БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
6. ФИЗИОЛОГИЯ ОБМЕНА ВЕЩЕСТВ
6.3. Водный обмен
Вода в живой клетке является не только универсальным растворителем, но и субстратом в процессах клеточного обмена веществ, например, донором электронов и протонов при фотосинтезе (см. 1.1). Вода как основная составная часть живой клетки несет также структурную функцию, и в процессе роста растения большая часть
увеличения его размеров обеспечивается за счет воды («вода роста»). Поскольку растения для фотосинтеза должны поглощать из атмосферы газообразный СО2, а в процессе эволюции так и не было найдено покровное вещество, которое пропускало бы только СО2, но не воду, то растение вынуждено постоянно путем испарения (транспирации) терять воду (см. 6.3.4.1) — эта «транспирационная вода» должна восполняться во избежание потери тургора. У наземных растений испарение воды дополнительно обеспечивает определенное охлаждение организма, однако эта функция не является жизненно необходимой. Об этом свидетельствуют, в частности, особенности многих растений жарких, аридных областей, так называемых САМ-растений (см. 6.5.9): на протяжении всего дня устьица у них закрыты, что сильно сокращает транспирацию. Однако в любом случае при высоких температурах растения уменьшают, а не увеличивают транспирацию. Поглощение воды, как правило, осуществляется теми же органами, которые обеспечивают поглощение питательных минеральных веществ, и энергетически связано с ним, поэтому водный обмен обычно рассматривают вместе с минеральным питанием растений. При этом внимание исследователей сосредоточено в основном на наземных высших растениях (мхах и семенных растениях). Для них регулируемый водный обмен имеет особое значение, поскольку они живут в местообитаниях, которые обычно характеризуются недостатком воды (сравнительно сухие почвы, сухой воздух).
Растения не могут транспортировать воду активно (см. 6.1.4.2). Она постоянно пассивно передвигается как на клеточные, так и на макроскопические расстояния по градиенту ее химического потенциала, т. е. из участка с более положительным водным потенциалом к участку с более отрицательным водным потенциалом (уравнение 6.10, 6.15), причем свободная энтальпия воды снижается. Следовательно, транспорт воды — это экзергоничный процесс, и протекает он самопроизвольно.
Непрерывный путь воды из почвы через корень, а затем через проводящую систему ксилемы, ведущую в ткани и, наконец, к местам испарения воды, а также сами процессы водоотдачи можно рассматривать в плане энергетики на основе концепции водного потенциала; однако на разных этапах передвижения воды движущие силы и транспортные механизмы различаются. При этом для большей ясности следует разделить этапы поглощения воды, ее передвижения и испарения. Рассмотрим прежде всего транспортные механизмы и водный обмен клетки.
В основе движения воды лежат два основных механизма: диффузия и массовый ток.
Под диффузией понимают пассивное перемешивание частиц, возникающее вследствие теплового движения, неупорядоченное относительно отдельной частицы. Посредством диффузии осуществляется (при наблюдении за многими частицами) нетто-течение вещества в определенном направлении в том случае, если в диффузионном пространстве имеются различия химического потенциала этого вещества. В смешанной фазе (например, в растворителе с растворенными в нем веществами) все компоненты, для которых существует градиент химического потенциала, в том числе и растворитель, находятся в нетто-течении до тех пор, пока не выравниваются различия химического потенциала (см. 1.4). В большинстве случаев концентрационный градиент является причиной различия химического потенциала, которое служит движущей силой диффузионного процесса.
Термином «поток» («скорость диффузии», Ji) обозначается диффундирующая масса вещества i, рассчитанная на единицу площади и временной интервал. Зависимость потока от концентрационного градиента 1 на отрезке х (∆сi/∆х) перпендикулярно площади поперечного сечения описывается первым законом диффузии Фикка:
![]()
и может выражаться в [моль м-2 с-1]. Таким образом, скорость диффузии прямо пропорциональна концентрационному градиенту диффундирующего вещества. Фактор пропорциональности Б называется коэффициентом диффузии (м2 с-1). В условиях постоянной температуры и постоянного давления D является специфичным для данного вещества и помимо того зависит от диффузионной среды. Газы, в частности СO2 и O2, диффундируют в воздухе приблизительно в 105 раз быстрее, чем в воде (СO2 в воздухе: 1 см с-1, в воде: 10 5 см • с1). Отрицательный знак в уравнении указывает, что положительный поток вещества наблюдается в направлении понижающегося концентрационного градиента (от более высокой к более низкой концентрации вещества).
Скорость движения молекул увеличивается с повышением температуры, отсюда скорость диффузии пропорциональна температуре (и равна нулю в точке абсолютного нуля, 0 К).
Второй закон Финка определяет зависимость расстояния (х), пройденного диффундирующим веществом за данный промежуток времени (t):
![]()
Таким образом, расстояние, пройденное в процессе диффузии, пропорционально корню квадратному из времени. Фактор пропорциональности к имеет размерность [м • с-1/2]. Итак, диффузия — это транспортный механизм, эффективный лишь при очень малых перемещениях, не пригодный для преодоления больших расстояний.
Это подтверждают некоторые цифры. Вот как диффундирует краситель флуоресцеин в воде (при одной и той же данной температуре и одном и том же данном концентрационном градиенте): за секунду — 87 мкм, за минуту — приблизительно 675 мкм, за час — приблизительно 5 мм и за год — приблизительно лишь 50 см. Следовательно, в пределах растительных клеток диффузия вполне эффективна. Однако при преобладающих концентрационных градиентах и прочих условиях молекула сахара, например, образованная в листе древесной кроны на высоте 30 м, за счет одной диффузии никогда не достигла бы корня за время жизни дерева, так же как поглощенный корнем питательный элемент ни за что не попал бы в лист таким образом.
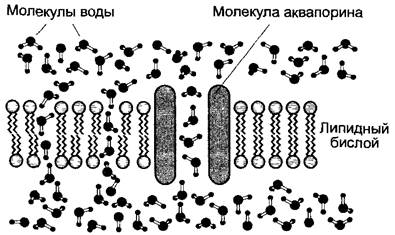
Рис. 6.25. Диффузия молекул воды через биомембраны и каналы, образованные водоспецифичными белками аквапоринами
Диффузия как механизм передвижения воды имеет значение:
✵ частично для транспорта воды между почвенным раствором и апопластом;
✵ транспорта воды между апопластом и симпластом;
✵ транспорта воды через клеточные мембраны;
✵ перемещения воды из апопласта в воздух межклетников, например, в листьях;
✵ перемещения воды из воздуха межклетников в атмосферу.
Быстрая диффузия воды через клеточные мембраны долгое время оставалась непонятной. В настоящее время считается, что незначительное сопротивление диффузии воды со стороны клеточных мембран (вода диффундирует через клеточную мембрану почти так же хорошо, как через слой воды равной толщины) объясняется тем, что молекулы воды диффундируют, во- первых, через «отверстия» в жидком липидном бислое, а, во-вторых, также и через каналы, образованные водоспецифичными белками аквапоринами,расположенными в клеточной мембране (рис. 6.25). Путем регуляции количества аквапоринов и их фосфорилирования клетка может регулировать проницаемость своих мембран для воды в зависимости от изменяющихся потребностей.
Для дальнего транспорта воды диффузия непригодна. Здесь имеют место массовые токи воды. Они характерны
✵ для транспорта воды в сосудах ксилемы,
✵ транспорта воды в почве,
✵ по обстоятельствам — для транспорта воды в апопластах, например, в листе и корне,
✵ массовый ток лежит также и в основе флоэмного транспорта
В то время как движущей силой диффузии считается разность концентрационных потенциалов воды, массовый ток вызывается преимущественно разностью потенциалов давления между двумя участками (см 6 3 5,6 8 3). Таким образом, в образование химического потенциала (соответственно водного потенциала) вносят различный вклад составные потенциалы, что создает движущую силу для транспорта воды путем диффузии или массового тока.
