БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
9. АЛЛЕЛОФИЗИОЛОГИЯ
Растения не только реагируют на физические или химические раздражители из своего неживого (абиотического) окружения (см. 7.7), но и вступают также в многообразные взаимодействия с другими живыми организмами. В качестве примера уже были упомянуты управляемые фитохромом реакции на затенение или соответственно на отраженный соседними растениями свет (см. 7.7.2.1). Исследование молекулярных процессов, происходящих при взаимодействии растений с другими организмами, представляет собой сегодня самостоятельную область физиологии, которая обобщенно представлена здесь под названием аллелофизиология (греч. — allelos, взаимный, обоюдный).
Аллелофизиология связана с экологией растений (см. 13.8) и, кроме того, с фитопатологией, что в рамках этой книги, однако, можно осветить лишь в общих чертах. Разнообразные связи растений с их опылителями упоминаются в разделе 11.2 при рассмотрении соответствующих таксонов. Поскольку растительные движения органов цветка связаны с опылителями, об этом говорилось в разделе, посвященном физиологии движения (см. 8.3.2).
Теснейшее взаимодействие организмов имеет место при симбиозе (см. 9.2). Под этим понимают тесное сосуществование двух различных видов организмов, из которого оба извлекают по меньшей мере временную пользу. Тем самым симбиоз отличается от комменсализма (польза для партнера без заметного влияния на другого) и паразитизма (см. 9.1.1, польза для одного через угнетение другого). Симбиотическое сосуществование позволяет также ясно увидеть, что оно возникло из взаимного паразитизма (аллелопаразитизм), при котором наступило равновесие между партнерами в поражении и защите, и теперь они взаимно отбирают питательные вещества и гормоны. Если один из партнеров в ходе симбиоза становится доминирующим, это равновесие может исчезнуть и снова перейти в паразитизм, как при переваривании клубеньковых бактерий их клетками-хозяевами (см. 9.2.1).
Между паразитами и патогенами также нет четкой границы. Возбудителями болезней обычно называют микробных паразитов, которые так вредят организму-хозяину, что появляются характерные симптомы поражения и паразиты при этом (часто значительно) размножаются (патогены, см. 9.3). Повреждение может привести к отмиранию хозяина или определенной его ткани. Если патоген берет питательные вещества из отмерших участков для себя, значит, он питается сапрофитно (см. 9.1.1).
К гетеротрофам относятся также животные, поедающие растения (травоядные, см. 9.4), которые покрывают свою потребность в органическом веществе исключительно или преимущественно за счет автотрофных растений — первичных продуцентов, стоящих в начале пищевой цепи.
Для всех названных взаимодействий можно установить степень специфичности — иногда очень высокую — в отношении взаимодействующих организмов. Растения устойчивы к большинству потенциальных возбудителей и лишь к небольшому числу восприимчивы; они защищены от большинства травоядных и атакуются лишь небольшим их числом; паразитические отношения и симбиозы образуются, как правило, также лишь между определенными партнерами. Основой специфичности к хозяину можно считать «процессы узнавания», в ходе которых между участвующими организмами часто происходит взаимный обмен сигнальными молекулами (от них, например, зависит, устойчиво или восприимчиво растение к патогену). Химическое взаимодействие между растениями, как между индивидуумами того же или (чаще) различных видов, называется аллелопатией (см. 9.5). В этом случае речь часто идет об ингибиторах, которые производятся одним индивидуумом и, выделяясь в среду, оказывают отрицательное влияние на рост растений-конкурентов.
9.1. Особенности гетеротрофного питания
Взаимодействие растений с другими организмами связано прямо или косвенно с питанием. Аллелопатия лежит в основе внутри- или межвидовой конкуренции среди автотрофных растений за ограниченное количество питательных веществ; при всех остальных взаимодействиях речь идет об аспектах гетеротрофного питания или самого растения и/или вступающего с ним в отношения другого организма.
