступні методи:
· Лабораторно-аналітичний;
· Вегетаційний (у контрольованих умовах);
· Польовий;
· Виробничий;
· Мічених атомів;
· Електронної мікроскопії;
· Ультрафіолетової та люмінесцентної мікроскопії;
· Електрофореза;
· Хроматографії.
Крім цього, використовують фітотрони та лабораторії штучного клімату, в яких вирощують рослин та проводять досліди в умовах певного складу повітря, потрібної температури та освітлення.
За допомогою цих методів фізіологи досліджують рослини на молекулярному, субклітинному, клітинному та організменному рівнях.
3. Фізіологія рослин як теоретична основа агрономічних наук.
Фізіологія рослин є теоретичною основою всіх агрономічних наук, оскільки управління життєвими процесами рослин та їх використання складають основне завдання фізіології рослин.
4. Коротка історія розвитку фізіологи рослин як науки і роль вітчизняних вчених у її розвитку.
Сучасний стан фізіології рослин є результатом довгого шляху розвитку, що пройшла ця наука за більш ніж двохсотліття її існування. Сформувалася вона в XVІІ-XVІІІ ст. як один із напрямів ботаніки.
Науковий підхід до фізіології рослин започаткував голландець Ян Батіст ван Гельмонт (1579-1644), який провів перший кількісний вегетаційний дослід задля вивчення шляхів живлення рослин. На основі своїх дослідів він зробив помилковий висновок про те, що вода є єдиним поживним субстратом для росин. І тільки пізніше було встановлено важливість ґрунту та повітря к джерел живлення рослин. Можна без перебільшення стверджувати, що своє існування фізіологія рослин розпочала з науки про живлення.
М. Мальпігі (1628-1694) - один із засновників мікроскопічної анатомії рослин, відкриття щодо певних мікроструктур поєднував з їхніми функціями, що мало важливе значення для майбутньої фізіології рослин.
С. Гейлс (англійський ботанік і хімік) у 1727 р. висловив припущення щодо можливості живлення рослин за участю листків із повітря. С. Гейлса, без сумніву, слід віднести до засновників фізіології рослин як науки.
Фізіологія рослин як наука сформувалася майже на століття пізніше, коли в 1771-1782 рр. англієць Дж. Прістлі, австрієць Я. Інгенхауз і швейцарець Ж. Сенеб’є виявили компоненти, за рахунок яких відбувається повітряне живлення рослин.
Н. Соссюр (1777-1845) шляхом точного хімічного аналізу довів, що рослини на світлі засвоюють вуглець із СО2. Так поступово складалось уявлення про фотосинтетичну функцію рослин.
Термін «фотосинтез» німецький учений І. Пфеффер запропонував лише в 1877 р. Після відкриття фотосинтезу, а також законів збереження матерії та енергії фізіологія рослин все частіше розглядала повітряне середовище й сонячне світло як основні матеріальні та енергетичні джерела існування зеленої рослини.
Як самостійна галузь науки фізіологія рослин остаточно сформувалась у 1800 р., коли Ж. Сенеб’є (1742-1803) ввів термін «фізіологія рослин». Він написав перший підручник з цієї дисципліни, де сформулював суть предмета, методи та завдання даної Науки.
У Росії фізіологія рослин довго не відокремлювалась у самостійну дисципліну, і лише в 1863 р. новим університетським статутом були створені кафедри фізіології рослин. Першим російським ботаніком-фізіологом треба вважати С. О. Рачинського (1833-1902) - професора фізіології рослин Московського університету.
З 1872 р. у цьому ж університеті почав працювати видатний учений К. А. Тімірязєв (1843—1920). У Петербурзькому університеті з 1861 р. курс фізіології рослин викладав проф. А. С. Фамінцин (1835—1918). На першому етапі розвитку фізіологія рослин була наукою університетською.
Перший російський підручник з фізіології рослин у 1887 р. написав проф. А. С. Фамінцин, а в 1891 р. вийшов у світ підручник В. І. Палладіна, який перевидавався дев'ять разів.
Вагомий внесок у розвиток різних розділів фізіології рослин у XIX ст. внесли: Ж. Буссенго, А. С. Фамінцин, К. А. Тімірязєв, М. С. Цвєт (фотосинтез); І. П. Бородін, О. М. Бах, Г. Бертран, В. І. Палладій, Л. Пастер (дихання); Г. Дютроше, Г. Де Фріз, Ю. Сакс (водний режим); Ю. Лібіх, Г. Гельрігель, Й. Кноп, С. М. Виноградський, М. Бейєрінк, Д. М. Прянишников (мінеральне живлення); Й. В. Баранецький, Г. Фехтінг, Г. Клебс (ріст і розвиток рослин) та ін.