Зависимость массового тока от разности потенциалов давления описывается законом Гагена—Пуазейля, который строго соблюдается лишь для идеальных капилляров
![]()
Следовательно, объемный поток ∆ \// ∆ t (он выражается, например, в [м3 с 1]) при постоянном радиусе капилляра г и для жидкости с постоянной вязкостью ц прямо пропорционален приложенной разности давлений ∆р/ ∆х, в свою очередь, при данных ∆р/ ∆х и ɳ он очень сильно зависит от радиуса капилляра при удвоении радиуса объемный поток на единицу времени возрастает шестнадцатикратно (24 = 16' Отрицательный знак показывает, что положительный поток движется в направлении убывающего гидростатического давления (∆р/∆х < 0)
Обратите внимание р идентичен потенциалу давления из уравнения водного потенциала (см уравнение 6 15), но я в уравнении 6 32 — это число «пи», выводимое из геометрии капилляра с круговым сечением, и его нельзя путать с осмотическим давлением П в уравне нии водного потенциала.
6.3.2. Водный обмен клетки
6.3.2.1. Осмос
Поглощение и соответственно испарение воды клетками осуществляются преимущественно осмотическим путем, а механизмом транспорта является диффузия. Под осмосом понимают диффузию частиц через избирательно проницаемую мембрану, каковую представляет собой биологическая мембрана (см 2 2 5). Эта мембрана хорошо проницаема для растворителя (воды), но для растворенных в нем веществ полностью (в идеальном случае) или лишь слабо проницаема. Если избирательно проницаемая мембрана разделяет две жидкости с различными концентрациями растворенных частиц, то в результате по обе стороны мембраны возникает также разность водных потенциалов (![]() , причем
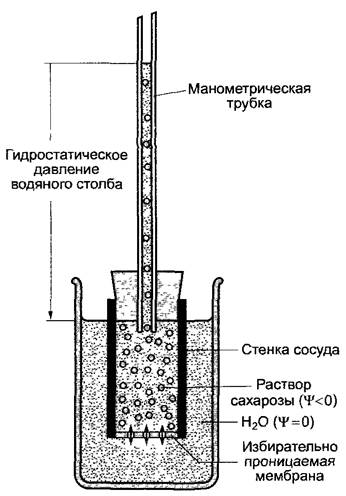
, причем![]() более отрицателен на стороне более высокой концентрации частиц). Молекулы воды диффундируют по своему концентрационному градиенту из более разбавленного в менее разбавленный раствор, что можно экспериментально показать в опыте с осмометром (ячейкой Пфеффера, рис 6 26). Вследствие этого (в результате разбавления) объем концентрированного раствора увеличивается, и развивается гидростатическое давление. Поглощение воды компартментом с отрицательным водным потенциалом продолжается до тех пор, пока гидростатическое давление, появившееся вследствие тока воды, не компенсирует разность потенциалов воды между обоими компартментами
более отрицателен на стороне более высокой концентрации частиц). Молекулы воды диффундируют по своему концентрационному градиенту из более разбавленного в менее разбавленный раствор, что можно экспериментально показать в опыте с осмометром (ячейкой Пфеффера, рис 6 26). Вследствие этого (в результате разбавления) объем концентрированного раствора увеличивается, и развивается гидростатическое давление. Поглощение воды компартментом с отрицательным водным потенциалом продолжается до тех пор, пока гидростатическое давление, появившееся вследствие тока воды, не компенсирует разность потенциалов воды между обоими компартментами![]() .
.
Рис. 6.26. Схема осмометра (ячейка Пфеффера)
Живая клетка является таким же осмометром Избирательно проницаемой мембраной служат плазмалемма и (поскольку осмотики накапливаются главным образом в вакуолярном соке) тонопласт При достаточной доступности воды клетка осмотическим путем поглощает воду до тех пор, пока не достигнет состояния![]() Мало
Мало
эластичные клеточные стенки допускают увеличение объема лишь в узких границах, поэтому вследствие осмотического поступления воды в клетке быстро возникает гидростатическое давление, называемое также тургором, или тургорным давлением. Дополнительный вклад в создание потенциала давления в структуре ткани вносится окружающими тургесцентными клетками, которые оказывают противодействие увеличению поглощающих воду клеток.
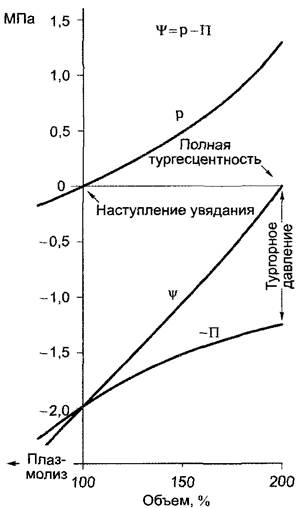
На клеточном уровне гравитационным потенциалом пренебрегают, и тогда водный потенциал клетки (или ткани) можно выразить в виде уравнения 6.15 (см. 6.1.4.2):
где р — гидростатическое давление, тургор; -П — осмотический потенциал.
Водный потенциал клетки (ткани)![]() колеблется при этом в пределах от
колеблется при этом в пределах от![]() (если р = П) до
(если р = П) до![]() = -П (если р = 0). Потенциал воды приобретает нулевое значение, если тургорное давление клетки полностью компенсируется осмотическим потенциалом (полная тургесцентность), при отсутствии тургора (р = 0, состояние увядания) клетка (ткань) развивает максимально отрицательный водный потенциал, величина которого определяется через П, отражая суммарную концентрацию всех осмотиков в клетке (ткани). Соотношения
= -П (если р = 0). Потенциал воды приобретает нулевое значение, если тургорное давление клетки полностью компенсируется осмотическим потенциалом (полная тургесцентность), при отсутствии тургора (р = 0, состояние увядания) клетка (ткань) развивает максимально отрицательный водный потенциал, величина которого определяется через П, отражая суммарную концентрацию всех осмотиков в клетке (ткани). Соотношения
между![]() , р, П и объемом клетки представлены на рис. 6.27.
, р, П и объемом клетки представлены на рис. 6.27.
Рис. 6.27. Изменение показателей состояния при осмотических процессах поглощения и потери воды клеткой
Клеточными осмотиками наряду с органическими соединениями (такими, как сахара и органические кислоты) служат в еще большей степени неорганические соли, накапливающиеся в цитоплазме, но преимущественно в вакуоли, т.е. в клеточном соке (см. 6.2.3). Особенно значимыми в количественном отношении осмотиками являются К+ и его противоионы (СI- и/или органические кислоты, как, например, малат). Общая концентрация осмотиков в клеточном соке составляет в большинстве случаев 0,2 — 0,8 М. В определенных клетках (например, в замыкающих клетках устьиц, см. 8.3.2.5) эта концентрация может претерпевать значительные и обратимые изменения. Раствор КС1 в концентрации 0,1 М при 25 °С обладает осмотическим потенциалом — 0,46 МПа.
Осмотический потенциал клетки можно определить в опытах с наблюдением плазмолиза. Под плазмолизом (см. рис. 2.60) подразумевают отхождение протопласта от клеточной стенки вследствие его сжатия в гипертонической среде (т.е. среде, для которой справедливо:![]() где С — среда, К — клетка). При этих условиях клетка теряет воду до тех пор, пока не уравняются
где С — среда, К — клетка). При этих условиях клетка теряет воду до тех пор, пока не уравняются![]() (вследствие тока воды сначала быстро понижается потенциал давления — тургор, а в конечном счете в результате продолжающейся потери воды возрастает П; в течение этого процесса
(вследствие тока воды сначала быстро понижается потенциал давления — тургор, а в конечном счете в результате продолжающейся потери воды возрастает П; в течение этого процесса![]() становится все более отрицательным, пока его значение не достигнет величины
становится все более отрицательным, пока его значение не достигнет величины![]() ). В гипотонической среде
). В гипотонической среде![]() процесс плазмолиза обратим, и тогда говорят о деплазмолизе. И в гипертонической среде со временем также наступает деплазмолиз, так как происходит медленное выравнивание концентраций внеклеточных осмотиков путем диффузии через клеточные мембраны, и клетки приводят концентрации эндогенных осмотиков в соответствие со средой.