В отличие от автотрофных организмов, усваивающих неорганические питательные вещества (см. 6.4 —6.9), гетеротрофы питаются органическими веществами. Если в основном автотрофный организм нуждается для роста в отдельных простых органических соединениях, говорят о миксотрофии, или прототрофии. Мутантов, потерявших способность к образованию отдельного, требующегося для роста органического вещества (например, аминокислоты, кофактора), обозначают как ауксотрофы: они должны получать такое вещество извне.
Среди гетеротрофов различают сапрофитов, берущих органическое питание из мертвого субстрата, и паразитов, которые используют живые организмы или клетки.
Сапрофитами является большинство бактерий и грибов, и, напротив, их нет среди высших растений. Требования к питательному субстрату, в частности, у сапрофитов очень различны. Наряду с неорганическими веществами им необходим источник углерода, в качестве которого могут служить не только углеводы, жиры или белки, но и спирты, органические кислоты и т.п., а также нефть, парафин, бензол и нафталин. Часто сапрофиты выделяют экзоэнзимы, которые внеклеточно разлагают высокомолекулярные субстраты (например, лигнин, целлюлозу, белки) до пригодных для поглощения продуктов. Усвоенный органический материал включается затем в нормальный (катаболический или анаболический) обмен веществ. Многие сапрофиты не нуждаются в органически связанном азоте. Так, например, дрожжи могут расти на NH+4, плесневый гриб Aspergillus niger — на NO-3 в качестве единственного источника азота (см. 6.6).
В природе чаще кооперируются целые группы различных организмов, при этом один вид усваивает продукты разложения или отходы другого и питается ими, в то время как его выделения могут в свою очередь служить другим видам в качестве питательного субстрата, частично также в качестве «сырья» для реакций энергообеспечения при хемосинтезе (Н2S, Н2, NH3). Такого рода процессы происходят, например, при гниении, когда органический материал, в частности, из отмерших растений, частей растений или животных, бактерии и грибы снова переводят в неорганические соединения (реминерализируют); гниение поэтому и является важным звеном круговорота веществ. На подобных процессах основано «биологическое самоочищение» загрязнений воды. При технической очистке сточных вод с помощью активного ила сообщества сапрофитов используются для переработки органических отходов. Похожие процессы минерализации протекают также в почве (например, при приготовлении компоста). Все названные процессы в совокупности имеют огромное значение для баланса обмена веществ на Земле.
Продукты микробиологического распада служат исходными веществами для образования гумуса, угля и нефти, но в этих процессах в дальнейшем играют решающую роль абиотические химические превращения, частично под высоким давлением (уголь и нефть).
Паразиты встречаются среди бактерий, грибов, лишайников и семенных растений. Некоторые гетеротрофные красные водоросли паразитируют на близкородственных представителях багрянок (адельфопаразитизм). Организмы, которые питаются в природе как сапрофитно, так и паразитически, называются факультативными паразитами, тех, кто от природы постоянно нуждается в живых организмах в качестве хозяев, — облигатными паразитами. Но в эксперименте и облигатные паразиты часто могут жить сапрофитно на подходящей искусственной питательной среде.
Микробиологические паразиты (бактерии, грибы) являются причиной многих заболеваний у растений, животных и человека, они являются возбудителями болезней (патогенами). Микробиологические патогены из-за большой сложности их взаимодействий с растениями представлены отдельно (см. 9.3).
Среди голосеменных известен лишь один паразитический вид: представитель семейства подокарповые Parasitaxus ustus, как и его хозяин Falcatifolium taxoides из того же семейства; этот эндемичный вид обитает в Новой Каледонии. Паразит контактирует с ксилемой хозяина и получает таким образом воду и питательные соли.