На початку XX ст. відбулась остаточна диференціація фізіології рослин на основні її розділи, причому деякі з них набули такого значення, що в перші десятиріччя перетворилися на самостійні дисципліни з власними теоретичними основами та широким впровадженням у практику: 1902 р. - вірусологія, 1910 р. - агрохімія, 1930 р. - мікробіологія та біохімія.
У цей період значного успіху досягнуто в розкритті біохімічних механізмів дихання (В. І. Палладін, Г. Віланд, С. П. Костичев, О. Варбург, Д. Кейлін, Т. Тунберг, Г. Кребс, А. Корнберг), фотосинтезу (Р. Вільштеттер, К. ван Ніль, Р. Хілл, М. Кальвін, Д. Арнон), дослідженні ендогенних регуляторів росту (М. Г. Холодний, Ф. Вент, Ф. Кегль, Й. Куросава, Т. Ябута, Ф. Скуг), вивченні механізмів стійкості рослин (М. О. Максимов), мінерального живлення (Д. А. Сабінін).
У 1934 р. було створено Інститут фізіології рослин АН СРСР, який координував усі дослідження в даній галузі науки і в якому плідно працювали А. Л. Курсанов, А. А. Ничипорович, М. X. Чайлахян, П. О. Генкель, І. І. Туманов та інші вчені.
Наступником і продовжувачем традицій фітофізіології на переломному етапі, коли в середині 50-х років фізіологія рослин в усьому світі піднялася на новий рівень, став академік А. Л. Курсанов, який, по суті, дав нову орієнтацію розвитку фізіології рослин, створивши відому курсанівську школу (А. Т. Мокроносов, О. М. Кулаєва, Р. Г. Бутенко, В. Ю. Семененко, Д. Б. Вахмістров, Б. Б. Вартапетян, Ю. Г. Молотковський, А. Т. Верещагін, І. А. Тарчевський, Р. К. Саляєв, Г. О. Санадзе, В. В. Польовий та ін).
З'явилися нові напрями наукових пошуків: кисневий обмін і транспортування кисню (Б. Б. Вартапетян), цитокініни та їхнє значення в регуляції метаболізму (О. М. Кулаєва), донорно-акцепторні системи рослин (А. Т. Мокроносов), організація рослинних мембран (Ю. Г. Молотковський, Р. К. Саляєв); іонний гомеостаз рослинної клітини (Д. Б. Вахмістров), метаболізм рослинних ліпідів (А. Т. Верещагін).
Інтенсивно досліджувалися механізми регуляторної дії фітогормонів, включаючи їхню рецепцію та передачу гормональних сигналів на генетичний апарат клітини (О. М. Кулаєва, В. В. Польовий), метаболізм фітогормонів і регуляція росту (В. І. Кефелі). Розпочались дослідження імунохімії рослин (О. Д. Володарський).
В кінці XX ст. змінилася техніка фізіологічних експериментів, проводяться фундаментальні дослідження, на основі яких народжуються принципово нові біотехнології рослин. Настав новий етап розвитку фізіології рослин, який характеризується процесом асиміляції та інтеграції даною наукою досягнень інших експериментальних біологічних наук.
5. Основні напрямки розвитку сучасної фізіології рослин.
Як фундаментальна біологічна наука і теоретична основа рослинництва, вона вирішує дві головні взаємопов'язані проблеми:
1) всебічне вивчення метаболічних процесів рослинного організму з метою пізнання суті життя - основного теоретичного питання біології;
2) вивчення закономірностей життєдіяльності рослин в онтогенезі у різних екологічних умовах з метою розв'язання практичних завдань рослинництва.
Для вирішення цих проблем фізіологія рослин має наступні наукові напрями.
Біохімічний - вивчає природу та функціональне значення органічних речовин, що входять до складу рослин, шляхи їх утворення і перетворення під час фотосинтезу та дихання; досліджує особливості кореневого живлення рослин та ряд інших питань.
Біофізичний - вивчає шляхи накопичення енергії у клітинах рослин, природу біологічних мембран, біофізичні процеси фотосинтезу і дихання, фізико-хімічні закономірності водного режиму і кореневого живлення, регуляторні системи фітохромів, природу подразнень та ін.
Синтетичний, або кібернетичний - вивчає енергетичний стан і кінетику таких взаємопов'язаних процесів, як фотосинтез і дихання, живлення та органоутворення, загальні закономірності росту рослин.
Екологічний - досліджує вплив зовнішнього середовища на процеси життєдіяльності у рослині, розробляє ефективні засоби впливу на рослину (застосування добрив, створення оптимального водного, світлового і повітряного режимів та ін.) для забезпечення її стійкості до хвороб і шкідників, формування врожаю тощо.
Онтогенетичний - вивчає вікові закономірності розвитку рослин і розробляє шляхи раціонального керування цим процесом (світлокультура, яровизація, загартовування, фотоперіодизм та ін.).