процесс плазмолиза обратим, и тогда говорят о деплазмолизе. И в гипертонической среде со временем также наступает деплазмолиз, так как происходит медленное выравнивание концентраций внеклеточных осмотиков путем диффузии через клеточные мембраны, и клетки приводят концентрации эндогенных осмотиков в соответствие со средой.
Если теперь мы поместим клетку (или клетки в составе ткани) в растворы с различными концентрациями осмотиков и найдем тот (изотонический) раствор, в котором наблюдается пограничный плазмолиз (состояние, при котором вследствие выхода воды из клетки протопласт только начинает отходить от клеточной стенки) и соответственно тургор как раз достигает нулевого значения (р = 0), следовательно,
![]()
Но![]() равняется -Пс, так как у раствора в равновесии с окружающей средой не обнаруживается гидростатического давления, поэтому справедливо Пс = Пк.
равняется -Пс, так как у раствора в равновесии с окружающей средой не обнаруживается гидростатического давления, поэтому справедливо Пс = Пк.
Еще один метод определения осмотического потенциала клеточного сока — криоскопия (от греч. kryos — мороз) выжатых клеточных соков, т.е. определение снижения точки замерзания.
Осмотические потенциалы могут весьма значительно различаться не только у отдельных видов растений (рис. 6.28), но и внутри одного растения в различных органах и тканях. В паренхимных клетках коры корня значения осмотического потенциала лежат между -0,5 и -1,5 МПа; в побегах они, как правило, по мере удаления от корня становятся отрицательнее и достигают в клетках ткани листа значений от -3 до -4 МПа. Типичные значения для листьев бука: в нижнем эпидермисе -1,4 МПа; в губчатой паренхиме -2,1 МПа; в палисадной паренхиме -3,8 МПа. У растений, произрастающих в очень сухих местообитаниях, например, в пустыне, или на засоленных почвах, например, на морском берегу или в солончаковой пустыне, осмотические потенциалы клеточных соков могут достигать значений, более отрицательных, чем -10 МПа (Limoniumна засоленной почве < -16 МПа, Аtriplex < -20 МПа). Некоторые плесневые грибы способны расти даже на концентрированных растворах сахаров (например, на фруктовом желе) с осмотическими потенциалами субстратов до -22 МПа.
Рис. 6.28. Колебания осмотического потенциала сока, выжатого из листьев растений различных экологических типов. Приведенная амплитуда получена из различий между самым низким и самым высоким значениями, найденными для видов, принадлежащих к одной и той же экологической группе

Растения, которые без вреда переносят большие колебания осмотического потенциала, называются эвригидрическими. Стеногидрические1 виды устойчивы лишь в пределах ограниченной осмотической амплитуды (рис. 6.28).
1 Данные термины не являются широко распространенными. — Примеч. ред.
В естественных условиях клетки и ткани растения лишь изредка достигают точки полной тургесцентности (р = П,![]() ), но обладают более или менее отрицательным водным потенциалом. Его можно измерить, например, компенсационным методом (рис. 6.29), в котором сегменты ткани с известной массой уравновешивают в средах с различными водными потенциалами (например, в растворах сахарозы), а затем снова взвешивают.
), но обладают более или менее отрицательным водным потенциалом. Его можно измерить, например, компенсационным методом (рис. 6.29), в котором сегменты ткани с известной массой уравновешивают в средах с различными водными потенциалами (например, в растворах сахарозы), а затем снова взвешивают.
Рис. 6.29. Компенсационный метод определения водного потенциала кусочков ткани посредством регистрации изменений веса до и после погружения в растворы непроникающего осмотика с различными осмотическими потенциалами (водными потенциалами)
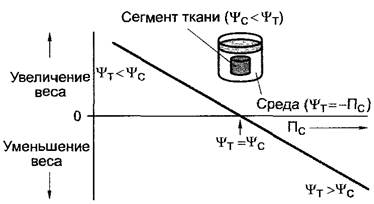
В гипертонической среде![]() ткань будет терять воду, в гипотонической среде
ткань будет терять воду, в гипотонической среде ![]() напротив, поглощать ее. Из графика изменений масса в зависимости от
напротив, поглощать ее. Из графика изменений масса в зависимости от![]() путем интерполяции можно найти точку, в которой не обнаруживаются изменения массы сегмента ткани (
путем интерполяции можно найти точку, в которой не обнаруживаются изменения массы сегмента ткани (![]() (см. выше), и отсюда
(см. выше), и отсюда
Таким образом, водный потенциал ткани![]() равен осмотическому потенциалу -Пс того раствора, в котором в ткани не наблюдается нетто-потока воды (и потому нет изменений массы).
равен осмотическому потенциалу -Пс того раствора, в котором в ткани не наблюдается нетто-потока воды (и потому нет изменений массы).
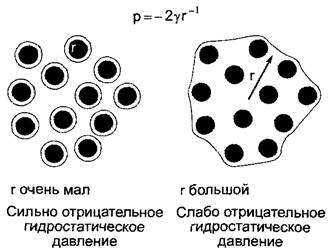
В водном обмене наряду с осмотическим движением молекул воды между областями с различными концентрациями растворенных веществ в протоплазме и особенно в клеточных стенках играют роль матричные эффекты. Эти чисто физические процессы в совокупности объединены термином набухание. Речь идет здесь, с одной стороны, о формировании водных оболочек (гидратации) вокруг полярных макромолекул, как, например, полисахариды и белки, и, с другой стороны, о капиллярных явлениях, как, например, капиллярные включения воды между микрофибриллами и межмицеллярными пространствами клеточной стенки. В обоих случаях влияния на водный потенциал можно оценить исходя из формирования локальных сильно отрицательных гидростатических давлений в тонких (толщиной в несколько слоев молекул) сильно искривленных водных пленках. Причиной является высокое поверхностное натяжение воды (у = 7,28 • 10-8 МПа • м).
Соотношение между потенциалом давления р из уравнения водного потенциала (уравнения 6.14; 6.15) и поверхностным натяжением воды у выражается уравнением
![]()
где г — радиус кривизны мениска.
Если г очень мал, то локальное гидростатическое давление становится очень отрицательным и соответственно таким же становится водный потенциал. Это происходит, например, в том случае, если клеточные стенки и протопласты высыхают (в сухих семенах или талломах лишайников), но имеет определенное значение также для клеточных стенок корней на сухих почвах, у транспирирующих листьев, и обусловливает сильно отрицательные водные потенциалы сухих почв.
С другой стороны, гидратированные структуры в водной среде (например, белки в водном растворе, структурные полисахариды в насыщенных водой клеточных стенках) вносят лишь небольшой вклад в общий водный потенциал, поскольку г становится сравнительно высоким (рис. 6.30).
Рис. 6.30. Появление локально очень отрицательного гидростатического потенциала давления в сильно искривленных тонких водных пленках. Слева, например, гидратные оболочки вокруг филаментов клеточной стенки, почвенных коллоидов, белков. При более полном насыщении водой (справа) эффекты из-за больших радиусов весьма ограничены. Поэтому белки цитоплазмы вносят лишь крайне ограниченный вклад в создание водного потенциала клетки
В системах, в которых водный потенциал определяют матричные эффекты (например, в сухих семенах и клеточных стенках, а также в почвах), и поэтому сложно определить по отдельности р и -П, оказывается пригодным их совместное определение, и эта величина обозначается как матричный потенциал т (он именуется также матричным водным потенциалом). Следует также заметить, что в создании матричного потенциала участвуют не новые силы, а исключительно те, что были рассчитаны из уравнения химического потенциала воды (уравнение 6.10). Из этого следует, что не нужно добавлять к общему уравнению водного потенциала дополнительную составляющую т.