У паразитических покрытосемепных, которые всегда являются облигатными паразитами, различают геми- и голопаразитов (полу- и полных паразитов). Полу паразиты (например, большинство омел и норичниковые Rhinanthus, Melampyrum, Pedicularis, Euphrasia) способны к фотосинтезу, усваивают неорганические питательные вещества и воду, но не корнями из почвы, а с помощью гаусторий из ксилемы хозяина. Но они, как правило, растут лишь на специфических хозяевах (различные формы Viscum album, например, на елях, соснах и лиственных деревьях), поэтому для них важны, по-видимому, также органические вещества, которые присутствуют в незначительной концентрации и в ксилеме (см. 6.3.5; 6.8). Эти гемипаразиты должны «перевести» содержимое проводящей системы хозяина в свое вегетативное тело против тургора, поэтому они имеют, как правило, особо интенсивную транспирацию на единицу поверхности листа (отсюда, например, быстрое увядание сорванного Melampyruml). Примечательно, что растения, которые во время определенных стадий онтогенеза (например, Tozzia и Bartsia) или на протяжении всей жизни (Lathraea) лишены развитых транспирирующих листьев, образуют на своих корневищных чешуях гидатоды, которые выделяют воду и таким образом поддерживают необходимую разность водного потенциала между хозяином и паразитом. Замыкающее звено этого ряда у подсемейства погремковые семейства норичниковые, паразитирующих на ксилеме, представляет Lathraea — растение с чешуйчатым корневищем, которое паразитирует на многолетних хозяевах и, очевидно, получает достаточно органического материала из ксилемы хозяина, чтобы жить как голопаразит.
И у омел известен полностью паразитический вид — безлистный Tnstenx aphyllus (Loranthaceae), который паразитирует на видах кактусовых. Еще не выяснено, присоединяется ли он к ксилеме или флоэме хозяина. Другие полностью паразитические покрытосеменные, например, Singa, Огоbranches Cuscuta (см рис 11 281), присоединяются к ситовидным трубкам хозяев, из которых они с использованием особенных поглощающих клеток (клеток переноса) не выясненным образом забирают ассимиляты.
Высшие растения часто служат хозяевами для паразитических животных (зоопаразитов). На побегах чаще паразитируют членистоногие, а на корнях нематоды (круглые черви) Нематоды ежегодно во всем мире причиняют значительный вред сельскому хозяйству. Состояния пораженных паразитами растений, связанные с проявлением характерных симптомов болезни, называются паразитозами. К паразитическим членистоногим относят, например, молей-пестрянок, чьи личинки питаются в листьях растений-хозяев тканями мезофилла Чаще, однако, присутствие паразитов связано с образованием растительного галла (цецидии). Вообще, так обозначается любое вызванное чужеродным, паразитическим организмом активное нарушение тканей и органов ограниченным ростом. Если поврежденные ткани растут неограниченно, их называют опухолями (см 9 3 3, 7 6 2 3, бокс 9 2). Симбиотические структуры (например, корневые клубеньки, см 9 2 1) сегодня больше не относят к галлам, однако существуют галловые образования, вызванные паразитическими бактериями или грибами (например, «ведьмипы метлы» развиваются в результате поражения Rhodococcus fascians, см 7 6 2 3 и грибами рода Taphrina, см 112, рис 11 29).
Органоидные галлы состоят из сильно измененных, но все же еще четко узнаваемых основных органов растения-хозяина (например, «ведьмины метлы»). Более распространенные гистоидные галлы (рис 9 1) не позволяют различить органное строение, а возникают как образования из частей стебля, листа или корня и вызываются, как правило, галлообразующими животными в области побега — в первую очередь галлипами, орехотворками, галлообразующими вшами и клещами, в области корня — цистообразующими нематодами родов Heterodera и Globodera или соответственно галловыми нематодами родов Мeloidogyne (рис 9 2). Паразит при этом проникает в растительную ткань, поэтому галлообразующие животные называются эндопаразитами. Эктопаразиты, напротив, не полностью проникают в растение-хозяина, а паразитируют на его поверхности (например, сосущие насекомые тли, растительноядные клопы и цикады, некоторые нематоды).