Еволюційний - вивчає особливості індивідуального виду, сорту рослин у відповідних екологічних умовах, зміни структури рослин залежно від факторів зовнішнього середовища, розглядає онтогенез як функцію генотипу.
ФІЗІОЛОГІЯ РОСЛИННОЇ КЛІТИНИ
План
1. Клітина як функціональна одиниця живої матерії.
2. Структурна організація клітини.
3. Клітинна стінка, її склад, будова і функції.
4. Цитоплазма, її будова і властивості.
5. Ядро, його будова і функції.
6. Біологічні мембрани.
7. Ендоплазматична сітка.
8. Рибосоми, їх хімічний склад та функції.
9. Хлоропласти, їх будова та функція.
10. Мітохондрії, їх будова та функції.
1. Клітина як функціональна одиниця живої матерії.
Структурною та функціональною одиницею рослинного організму, як і інших живих істот, є клітина. Вперше цей термін запропонував у 1665 р. англійський вчений Роберт Гук, який вивчав під мікроскопом структуру корка та помітив утворення, що мали вигляд бджолиних сотів, які він і назвав клітинами. Пізніше, вже в 1838-1839 р., німецькі вчені гістолог Т. Шванн і ботанік М. Шлейден запропонували загально-біологічну концепцію клітинної теорії, згідно з якою основною одиницею структури і функції всіх організмів є клітина. В основу цієї теорії було покладено два фундаментальні положення:
1) Клітина є основною одиницею всіх організмів;
2) Ріст, розвиток і диференціація тканин живих організмів зумовлені процесом утворення нових клітин.
Ця теорія була розвинена і вдосконалена Рудольфом Вірховим (1885 р.), який увів ряд нових положень про роль клітинних структур у організмі. Основним положенням його теорії є: поза клітиною не існує життя і кожна клітина походить із клітини. Це було підтверджено подальшим розвитком біології.
За сучасним уявленням, клітина — це основна структурно-функціональна одиниця всіх живих організмів, елементарна жива система. Лише віруси являють собою неклітинні форми життя.
Рослинна клітина є функціональною структурною одиницею живої матерії, так як для неї властиві наступні особливості:
· Обмін речовин та енергії;
· Здатність до росту та самовідтворення;
· Збереження та передача спадкової інформації.
Сталість клітинної структури в усьому органічному світі зумовлена тим, що тільки вона забезпечує найкраще зберігання, репродукцію і передачу спадкової інформації. Зі структурою пов'язана здатність організмів зберігати, переносити енергію і перетворювати її у роботу.
2. Структурна організація рослинної клітини.
Згідно сучасних уявлень, рослинна клітина складається з трьох основних частин - оболонки, протопласта і вакуолі. Клітинна оболонка відносно жорстка, складна у хімічному відношенні і є продуктом діяльності протопласта. Протопласт, тобто жива частина клітини, є колоїдним розчином із розміщеними у ньому структурними компонентами (ядро, пластиди, мітохондрії, ендоплазматична сітка, комплекс Гольджі, рибосоми). Вакуолі - це неживі утворення, заповнені розчинами неорганічних солей, поглинутих клітиною, і органічних речовин - продуктів метаболічної діяльності клітини. Розміри клітин варіюють дуже широко (від одного до декількох сотень мікронів). Морфологічні відмінності клітин зумовлені характером біологічних процесів і типом обміну речовин. Якщо рослинна клітина вирощується ізольовано, то її форма наближається до сферичної, якщо в оточенні інших клітин, то може набувати форму багатогранника. Клітини зони росту стебла або кореня за формою нагадують коробочку завдовжки близько 50 мкм, завширшки 20 мкм і висотою 10 мкм. Клітини ембріональних тканин і конуса наростання дуже малих розмірів.
У молодих клітин близькі поздовжні і поперечні розміри, тонка оболонка. Вони повністю заповнені протопластом. З часом, зі збільшенням об'єму клітини, кількість протопласта поступово зростає, у ньому утворюється багато малих вакуоль, які поступово зливаються в одну велику; оболонка потовщується.
Протопласт ззовні і зсередини оточений поверхневими біологічними мембранами: від клітинної стінки його відокремлює плазмалема, від вакуолі - тонопласт.
Протопласт і клітинна оболонка не ізольовані повністю. Вони сполучаються за допомогою спеціалізованих отворів - пор. Через пори в оболонці за допомогою цитоплазматичних тяжів (плазмодесм) протопласт одних клітин з'єднується з протопластом інших. Таким чином, завдяки мембранним утворенням і плазмодесмам усі клітини об'єднані. Плазмодесми за своєю будовою нагадують трубочки діаметром від 20 до 100 нм. У кожній плазмодесмі існує канал (десмотрубочка), по якому різні речовини можуть переходити від однієї клітини до іншої.
3. Клітинна стінка, її склад, будова і функції.