С учетом сказанного в разделах 6.3.1 и 6.3.2 удастся лучше понять водный обмен всего растительного организма.
6.3.3. Поглощение воды растением
Талломные растения, у которых еще не выработана защита от транспирации, способны поглощать воду из влажных подстилок или после увлажнения каплями дождя или росы всей поверхностью, причем значительную роль могут играть не только осмос в оводненном состоянии талломов, но и матричные потенциалы у высушенных экземпляров. Некоторые водоросли, лишайники и определенные мхи при высыхании развивают настолько значительные отрицательные водные потенциалы (< -100 МПа, см. табл. 6.11), что еще способны поглощать водяной пар из воздуха в объеме, позволяющем сохранять положительный нетто-фотосинтез без поступления жидкой воды. Процесс набухания также вносит вклад в поглощение воды сухими семенами.
Таблица 6.11. Относительная концентрация водяного пара (процентная относительная влажность) воздуха, который находится в равновесии с раствором с определенным осмотическим потенциалом при 20 °С в закрытой системе
Относительная влажность воздуха, % |
-П, МПа |
Относительная влажность воздуха, % |
-П, МПа |
100 |
0 |
94,0 |
-8,32 |
99,5 |
-0,67 |
93,0 |
-9,79 |
99,0 |
-1,35 |
92,0 |
-11,2 |
98,5 |
-2,03 |
91,0 |
-12,6 |
98,0 |
-2,72 |
90,0 |
-14,1 |
97,5 |
-3,41 |
80,0 |
-30,1 |
97,0 |
-4,10 |
70.0 |
-48,1 |
96,0 |
-5,50 |
60,0 |
-68,7 |
95,0 |
-6,91 |
50,0 |
-93,3 |
Погруженные водные растения, которые имеют весьма проницаемую кутикулу или совсем лишены ее, поглощают воду всей поверхностью посредством осмоса. У некоторых наземных растений на надземных частях имеются определенные участки, пропускающие воду, например, волоски, основания внутренней стороны пар игл (например, у сосны) или специальные, способные к набуханию «сосущие чешуи» (например, у эпифитных бромелиевых, Bromeliaceae, см. 1Î.2, рис. 4.74). Эти пропускные участки не кутинизированы или кутинизированы лишь слабо и поглощают воду в основном посредством капиллярных явлений. Они функционируют преимущественно в условиях сухости посредством соответствующего изменения положения при сильной потере воды.
Воздушные корни некоторых эпифитов, например, орхидей рода Dendrobium, имеют водопоглощающую ткань веламен (см. 4.3.3.3, рис. 4.73), которая удерживает воду капиллярными силами. Из этого резервуара вода поглощается корнями осмотическим способом.
Наземные высшие растения (папоротники, семенные растения) поглощают воду преимущественно корнями (за исключением только что упомянутых специализированных образований, относящихся к побегу), так как обычно кутикула или пробковая ткань побега оказывают большое сопротивление диффузии воды. Вследствие этого поглощение воды надземными частями даже после увлажнения росой или дождем вряд ли играет какую-то роль. Вода не проникает в открытые устьица побега также из-за своего высокого поверхностного натяжения; предполагают, что тоже имеет место и в чечевичках.
Процесс поглощения воды корнем можно разделить на несколько этапов:
✵ поглощение воды клетками из апопласта;
✵ дополнительное поступление поглощенной воды из почвенного раствора в апопласт;
✵ транспорт воды внутри корня до сосудов ксилемы.
Поглощение воды корнем из почвы возможно лишь тогда, когда существует соответствующий градиент водного потенциала![]() . Водный потенциал почвы лишь частично определяется осмотическими явлениями, так как почвенный раствор очень разбавлен (типичные величины для
. Водный потенциал почвы лишь частично определяется осмотическими явлениями, так как почвенный раствор очень разбавлен (типичные величины для![]() близки к -0,02 МПа, а в засоленных почвах ≤ -0,2 МПа). В значительной части это обусловлено сильно колеблющимся из-за влажности почвы потенциалом давления (матричным потенциалом) (происхождение — см. уравнение 6.34). В насыщенных водой почвах после выпадения осадков или в более глубоких слоях почвы вблизи горизонта грунтовых вод этот потенциал близок к нулю
близки к -0,02 МПа, а в засоленных почвах ≤ -0,2 МПа). В значительной части это обусловлено сильно колеблющимся из-за влажности почвы потенциалом давления (матричным потенциалом) (происхождение — см. уравнение 6.34). В насыщенных водой почвах после выпадения осадков или в более глубоких слоях почвы вблизи горизонта грунтовых вод этот потенциал близок к нулю![]()
Часть воды, попавшей в почву в виде осадков, адсорбируется как «адгезионная, или пленочная, вода» и удерживается в капиллярах, часть просачивается как «гравитационная вода» до горизонта грунтовых вод. В общем доступной для корневой системы остается лишь большая или меньшая часть «пленочной воды». Водоудерживающая способность почвы для «пленочной воды» (количество грамм Н2О на 100 мл объема почвы) обозначается как ее влагоемкость. Она повышается с увеличением содержания в почве тонкодисперсного и органического материала и поэтому возрастает в ряду от песка до суглинка и глины и особенно до болотистых почв. Чем мельче и пористее частицы почвы, тем более отрицательными становятся водные потенциалы при ее высыхании (см. уравнение 6.34).
По мере убывания содержания воды в почве ее водный потенциал становится отрицательнее. Он легко достигает значений -2 МПа и ниже. Водные потенциалы корня, как уже обсуждалось, в основном определяются осмотическими потенциалами клеточного сока. Они могут в определенных границах приспосабливаться к текущим потребностям посредством изменений концентрации осмотиков, они также существенно колеблются в зависимости от вида. Так, у фасоли (Phaseolus) осмотические потенциалы в клетках корня составляют от -0,2 до -0,35 МПа, у пеларгонии (Pelargonium)- -0,5 МПа, у галофитов (со
леросов) — менее -2 МПа, а у пустынных растений — даже меньше -10 МПа. Этих величин достаточно для того, чтобы извлекать из любой почвы большую часть ее «пленочной воды».
Поглощение воды корнем можно охарактеризовать формулой
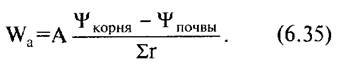
Соответственно этому масса воды, поглощенной корневой системой в единицу времени Wа, прямо пропорциональна поверхности корня А, приспособленной для поглощения воды (по существу, поверхности корневых волосков) и разности водных потенциалов корня и почвы и обратно пропорциональна сумме всех сопротивлений транспорту воды в почве и при переходе из почвы в растение (Σг).
Поверхность корневых волосков, приспособленная для поглощения воды, часто достигает очень больших размеров. Так, у одного растения ржи, которое пронизывало корнями объем почвы, равный 56 л, общее число живых корневых волосков составляло 1,43 • 1010, их общая поверхность оценивалась в 400 м2 и соответственно превосходила относящуюся к транспирации общую контактную поверхность клеток листового мезофилла с межклетниками (см. 6.3.4.1) более чем в 10 раз.
Зоны корневых волосков кончиков корней, следовательно, являются преимущественными участками поглощения как воды, так и ионов. Однако корневые волоски отсутствуют у растений с эктомикоризой (см. 9.2.3). В этом случае симбиотический микоризный гриб берет на себя функции корневых волосков. Корневые волоски входят в очень тесный контакт с почвой (см. рис. 6.20). Из апопластов, находящихся в равновесии с почвенным раствором, вода посредством осмоса поступает в корневые волоски, отчего гидростатическое давление в них снижается. Образующийся в почвенном растворе градиент давления дополнительно поставляет воду в апопласт посредством массового тока (капиллярный ток). Поглощение почвенной воды в зоне корневых волосков снижает гидростатическое давление и, следовательно, водный потенциал почвы в зоне корневых волосков по отношению к более глубоким и соответственно не пронизанным корнями соседним зонам. Вода перемещается массовым током по градиенту давления.