. Рис. 9.1. Различные гистоидные галлы на листе Fagus sylvatica Специфическая форма галлов обусловлена действием животных: 1 — кармашковидный галл, вызванный галли- цей буковой волосистой Mikola fagi (рис начальная страница часть 4) 2 — опушенный кармашковидный галл галлицы Hartigiola annuhpes, 3 — войлочный галл на жилках листа вызванный клещом Eriophyes nervisequus
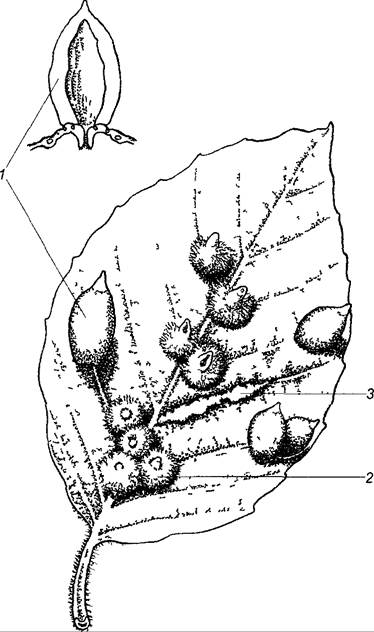
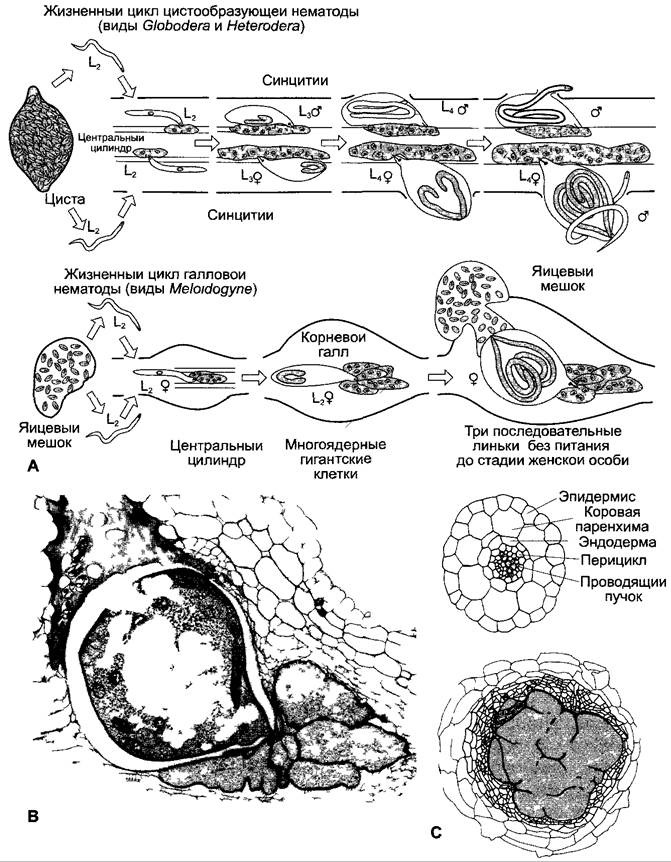
Рис. 9.2. Галлообразующие, эндопаразитические нематоды: А — жизненные циклы цистообразующих и галловых нематод В — взрослая женская особь галло вой нематоды (Meloidogyne incognita) в галле корня огурца С — поперечный срез корня Arabidopsis thahana в непораженном состоянии (сверху) а также (ниже) поперечный срез через синцитиальную область пораженного корня (синцитий внизу, цистообразующая нематода Heterodera schachtu жен ская личинка в четвертой личиночной стадии) L — личиночные стадии
Гистоидные галлы приспособлены прежде всего к потребностям галлообразующих животных. Эти приспособления часто необычны и сложны. Кармашковидный галл, например, часто возникает на листьях бука (см. рис. 9.1) из-за локального роста поверхности, вызванного личинками галлицы буковой волосистой. Личинки «лепят» себе галловый футляр с помощью своей слюны. В конце концов смоченные слюной области выгибаются в свод в виде кармашка, так что возбудители оказываются полностью закрытыми в кармашках, имеющих внизу выводной проток. У многих галлов также происходит последующее утолщение и образование склеренхимных элементов. Возникает стойкий футляр для защиты развивающегося животного. Обильные волосистые образования и тонкостенные, богатые питательными веществами клетки внутри футляра обычно служат для питания галлообразующих животных.