Оболонка клітини утворюється з продуктів секреторної діяльності протопласта, які послідовно нашаровуються у процесі розвитку клітини. Головною складовою клітинної стінки є целюлоза (клітковина - (С6Н10О5)n). Молекули целюлози - це нерозгалужені ланцюжки, які складаються із залишків D-глюкопіранози, поєднаних між собою глікозидними (В1-4)-зв’язками. У середньому одна молекула целюлози містить до 8000 залишків глюкози.
Між оболонками сусідніх клітин існує серединна пластинка, яку утворюють спочатку студневидні пектинові речовини, але пізніше вона доповнюється целюлозою та іншими полісахаридами і набуває жорсткості. У здерев'янілих клітин оболонка насичена лігніном.
Первинна оболонка клітини складається з целюлозних мікрофібрил, занурених у основну речовину - матрикс. Завдяки проміжкам між фібрилами клітинна стінка має достатню гнучкість.
Здатність клітинної стінки розтягуватися залежить від характеру розміщення целюлозних фібрил. При безладному їх розміщенні оболонка розтягується рівномірно в усіх напрямах. Якщо ж фібрили розміщені паралельно, то розтягування відбувається під прямим кутом до осі. Під час формування клітини зовнішня оболонка зазнає сильного тиску з боку протопласта, розтягується, і при цьому до неї додається новий будівельний матеріал - відбувається її ріст і потовщення. У деяких клітин, наприклад мезофілу, формування клітинної оболонки завершується, як тільки клітина досягає своєї максимальної величини. В інших тканинах у клітин, що закінчили свій ріст, із внутрішнього боку утворюється вторинна клітинна оболонка із жорсткою структурою. Таке потовщення оболонки скорочує об'єм протопласта. З часом весь протопласт відмирає повністю, залишаються лише порожнисті циліндри з клітинних оболонок, що виконують механічну функцію або функцію провідних тканин.
Вторинна клітинна оболонка пронизана численними порами. Дільниця оболонки з порами досить тоненька, складається лише із серединної пластинки і первинної клітинної стінки.
У зрілих живих клітин пори представлені канальцями, що з'єднують внутрішню частину вторинної клітинної стінки і зовнішню частину первинної. Пори можуть розміщуватись групами, утворюючи порові поля, які відіграють важливу роль у пропусканні води, розчинів мінеральних і пластичних речовин.
Функції клітинної оболонки. Клітинна оболонка є секреторним продуктом протопласту і в зрілій клітині, ймовірно, нежива. Однак в ній є білки, серед яких і ті, що мають ферментативну активність.
Клітинна оболонка забезпечує окремим клітинам і рослині в цілому механічну міцність і опору.
Вона визначає розмір, форму та стабільність рослинної клітини, захищає протоплазматичну мембрану від руйнування в разі дії гідростатичного тиску, що формується всередині клітини.
Клітинна оболонка є протиінфекційним бар'єром, бере участь в поглинанні, транспортуванні та виділенні речовин. Систему поєднаних одна з одною клітинних оболонок називають апопласт, який слугує головним шляхом для пересування води і розчинених речовин по рослині. Через пори в клітинних оболонках проходять цитоплазматичні тяжі, які зв'язують вміст окремих клітин, тобто об'єднують всі протопласти в одну систему — симпласт.
Є дані, що вуглеводні компоненти клітинної оболонки, взаємодіючи із фітогормонами, викликають фізіологічні зміни у клітині.
4. Цитоплазма, її будова і властивості.
Протопласт - це колоїдна система. У живій клітині вона перебуває у постійному русі, завдяки якому забезпечується оптимальне розміщення органел, краще протікання біохімічних реакцій, видалення продуктів обміну у вакуолю і за межі клітини тощо. Основу протопласта складає цитоплазма, також сюди входить ядро.
Цитоплазма - напіврідка прозора і в’язка гомогенна маса, розташована під клітинною оболонкою переважно у вигляді відносно тонкого пристінного шару.
Хімічний склад цитоплазми (вода - 75-85%, білки та амінокислоти - 10-12%, вуглеводи - 4-6%, жири і ліпіди - 2-3%, інші органічні речовини - близько 1%, мінеральні речовини - 2-3%) сприяє утворенню колоїдного розчину, який не змішується з водою і речовинами вакуолей.
Цитоплазмі властива еластичність і досить висока густина. Наприклад, у клітинах паренхіми кори бобів її в'язкість у 24 рази вища, ніж у води.
В'язкість цитоплазми є неоднорідною. Периферійна частина її, яка прилягає до оболонки, більш в'язка і відділяється від неї поверхневою мембраною - плазмалемою. З боку вакуолі цитоплазма відокремлена другою поверхневою мембраною - тонопластом. Між цими мембранами знаходиться з меншою в'язкістю внутрішній шар цитоплазми - мезоплазма, яка є цитоплазматичним матриксом, що пронизаний ендоплазматичною сіткою (внутрішньою мембраною). У цитоплазматичному матриксі постійно відбуваються процеси обміну речовин. Вважається, що цитоплазма містить фібрилярні структурні елементи, які сприяють формуванню гелей.