Однако эта проводящая способность почвы в значительной степени зависит от типа почвы и даже у тонкопористых почв (например, глины) со сравнительно хорошей проводимостью осуществляется лишь очень медленно и на очень
короткие расстояния (самое большее — несколько сантиметров). Растение преодолевает эту трудность за счет того, что корни растут вслед за водой. При этом части корневой системы могут отмирать, в то время как другие интенсивно разрастаются в более богатых водой участках почвы, так что вся корневая система может развиваться весьма асимметрично. При соответствующих градиентах водного потенциала корни могут также отдавать воду в почву. Следовательно, через корни возможен транспорт воды от более влажных, в основном более глубоких слоев почвы к более сухим, преимущественно вышележащим слоям (англ, hydraulic lift — гидравлический подъем) (см. рис. 13.25).
Существенным уменьшением поглощения воды при пониженных температурах (для многих растений уже при нескольких градусах выше 0 °С) наряду с повышением сопротивления транспорту в почве и понижением проницаемости плазмалеммы для воды прежде всего объясняется торможение роста корней. При температуре менее -1 °С пленочная вода в почве замерзает и поглощение воды становится невозможным («зимняя засуха»); последствия этого часто ошибочно истолковывают как вымерзание.
Если почва высыхает так сильно, что вся корневая система не может получить достаточное количество воды или не поглощает ее совсем, либо даже теряет воду из-за обращения разности водных потенциалов, то наступает увядание растений, которое становится необратимым при определенном водном потенциале почвы. Приспособленные к влажным условиям травы достигают этого состояния при величинах водного потенциала почвы приблизительно от -0,7 до -0,8 МПа, большинство полезных сельскохозяйственных растений — при величинах от -1 до -2 МПа, растения умеренно сухих биотопов и различные древесные растения — при величинах от -2 до -3 МПа. В сельскохозяйственной практике за точку устойчивого завядания принимают величину водного потенциала почвы -1,5 МПа.
В корне вода может диффундировать по симпласту по градиенту осмотического потенциала, становящегося более отрицательным в направлении эндодермы. Поскольку в коре вода может также поглощаться посредством осмоса клетками из апопласта вплоть до преграды в виде поясков Каспари, вероятно и дополнительное
перемещение воды в апопласт из периферической зоны корня путем радиального массового тока. В центральный цилиндр корня вода поступает осмотическим путем, в особенности при хорошем водоснабжении растения, и более ограниченно — при отсутствии транспирации (например, ночью). Транспорт ионов из эндодермы и сосудистой паренхимы в апопласт центрального цилиндра ведет к снижению в нем водного потенциала, что влияет на переход воды из клеток в апопласт.
При условиях хорошего водоснабжения, ограниченной транспирации в ксилеме корней развивается положительное гидростатическое давление — корневое давление. Вторая существенная функция поясков Каспари, как и эндодермы, таким образом, наряду с барьерной функцией для растворимых составных частей почвенного раствора, диффундирующих через апопласт в направлении центрального цилиндра, состоит в «сгущении» содержимого центрального цилиндра, что обеспечивает существование корневого давления. Это давление можно определить, если удалить побег немного выше корня и надеть манометр на остаток побега, и обычно оно составляет <0,1 МПа, но у берез может повышаться до 0,2 МПа и более, а у растений томатов может превосходить 0,6 МПа.
Таким образом, корневое давление при определенных условиях (см. выше) вносит вклад в обеспечение дальнего транспорта воды.
При менее обильном водоснабжении или более интенсивной транспирации (см. 6.3.4) в ксилему все же постоянно поступает столько воды, что в ней не создается положительное гидростатическое давление, и в области корня также доминирует отрицательное гидростатическое давление (и соответственно отрицательный водный потенциал). Наличие отрицательного водного потенциала может приводить к заимствованию воды из протопластов клеток эндодермы и сосудистой паренхимы, вследствие чего их осмотический потенциал (и в результате водный потенциал) становится более отрицательным: вода или диффундирует по симпласту из периферической зоны корня, или перетекает из апопласта коровой паренхимы корня. В этой физиологической ситуации очень велико также значение поясков Каспари как барьера, так как они снижают неконтролируемое «просачивание» почвенного раствора.
6.3.4. Выделение воды растением
Большую часть поглощенной воды растение выделяет в виде водяного пара («транспнрационная вода», транспирация, см. 6.3,4.1), часть воды служит для увеличения объема растущего растения («вода роста»). В особых случаях вода выделяется из растения в виде капель и струек (гуттация, см. 6.3.4.2).
У быстро растущих травянистых растений вода роста может составлять значительную часть общего водного баланса, например, у кукурузы на нее приходится 10 — 20 % транспирационной воды. Транспирация является существенной движущей силой дальнего транспорта воды в ксилеме при открытых устьицах. Ток воды по ксилеме поддерживается и при закрытых устьицах (например, ночью), и при сниженной (в случае высокой относительной влажности воздуха) транспирации, а также и при почти полностью подавленной (экспериментально) транспирации (в воздухе, насыщенном водяными парами). Ксилемный ток осуществляется вследствие:
✵ создания корневого давления (см. 6.3.3);
✵ наличия сильно отрицательных осмотических потенциалов периферических органов с активным обменом веществ, особенно фотосинтетически активных листьев, которые в побеге также «связывают» преобладающую часть воды роста;
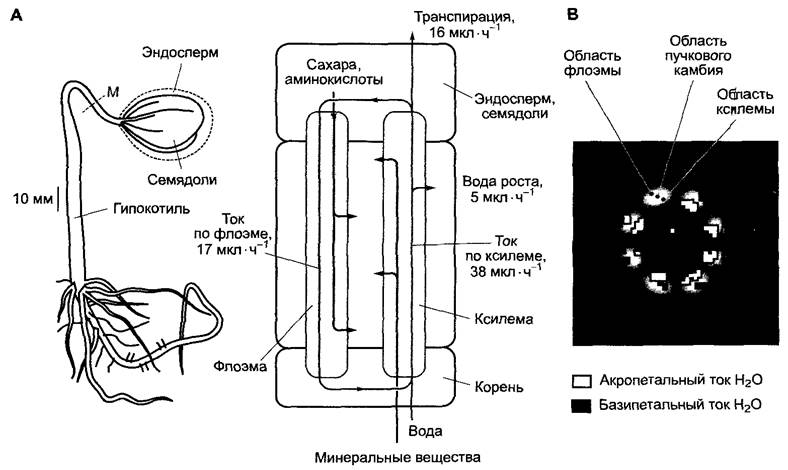
✵ потока воды по флоэме, который в местах загрузки флоэмы выравнивается ксилемной водой (см. 6.8.2); этот внутренний круговорот воды лишь недавно впервые был наглядно показан непосредственно в живом растении методом ядерно-магнитного резонанса (рис. 6.31); его относительные величины оцениваются у деревьев приблизительно как 1—3%, у кукурузы — как 5 —10 % транспирационной воды.