В названных примерах после этого под влиянием чужеродного организма образуются такие клеточные и органные формы, для которых, возможно, имеется генетический потенциал в растении, но они не образуются в обычных условиях. Нет сомнений, что различные галлы появляются благодаря специфическому1 влиянию галлообразующих организмов. При этом важную роль, по-видимому, играют фитогормоны.
1 Эта специфика определяется временными и пространственными факторами развития тканей растения. — Примеч. ред.
В связи с потребностями сельского хозяйства в последнее время были подробно исследованы реакции растений, вызываемые эндопаразитическими нематодами (рис. 9.2). Во второй личиночной стадии (L2) нематоды заражают корни растений вблизи кончика корня. Цистообразующие нематоды проникают своим стилетом в клетки прокамбия, которые — из-за секретов слюнных желез животных — сначала сильно вздуваются. Затем благодаря частичному растворению клеточных стенок и слиянию протопластов образуются большие по объему, величиной более 200 клеток, и обладающие высокой активностью обмена веществ синцитии («цисты»), из которых теперь уже неподвижный паразит высасывает питательные вещества (рис. 9.2, А, С). Хорошо обеспеченные питанием животные развиваются в женские особи, которые при отмирании несут многочисленные уже содержащие эмбрионы яйца, способные сохраняться в почве несколько лет. Корневые галловые нематоды вызывают резкие изменения прокамбиальных клеток в кончике корня. Образуются не синцитии, а многоядерные (благодаря эндомитозам) гигантские клетки с высокой активностью обмена веществ (рис. 9.2, А, В), содержащие до 100 крупных клеточных ядер. Функционируя как сильные акцепторы (см. 6.8.3), такие клеточные структуры вызывают значительный отток питательных веществ из продуктивных органов растения. Паразит получает питательные вещества по симпластическому пути из гигантских клеток. Общеизвестно, что реакции растения вызываются продуктами обмена веществ животного. Природа растворяющих веществ нематод еще совсем не изучена. Зато были найдены виды или сорта, например, внутри родов Beta и Solanum, отличающиеся значительной устойчивостью к поражению нематодами. Клонирование генов (см. бокс 7.3), обусловливающих устойчивость (гены устойчивости, R-гены), дало неожиданный результат: часть этих генов очень сходна с генами, влияющими на устойчивость к патогенным бактериям или вирусам (см. 9.3.4). Растения обладают механизмами устойчивости, направленными против широкого спектра вредителей (см. 9.3.4; 9.4.1).
Растения, способные ловить животных (плотоядные — см. бокс 4.4), за исключением грибов, образующих ловушки (см. 11.2; рис 11.62), всегда обладают хлорофиллом, способны к С3-фотосинтезу и позволяют легко культивировать их при достаточном минерально-солевом питании без животной пищи. Лишь при недостатке питательных веществ в их естественной среде обитания, как это часто бывает (например, в верховых болотах), они отлавливают животных прежде всего для обеспечения себя азотом и фосфором. У Utricularia exoletaобразование цветков значительно стимулируется животным питанием.
Приспособления хищных растений позволяют привлечь определенных животных и удержать их ловчими структурами (ловчие механизмы, см. бокс 4.4, а также раздел
8.3.2.4). Переваривание происходит при участии экзоферментов, прежде всего протеаз, которые выделяются специальными железками после раздражения жертвой (например, у Drosera) либо независимо от этого (например, пепсиноподобная протеаза с оптимумом pH в сильно кислой области в кувшинах Nepenthes', см. бокс 4.4, рис. А). У Sarraceniaпищеварительные ферменты должны выделяться бактериями в ловчую жидкость. Продукты переваривания всасываются растением часто с помощью абсорбционных волосков и включаются в обмен веществ.