У структурній і функціональній організації цитоплазми виключно важливе значення має вода. Властивості її, як розчинника і речовини, що має велике біологічне значення, визначаються особливостями її внутрішньо-молекулярної структури, насамперед, полярністю молекули. Ця полярність зумовлена несиметричним розміщенням електронів водню і кисню у молекулі і відповідно - нерівномірним розподілом позитивних і негативних зарядів.
5. Ядро, його будова і функції.
Центральною і найважливішою органелою клітини є ядро, яке має досить складну структуру. Форма його здебільшого кулеподібна або овальна. Розмір коливається у досить широких межах.
У ядрі зосереджена генетична інформація у відповідних структурних одиницях - хромосомах. їх переплетіння утворює цілісну масу - хроматин. Крім того, ядро має одне або кілька ядерець. Простір між ядерними структурами заповнює безбарвна речовина - каріоплазма, або нуклеоплазма.
Зверху ядро оточене мембраною пористої структури, що має рибосоми та за допомогою елементів ендоплазматичної сітки сполучена з мембранами інших клітинних структурних компонентів.
Внутрішня мембрана подекуди випинається всередину ядра. На 1 мкм ядерної оболонки може бути від 10 до 100 пор діаметром до 20 нм кожна. Пори ядра - динамічні структури, які періодично закриваються та відкриваються, регулюючи обмін речовин між ядром і цитоплазмою.
Ядро не лише вміщує генетичну інформацію, але й передає її до цитоплазми (синтез інформаційної РНК та ін.), від клітини до клітини (поділ ядра, поділ клітини, розмноження, успадкування). Серед білків ядра переважають нуклеопротеїди. Структура ядра залежить від функціонального стану клітини.
Внутрішня будова ядра змінюється залежно від його стану. Розрізняють два періоди в житті ядра — метаболічний (між поділом) та період поділу. В метаболічний період в ядрі є одне або декілька ядерець, які складаються з міцно переплетених ниток — нуклеонеми і містять до 80 % білка, 10... 15 % РНК і незначну кількість ДНК.
Ядро заповнене нуклеоплазмою і переплетеними, скрученими хроматиновими нитками. Хроматин складається із ДНК, гістонових і негістонових білків, незначної кількості РНК і ліпідів. Характерною рисою метаболічного періоду ядра є процес самовідтворення (подвоєння) молекул ДНК — реплікація. Лише після цього ядро переходить до поділу, тобто мітозу. В інтерфазному ядрі хромосоми мають вигляд досить безформених скупчень хроматину, що набубнявіли. Перед початком поділу ядра кожна хромосома складається з двох хроматид, які в анафазі діляться. Надалі у новій клітині хроматида подвоюється і є хромосомою з повним набором генетичної інформації.
Основна функція ядра — контроль за синтезом білків і життєдіяльністю клітини, збереження та передача генетичної інформації дочірнім клітинам у процесі поділу клітин. Ядро оточене подвійною ядерною мембраною і має в своєму складі хроматин, ядерця та нуклеоплазму.
Хромосоми, або хроматин, в еукаріотів складаються з чотирьох видів молекул: 1) ДНК (близько 35%); 2) РНК (близько 12%); 3) лужного низькомолекулярного білка - гістону (близько 40%); 4) кислого негістонового білка, у тому числі ферментів (близько 10%) і незначної кількості ліпідів, полісахаридів та іонів металів.
Ядерце - це округле утворення високої щільності, яке не має мембран. Воно складається з більш компактного, ніж ядро матеріалу, містить рибонуклеїнову кислоту (15%) і білки (80%). У ядерці містяться у великій кількості субодиниці рибосом. Ці рибонуклеопротеїдні гранули разом з рибонуклеопротеїдними ниткоподібними структурами (фібрилами) занурені у нуклеоплазму. У ядерцісинтезуються численні рибосомні білки-гістони, накопичується РНК перед виходом до цитоплазми. Ядерцева рибосомальна РНК і білки об'єднуються у рибосомні субодиниці. Крім того, у ядерці збираються інші типи РНК (транспортна РНК). Тому воно -: місцем перерозподілу РНК.
Основна функція ядерця — синтез ядерних білків. Ймовірно, що в них синтезується і певна кількість специфічних рибосомальних білків. У ядерці можливе також самоскладання рибосом. У разі руйнування ядерець, наприклад ультрафіолетовим випромінюванням, ядро втрачає здатність до поділу.
6. Біологічні мембрани.