Рис. 6.31. Водный режим и внутренний круговорот воды у проростков клещевины (Ricinus) в возрасте 6 дней (определены при 95%-й относительной влажности воздуха и 28 °С в темноте) (по W. Kuckenberger, с любезного разрешения): А — внешний вид проростка (слева) и водный баланс (справа). Измеренная скорость потока в месте измерения М составляла: в ксилеме — 1,7 м • ч-1 (акропетально), во флоэме — 2,1 м • ч-1 (базипе- тально); В — наличие тока воды в живом проростке наглядно показано методом ЯМР, т.е. ядерно- магнитного резонанса (англ. — 1H-NMR, nuclear magnetic resonance). ЯМР — изображение получено для среза стебля толщиной 1 мм в месте измерения М (см. А). Во флоэме (наружные участки) вода перемещается от верхушки побега в направлении к корню (базипетально), в ксилеме — в противоположном направлении (акропетально), что иллюстрируется изображением на экране (базипетальный и акропетальный ток). В проростке по ксилеме транспортируются минеральные вещества и вода для роста и заполнения флоэмы. Вследствие испарения происходит постоянная потеря воды. По флоэме транспортируются питательные вещества из эндосперма
В подобных ситуациях, т. е. при сниженной транспирации, содержимое ксилемы переносится намного медленнее, чем при более интенсивной транспирации, однако это компенсируется более высокой концентрацией ионов в ксилеме, так что независимо от интенсивности транспирации обеспечивается достаточное снабжение растения минеральными веществами. Об этом свидетельствует также быстрый рост растений в опыте при более низкой относительной влажности воздуха (по сравнению с более высокой) и при сниженной (до 15 раз) транспирацией. Акропетальный (направленный от основания к верхушке побега) транспорт воды можно обнаружить
в побегах погруженного и потому не транс- пирирующего водного растения, лютика Ranunculus trichophyllus. Его скорость (>80 см • ч-1) вполне достаточна для поддержания максимального роста и снабжения питательными веществами.
Следовательно, транспирация должна рассматриваться скорее, как неизбежное зло, а не как жизненно необходимый транспортный механизм наземных растений.
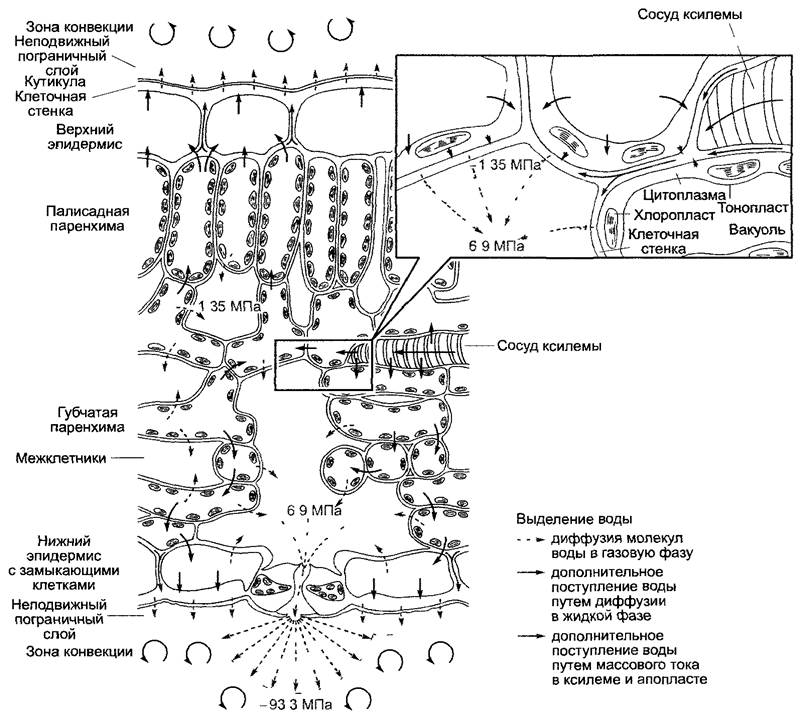
Переход молекул воды из жидкой фазы в газовую фазу (транспирация, испарение) осуществляется на всех поверхностях растения, граничащих с воздухом, не насыщенным водяными парами. У слоевищных растений это внешние поверхности таллома, у кормофитов — во-первых, внешние поверхности побегов, обычно кутинизированные или опробковевшие для снижения транспирации, во-вторых — поверхности клеток внутри побега, граничащие с межклетниками Из межклетников водяной пар диффундирует из растения через устьица, причем сначала он должен преодолеть пограничный слой (тонкий слой неподвижного воздуха, примыкающий непосредственно к поверхности растения), прежде чем достигнет свободной атмосферы, где путем перемешивания (конвекции) быстро удаляется от растения (рис 6.32).
Рис. 6.32. Транспорт воды в листе, имеющем устьица на нижней стороне. Серым тоном выделены объемы, заполненные жидкой водой, не выделены пространства, заполненные газом (воздух межклетников, атмосферный воздух) Цифры показывают величины водных потенциалов -1,35 МПа — 99 % относительной влажности воздуха, -6,9 МПа — 95 % и -93,3 МПа — 50 % (в каждом случае при 20 °С, табл 6 11)
Движущей силой транспирации служит также градиент водного потенциала, причем критической областью является разность водных потенциалов между воздухом
внешней среды и воздухом межклетников. Водный потенциал воздуха рассчитывается по формуле
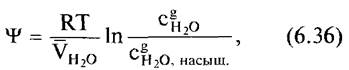
где R — универсальная газовая постоянная; Т — абсолютная температура;![]() парциальный молярный объем жидкой воды;
парциальный молярный объем жидкой воды;![]() реальная концентрация воды в газовой фазе;
реальная концентрация воды в газовой фазе;![]() насыщающая концентрация воды в газовой фазе;
насыщающая концентрация воды в газовой фазе;
называется также относительной влажностью воздуха и обычно выражается в процентах.
При снижении степени насыщения воздуха газообразной водой водный потенциал очень быстро снижается до весьма отрицательных значений (см. табл. 6.11). Относительная влажность воздуха межклетников может достигать 99 %![]() = -1,35 МПа), в подустьичной полости при открытых устьичных щелях — около 95 %
= -1,35 МПа), в подустьичной полости при открытых устьичных щелях — около 95 % ![]() -6,9 МПа), а при средней влажности атмосферного воздуха непосредственно над устьичной щелью — около 50 %
-6,9 МПа), а при средней влажности атмосферного воздуха непосредственно над устьичной щелью — около 50 % ![]() = -93,3 МПа). Некоторые значения средних водных потенциалов облиственных ветвей различных групп растений представлены на рис. 6.33.
= -93,3 МПа). Некоторые значения средних водных потенциалов облиственных ветвей различных групп растений представлены на рис. 6.33.
Рис. 6.33. Колебания водного потенциала листьев и ветвей у различающихся экологически групп растений. Измерения проведены с применением камеры давления Сколандера (см. рис. 6.39) днем при сильном солнечном излучении

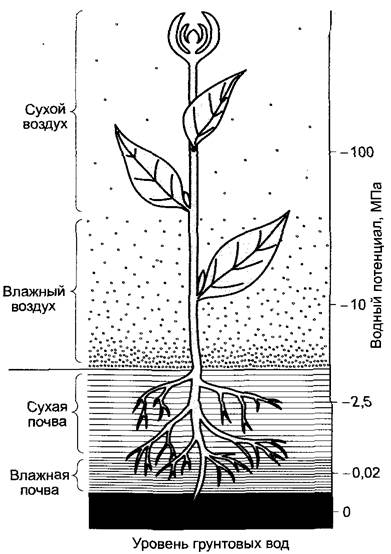
Следовательно, растение как бы «заполняет промежуток» между сравнительно высоким водным потенциалом почвы и низким водным потенциалом воздуха (рис. 6.34). Движущей силой транспирации является чрезвычайно большая разность водных потенциалов между ненасыщенным водяными парами, воздухом внешней среды и воздухом межклетников (соответственно пограничного слоя, не смешанного с окружающим). Молекулы воды диффундируют в газах намного быстрее, чем в жидкой воде. Потеря воды из межклетников (соответственно из пограничного слоя) стимулирует диффузию молекул воды из апопласта в воздух межклетников. В апопласте, таким образом, развиваются сильно отрицательные гидростатические давления (уравнение 6.34), которые влияют на проведение из сосудов ксилемы и соответственно из живых клеток тканей листа. Проводящие сосуды листьев на концах сильно разветвлены, так что большинство клеток листа отстоят от ближайших сосудов ксилемы не более чем на 0,5 мм.