Протопласт зовні та зсередини обмежений відповідними мембранами: плазмалема відокремлює його від клітинної оболонки, а тонопласт — від вакуолі. Розрізняють також мембрани ядра, мітохондрій, пластид, субодиниць апарату Гольджі, а також внутрішні мембрани цитоплазми — ендоплазматичного ретикулуму, мітохондрій і хлоропластів. Мембрани — високоорганізовані структури клітин, склад яких залежить від типу і функції мембрани, але завжди в них є ліпіди та білки.
У 1959 р. англійський вчений Д. Робертсон висунув гіпотезу щодо будови «елементарної» мембрани, в якій постулював структуру — спільну для всіх біологічних мембран. Згідно з цією гіпотезою всі мембрани мають вигляд тришарової структури, в якій центральний ліпідний подвійний шар розташований між двома шарами білка загальною товщиною 7,5 нм. У 1972 р. англійські вчені
С. Зінгер і Г. Ніколсон запропонували рідинно-мозаїчну модель мембрани, за якою молекули білків у ліпідах утворюють щось чиє до мозаїки.
Ліпіди в мембранах представлені фосфоліпідами, гліколіпідами та стеролами. Основу ліпідного шару складають фосфоліпіди, галактоліпіди, жирні кислоти, стерини. Ймовірно, ліпідний шар пронизують білкові молекули, які можуть бути зв'язані своїми гідрофобними бічними ланцюгами з внутрішніми гідрофобними частинами молекул ліпідів. Гранули внутрішніх ділянок мембран найчастіше є білками, які гідрофобно закріплені в ліпідному матриксі мембрани.
Біологічні мембрани розділяють цитоплазму на відсіки, в яких відбуваються певні біохімічні перетворення.
Лабільна структура мембран дає їм змогу, в свою чергу, виконувати найрізноманітніші функції: бар’єрну, осмотичну, транспортну, електричну, структурну, енергетичну, біосинтетичну, секреторну, рецепторно-регуляторну та інші.
7. Ендоплазматична сітка.
Ендоплазматичний ретикулум (сітка) — це складна тривимірна мембранна система, форма і протяжність якої визначається типом клітини та стадією її диференціювання. В трьох вимірах вона має пластинчасту будову і складається з багатьох мембранних мішечків — цистерн, на поверхні яких за допомогою електронного мікроскопа можна виявити численні гранули діаметром до 30 нм — рибосоми, де відбувається синтез білка. Через таку будову часто називають шорстким ендоплазматичним ретикулумом (ЕР).
Клітини, де відбувається інтенсивний синтез ліпідів, мають інший досить розгалужений гладенький трубчастий ендоплазматичний ретикулум. Обидва типи ЕР можуть бути одночасно навіть в одній клітині, причому між ними існує взаємозв'язок.
Функції шорсткого ЕР пов'язані із транспортуванням білків, синтезованих рибосомами на його поверхні. На початку синтезу білків початкову частину синтезованого поліпептидного ланцюга становить так звана сигнальна послідовність, яка за своєю конфігурацією відповідає специфічному рецептору на мембрані ЕР. Рецептор утворює канал, по якому білок переходить в цистерни ЕР для подальшого транспортування. В процесі перенесення білок зазнає істотних змін, наприклад, фосфорилюється, ацетилюється, перетворюється на глікопротеїни тощо.
Ендоплазматичний ретикулум має велике значення в біогенезі клітинних мембран, оскільки є основним місцем синтезу як мембранних білків, так і ліпідів клітин. В ЕР локалізовані кінцеві етапи синтезу мембранних ліпідів, зокрема гліколіпідів і фосфоліпідів. Останні забезпечують процес формування мембран мітохондрій і хлоропластів. Крім того, в ЕР відбувається біосинтез стероїдів, синтез усіх ненасичених кислот: саме тут синтезуються властиві лише рослинам жирні кислоти — лінолева, ліноленова, арахідонова. Похідними мембран ЕР є мембрани вакуолі, сферосом, мікротілець тощо. Ретикулум безпосередньо пов'язаний з ядерною оболонкою. Через мембранну систему АГ він бере участь в синтезі компонентів плазмалеми. Перехід мембран в різні види органел дістав назву потік мембрани. Існує уявлення про взаємодію мембранних компонентів клітин як концепцію ендомембранної системи, згідно з якою потік мембран пояснює функціональну неперервність мембран і участь їх у життєдіяльності всіх органел клітини.
Ендоплазматичний ретикулум є екстенсивною системою мембран, яка поділяє вміст еукаріотичної клітини на компартаменти і канали. Він формує одне ціле із зовнішньою мембраною ядерної оболонки. Шорсткий ЕР забезпечує синтез і транспортування білків крізь мембрану, тоді як гладенький ЕР відповідальний за синтез ліпідів та інші біосинтези.
8. Рибосоми, їх хімічний склад та функції.
Рибосоми — це дрібні сферичні частинки діаметром 0,2 мкм, які складаються в основному з високомолекулярної РНК (до 60 %) і білка. Рибосомальна (рРНК) синтезується в ядерці. Через самоскладання РНК разом із білком утворюють складну тривимірну структуру. Рибосома складається з малої (40S) і великої (60S) субодиниць. Мала містить одну 18S РНК і до 40 молекул білків, а велика субодиниця — по одній молекулі 5S РНК; 5,8S РНК; 25S РНК і до 45 молекул білків. В кожній клітині їх кілька десятків тисяч. Рибосоми можуть бути прикріплені до ЕР або вільно локалізовані в цитоплазмі. Є вони також і в ядрі, пластидах, мітохондріях. Тому розрізняють два типи рибосом: цитоплазматичні — з коефіцієнтом седиментації 80S (константу седиментації вимірюють в одиницях Сведберга, 1S=10-13 с) і рибосоми органел з коефіцієнтом 70S. Рибосоми досить часто утворюють комплекси — полірибосоми, які формуються поетапно з окремих субодиниць. Спочатку монтуються специфічні для кожної субодиниці білки за участю 28S і 18S рРНК. При цьому менша рибосомальна субодиниця взаємодіє з ініціаторною тРНК за наявності АТФ, ГТФ і білкових факторів ініціації. Цей комплекс з'єднується з мРНК у присутності йонів магнію, останньою до мРНК приєднується велика рибосомальна одиниця.
Рибосоми відповідають за синтез білка в клітині. Послідовність амінокислот у синтезованих на рибосомах білкових молекул визначається особливими молекулами матричної, або інформаційної РНК (мРНК), які переносять інформацію від геному ядра до рибосом. Мітохондрії та хлоропласти мають власний геном. Під час синтезу білка на рибосомах амінокислоти, з яких синтезується поліпептидний ланцюжок, приєднуються послідовно одна за одною. Саме в рибосомі молекули амінокислот можуть зайняти відносно одна одної цілком визначене місце. Таким чином, у синтезі білка беруть участь мРНК, яка несе генетичну інформацію, транспортна тРНК, яка постачає до рибосоми необхідні амінокислоти, і поліпептидний ланцюг, що нарощується. Необхідні також фактори, які відповідають за ініціацію, елонгацію та термінацію поліпептидного ланцюга. Під час синтезу білка в активному стані більшість рибосом прикріпляються до довгих ниткоподібних молекул мРНК, утворюючи в присутності іонів магнію полірибосоми. Це дає змогу одночасно синтезувати кілька десятків молекул одного й того самого білка.
9. Хлоропласти, їх будова та функція.
Пластиди - це продукти життєдіяльності рослинної клітини. Вони утворюються з пропластид - маленьких амебовидних тілець (d 0,05-0,5 мкм), які беруть початок від ініціальних часток, що відокремлюються від ядра і містять нуклеоплазму. У рослинних клітинах містяться три типи пластид: лейкопласти (безбарвні), хлоропласти (зелені) і хромопласти (оранжеві). Сукупність пластид прийнято позначати поняттям "пластидом".
Пластиди всіх трьох типів можуть взаємно перетворюватися. Наприклад, лейкопласти перетворюються у хлоропласти при позеленінні картоплі на світлі, у темряві хлоропласти втрачають зелене забарвлення і перетворюються в лейкопласти, при дегенерації хлоропластів і розпаді хлорофілу можуть утворюватись хромопласти.
Хлоропласти мають зелене забарвлення, що зумовлюється наявністю в них хлорофілу. Вони здатні самовідтворюватись: доведена їх висока автономність, яка полягає у здійсненні біосинтезу ряду білкових і ліпідних компонентів. Специфічна ДНК пластид відрізняється від ДНК ядра. Експериментально підтверджено її синтез безпосередньо у пластидах. У хлоропласті міститься близько 5% усієї клітинної ДНК. Вміст її змінюється залежно від фізіологічних умов. Число, розміри, форма і розміщення ділянок локалізації ДНК та їхня морфологія відрізняються у хлоропластів різних видів рослин.
Важливе значення у розвитку хлоропластів належить світлу, що необхідне для синтезу хлорофілів. Молекули хлорофілів локалізуються у внутрішніх мембранах, які формують два типи намел, занурених у гідрофільний білковий матрикс або строму. Одні з них тягнуться вздовж всієї пластиди - ламели строми, а інші - коротші, розташовуються одна над одною, утворюючи грани. За відсутності світла замість ламел формується проламелярне тіло — впорядкований центр пухирців і каналів. Проламелярними їх назвали для того, щоб підкреслити, що вони — попередники ламел. Після стимуляції світлом структури проламелярних тіл змінюють свою орієнтацію і швидко трансформуються в систему ламелярних мембран.
Пластиди, які містять проламелярні тіла, називають етіопластами. їх можна розглядати як певну стадію розвитку хлоропластів. Етіопласти утворюються в первинних листках або сім'ядолях паростків до того, як вони вийдуть із ґрунту на світло. У квіткових рослин хлоропласти із пропластид розвиваються лише на світлі, тоді як у деяких голонасінних це перетворення відбувається і за його відсутності.