Рис. 6.34. Перепады водного потенциала между почвой, растением и воздухом. Самый большой скачок потенциала происходит не между почвой и растением, а между растением и воздухом (см. рис. 6.32)
Увеличение транспирирующей поверхности также приводит к усилению транспирации, как и все факторы, которые делают более крутым падение водного потенциала между растением и воздухом. Повышение температуры воздуха уменьшает относительную влажность воздуха и этим снижает его водный потенциал (![]() воздуха становится более отрицательным). Повышение температуры транспирирующих органов (например, листьев) вследствие поглощения солнечного излучения стимулирует переход воды из жидкой фазы в газовую. Высокое содержание воды в растении
воздуха становится более отрицательным). Повышение температуры транспирирующих органов (например, листьев) вследствие поглощения солнечного излучения стимулирует переход воды из жидкой фазы в газовую. Высокое содержание воды в растении ![]() слабоотрицателен) также увеличивает разность потенциалов. Ветер уменьшает толщину пограничного слоя с его относительно высоким содержанием водяного пара и этим повышает градиент потенциала. Сопротивление пограничного слоя транспорту водяного пара при скорости ветра 0,1 м с-1 составляет около 1 — 3 с см-1, при 10м с1 оно снижается до 0,1 — 0,3 с см 3
слабоотрицателен) также увеличивает разность потенциалов. Ветер уменьшает толщину пограничного слоя с его относительно высоким содержанием водяного пара и этим повышает градиент потенциала. Сопротивление пограничного слоя транспорту водяного пара при скорости ветра 0,1 м с-1 составляет около 1 — 3 с см-1, при 10м с1 оно снижается до 0,1 — 0,3 с см 3
Основными транспирирующими органами кормофитов служат листья. Из-за больших поверхностей облиственных растений потери воды вследствие транспирации часто весьма значительны. Поскольку растение не должно страдать от дефицита воды во время максимальной транспирации, по крайней мере большая часть этих потерь воды должна возмещаться путем поглощения воды из почвы.
Рассчитано, что в буковом лесу около 60 % осадков, выпадающих на него в течение года, возвращается в атмосферу в виде водяного пара Подсолнечник способен испарить в течение солнечного дня 1 л пара, береза (имеющая приблизительно 200000 листьев) — до 60 — 70 л, а в особенно жаркие и сухие дни даже до 400 л В пустыне Кара-Кум у растения из семейства бобовых Smurnovia turkestana потеря воды уже в течение 1 ч в 7 раз превышает его водный запас У некоторых растений на сухих склонах Кайзерштуля дневная транспирация в 12 раз больше содержащейся в них воды. По некоторым оценкам, только за 4 000 лет все запасы земной воды проходят через корневые системы растений к транспирирующим органам, откуда выделяются в форме водяного пара.
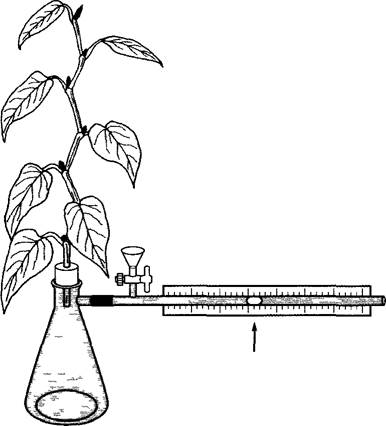
Транспирацию растения или части растения за небольшие отрезки времени можно измерить путем взвешивания в начале и в конце опыта; потери массы вследствие дыхания или увеличение ее вследствие фотосинтеза не играют существенной роли за короткие промежутки времени. Более точные и долговременные измерения, а также определения на крупных растениях требуют других методов. Поскольку потеря воды компенсируется ее поглощением, транспирацию можно определить также, используя потометр (рис. 6.35). Комбинация взвешивания и измерения при работе с потометром позволяет определить, как поглощение, так и отдачу воды, т.е. рассчитать водный баланс растения (см. 6.3.6, 13.5).
Рис. 6.35. Схема простого потометра. Стрелка указывает на пузырек воздуха, за перемещением которого в капилляре можно наблюдать.
Водный потенциал надземной части высшего растения (см. рис. 6.34) гораздо ближе к водному потенциалу почвы по сравнению с атмосферой. Это объясняется значительными сопротивлениями диффузии водяного пара, которые у растений создаются на транспирирующих поверхностях, прежде всего внешних, для защиты от транспирации.
Интенсивность транспирации (ТR!!!!!, моль м-2 с-1) вычисляется как
![]()
где![]() реальная концентрация воды в газовой фазе; Σг — сумма всех сопротивлений диффузии.
реальная концентрация воды в газовой фазе; Σг — сумма всех сопротивлений диффузии.
Защита от транспирации обеспечивается прежде всего кутикулой (химический состав см. 6.17.3), которая впервые появляется у мхов и (наряду с наличием суберина и лигнина) служит необходимой предпосылкой для развития крупных наземных растений с регулируемым водным обменом (так называемых гомойогидрических растений, см. 13.5). Выделенные сплошные листовые кутикулы имеют чрезвычайно ограниченную проницаемость для воды (коэффициент проницаемости от 10-7 до 10-8- см • с-1); это объясняется главным образом наличием в ней воска. Водопроницаемость интактного листа еще больше снижается вследствие наложения дополнительных слоев воска на кутикулу (см. рис. 3.11) и отложения кутина во внешних стенках эпидермиса. Покров из отмерших волосков, который можно видеть на некоторых листьях (например, у эдельвейса), также служит для уменьшения транспирации, создавая безветренные, насыщенные водяными парами пространства (см. рис. 3.14). Аналогичное влияние оказывает также погружение устьиц в углубления, защищенные от ветра.
Кутикулярная транспирация обычно не превышает (даже у нежных листьев влажных местообитаний) 10 % от испарения со свободной водной поверхности равной площади (эвапорации, т.е. испарения без сопротивления со стороны диффузии и при неограниченном восполнении воды). У хвои и жестких листьев она составляет 0,5 %, а у кактусов, которые вынуждены в течение долгих засушливых периодов сохранять от испарения запасную воду, — лишь 0,05 % от эвапорации.
Как и кутин, слои суберина служат изолирующим материалом, например, в покровных тканях, пробке и коре (см. 3.2.2.1, 3.2.2.2). На этом основано закупоривание бутылок шампанского корковой пробкой, обеспечивающей непроницаемость для воды и газа. Длительность хранения клубней картофеля также обусловлена их тонкой пробковой «кожурой», поэтому очищенный картофель быстро высыхает.
Подобно покрытию кутином и суберином, изоляция органов растений с помощью лигнина (структура и биосинтез, см. 6.17.3) может не только снизить выход
водяного пара, но и ограничить диффузию других жизненно важных для растения газов (прежде всего СО2 для фотосинтеза, см. 6.4). Поэтому на листьях растений — важнейших органах, осуществляющих газообмен, а также на других зеленых органах (на первичных осях побегов, на плодах) возникли регулируемые поры, устьица (см. 3.2.2.1). Опробковевшие ткани выделяют воду через нерегулируемые поровые системы, чечевички (см. 3.2.2.2), с локально сниженными сопротивлениями диффузии.
Устьица, с одной стороны, облегчают поступление СO2, необходимого для фотосинтеза (или для темповой фиксации СО2 у САМ-растений, см. 6.5.9), вследствие снижения сопротивления диффузии (открыванием устьиц); с другой стороны, они уменьшают устьичную транспирацию при напряженном водном режиме или при отсутствии условий для фотосинтеза (в темноте) путем повышения сопротивления диффузии (закрыванием устьиц).
Полностью открытые устьица резко снижают сопротивление диффузии по сравнению с сопротивлением при кутикулярной транспирации (табл. 6.12). Различия между видами и локальными формами растений зависят при этом от расположения устьиц (на обеих сторонах листа или только на нижней), их частоты, величины, а также от особенностей строения («геометрии»).