Хлоропласти трапляються переважно в клітинах паренхіми листка, тоді як у меристемі вони відсутні. У клітині може бути від одного до сотень хлоропластів діаметром 5...8 мкм і товщиною до 1 мкм.
Кожен хлоропласт оточений подвійною мембраною з вибірковою проникною здатністю. Основна структурна одиниця внутрішньої мембрани хлоропластів — тилакоїд, тонкий плоский диск, оточений одношаровою мембраною.
У його мембрані містяться хлорофіл а і в, каротиноїди та білки, які беруть участь у фотосинтетичних реакціях. Існують два типи тилакоїдів — великий тилакоїд строми, який за довжиною наближається до розмірів самого хлоропласту, і менший тилакоїд, або тилакоїд грани, діаметр якого 30...60 нм. Хлоропласт може мати 40...60 гран і, як правило, — від 5 до 20 тилакоїдів у грані. В гранах виявлено перфорації, крізь які мембрани гран поєднуються, а отже і їхній внутрішньотилакоїдний простір сполучається за допомогою вузеньких трубочок — фрет.
Тилакоїдна система — це єдиний компартмент, відокремлений від строми тилакоїдною мембранною системою. Розрізняють три важливі компартменти хлоропластів:
· міжмембранний простір між зовнішньою та внутрішньою мембранами, які оточують хлоропласт;
· стромальний;
· внутрішньотилакоїдний.
Зовнішня мембрана проникна для метаболітів. Цю проникність забезпечують специфічні мембранні білки — поріни, які формують пори в мембрані, крізь які вільно проходять речовини з молярною масою до 10 кДа. Встановлено, що розміри відкритих пор, сформованих порінами у хлоропластів, досягають 3 нм.
Стромальний компартмент забезпечує важливі метаболітичні цикли фотосинтезу, а внутрішньотилакоїдний — формує протонний градієнт в ході світлових реакцій фотосинтезу.
10. Мітохондрії, їх будова та функції.
Мітохондрії містяться у цитоплазмі всіх клітин еукаріотів. У процесі статевого розмноження промітохондрії передаються нащадкам через яйцеклітину. Розмноження мітохондрій відбувається шляхом поперечного поділу, а також брунькуванням. Утворені дочірні органели промітохондрії - з часом перетворюються у зрілі мітохондрії.
До складу мітохондрій входять білки, ліпіди, вітаміни, рибосоми, РНК, ДНК, ферментативний комплекс.
Основною функцією мітохондрій є забезпечення процесів життєдіяльності клітини необхідною енергією шляхом перетворення енергії хімічних зв'язків при окисленні дихального субстрату в енергію макроергічних зв'язків аденозинтрифосфорної кислоти (АТФ). Утворення АТФ відбувається при ферментативному розщепленні вуглеводів, жирних кислот, амінокислот у процесі окислювального фосфорилювання. Крім того, у мітохондріях проходить біосинтез ліпідів і білків, які беруть участь у загальному процесі транспорту іонів у клітині.
Тривалість життя мітохондрій не перевищує кількох діб. Постійним новоутворенням і розпадом органел забезпечується велика різноманітність їх форм в одній клітині. Довжина мітохондрій коливається від одного до кількох мкм, а ширина становить близько 0,5 мкм. Інколи в особливо активних клітинах розміри мітохондрій більші. Кількість їх у різних клітинах може коливатися у досить широких межах і досягати 2000 штук.
Мітохондрія утворена подвійними мембранами - зовнішньою і внутрішньою. Зовнішня мембрана гладенька, пориста, містить ферменти і білки, здатна пропускати речовини з невеликою молекулярною масою, а також іони. Вона є легкопроникною для пірувату, що утворився при неповному окисленні шестивуглецевих сполук у цитоплазмі.
Внутрішня мембрана має досить складну будову, утворює численні складки різної форми, які називаються кристами. Поверхня цієї мембрани густо вкрита грибоподібними (елементарними) частками, у яких при сутні фактори, що забезпечують синтез ДНК. Внутрішня мембрана проникна тільки для води і невеликих нейтральних молекул. У ній локалізований дихальний ланцюг, що складається з переносників електронів і невеликої кількості білків.
Завдяки кристам внутрішній вміст мітохондрій - матрикс - поділений на відсіки. Мітохондріальний матрикс є гелеподібною речовиною, що містить до 50% білків. У ньому локалізовані ферменти циклу три-карбонових кислот (циклу Кребса) і синтезу ліпідів та білків. У матриксі також розміщені рибосоми і мітохондріальна ДНК.
Незважаючи на наявність власної генетичної і білоксинтезуючої системи, мітохондрії не повністю автономні. Значна частина інформації про їх структуру і функції міститься у хромосомах ядра.