Таблица 6.12. Транспирация листьев различных растений (в мг Н2О на 1 дм2 поверхности обеих сторон листа в час) при эвапорации (в эвапориметре Пише)
3 360 мг Н2О дм-2 ч 1
Растение |
Общая транспирация при открытых устьицах |
Кутикулярная транспирация после закрывания устьиц |
Кутикулярная транспирация, %, от общей транспирации |
Травянистые растения солнечных местообитаний |
|||
Coronilla varia |
2 000 |
190 |
9,5 |
Stachys recta |
1 800 |
180 |
10 |
Oxytropis pilosa |
1 700 |
100 |
6 |
Теневыносливые травы |
|||
Pulmonaria officinalis |
1000 |
250 |
25 |
Impatiens noli-tangere |
750 |
240 |
32 |
Asarum europaeum |
700 |
80 |
11,5 |
Oxalis acetosella |
400 |
50 |
12,5 |
Деревья |
|||
Betula pendula |
780 |
95 |
12 |
Fagus sytvatica |
420 |
90 |
21 |
Picea abies |
480 |
15 |
3 |
Pinus sylvestris |
540 |
13 |
2,5 |
Вечнозеленые Ericaceae |
|||
Rhododendron |
600 |
60 |
10 |
ferrugineum |
|||
Arctostaphylos uva-ursi |
580 |
45 |
8 |
При полностью открытых устьицах лист может потерять вследствие транспирации максимально 50 — 70 % того количества водяного пара, которое путем эвапорации испарилось бы с равной водной поверхности. Это удивительно много, так как устьиц хотя и насчитывается несколько сотен на квадратный миллиметр, но их общая площадь при максимальной степени открытости из-за ограниченной ширины устьичных щелей (несколько мкм) редко достигает 1 — 2 % площади листа.
В модельных опытах установлено, что множество маленьких пор обеспечивают значительно большее пропускание воды, чем небольшое количество больших пор при равной общей площади. Это обусловлено «краевым эффектом», т.е. тем, что молекулы водяного пара, выходящие с краев поры, имеют и сбоку свободное поле для диффузии, в то время как молекулы, диффундирующие из центральной части, со всех сторон окружены соседними молекулами (см. рис. 6.32). Вероятно, по этой же причине первое же незначительное раскрывание устьиц, исходно полностью закрытых, оказывает самое сильное влияние на транспирацию.
Дневной ход транспирацииу кормофитов в большинстве случаев обнаруживает характерную последовательность: с началом освещения транспирация начинается вследствие фотоактивного открывания устьиц (см. 8.3.2.5), затем она возрастает при полностью открытых устьицах до полудня благодаря увеличивающемуся нагреванию листа и воздуха (уменьшению относительной влажности воздуха), а потом снова снижается до тех пор, пока при наступлении сумерек устьица вновь не закроются. При возрастающей напряженности водного режима (см. 13.5.2) это приводит к более раннему ограничению открывания устьиц, редко — к полуденному ограничению. Если в течение дня поступление воды больше не возмещает ее расход, то этот дефицит обычно может снова уравновешиваться в течение прохладной и относительно влажной ночи.
В связи со значимостью устьиц для общего газообмена у растений всех видов с функционирующими устьицами важную роль в физиологической регуляции газообмена играют факторы, регулирующие ширину устьичных щелей. Они будут рассмотрены ниже (см. 6.5.7; механизм регуляции, см. 8.3.2.5).
Чечевички также являются местами сниженного сопротивления диффузии водяного пара. Так. коэффициент проницаемости чечевичек у перидермы березы примерно в десять раз выше, чем у сплошной перидермы, но они в противоположность устьицам не регулируются физиологически.
Транспирацнонный коэффициент (кт) показывает, сколько воды выделяется при транспирации, когда фиксируется 1 г СO2, т. е. это мера экономии воды. Часто используется обратная величина (кт), обозначаемая как эффективность использования воды, или продуктивность транспирации:
![]()
Употребляются также величины, выраженные в молях. Транспирационный коэффициент видо- и сортоспецифичен и в значительной степени зависит от типа фотосинтеза: 200—800 у С3-растений, 200 — 350 у С4-растений, 30—150 у САМ-растений (фиксация СО2 ночью) (С4- и САМ- метаболизм — см. 6.5.8 и 6.5.9).
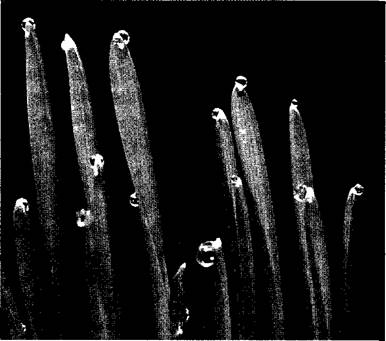
Вероятно, необходимостью поддерживать ток воды в растении даже в отсутствие транспирации объясняется явление гуттации, т. е. выделение капельножидкой воды. Соответственно это явление наблюдается прежде всего при высокой относительной влажности воздуха, в частности в тропических дождевых лесах, а в нашем климате, например, ночью. Такие капли выделяются на растениях в определенных местах, чаще всего на листьях, через гидатоды (см. 3.2.2.1) или через железистые волоски (волосковые гидатоды). Нередко эти капли ошибочно принимают за капли росы, например, у манжетки (Alchemilla), фуксии (Fuchsia), настурции (Tropaeolum) или на кончиках листьев многих злаков (рис. 6.36). У Colocasia nymphaeifolia (сем. Ароидные, Агасеае) растения из тропического дождевого леса, с одного крупного листа в течение одной-единственной ночи может «накапать» до 100 мл жидкости. Гуттация обнаруживается и у низших растений, прежде всего у грибов, например, у домового гриба Serpula (Merulius) la cry та ns, благодаря чему он и получил свое видовое название («слезящийся»).
Рис. 6.36. Гуттационные капли на кончиках листьев молодых растений пшеницы
Движущей силой для выделения гуттационной жидкости пассивными гидатодами, например, у листьев злаков, служит корневое давление (см. 6.3.3). Гидатоды представляют собой поровые системы, через которые содержимое ксилемы под действием собственного давления выступает наружу, причем часто проходит через водные устьица. Этот вид гуттации перестает работать, если гидатоды изолировать от корня. У активных гидатод (вероятно, они составляют большинство эпитемных гидатод), например, у настурции (Tropaeolum) и камнеломки (Saxifraga), и всех волосковых гидатод, например, у нута (Cicer), фасоли (Phaseolus) и др., имеются водные железы, работающие независимо от корневого давления. Механизмы выделения в данном случае еще не выяснены в деталях, как и для других желез.
Принято исходить из того, что осмотически активные вещества выводятся наружу, а вода следует за ними пассивно. Активные гидатоды, возможно, родственны солевым и нектарным железкам (см. 6.18). Гуттационная жидкость поставляет не чистую воду, а разбавленный водный раствор органических, а также и неорганических веществ.
Дальний транспорт воды осуществляется в элементах ксилемы (см. 3.2.4.2), которые специально приспособлены для этой функции. Движущей силой процесса у растущих и транспирирующих растений служит преимущественно присасывающее действие транспирации, а корневое давление1 (см. рис. 6.31) позволяет обеспечить путем массового тока воды по ксилеме распределение растворенных в ксилемном соке веществ, прежде всего элементов минерального питания (особенно у травянистых растений или проростков в фазах сильно уменьшенной или отсутствующей транспирации). Вода в этих случаях выделяется путем гуттации (см. 6.3.4.2; рис. 6.36), так что она не возвращается во флоэму и подлежит остаточной транспирации.
1 В связи с этим в отечественной литературе вводят понятие о верхнем концевом двигателе (транспирация) и нижнем концевом двигателе (корневое давление) водного потока. — Примеч. ред.
У проростка клещевины (Ricinus) методом ядерно-магнитного резонанса определили, что ток воды в ксилеме составлял 38 мкл • ч-1, во флоэме — 17 мкл • ч-1, при интенсивности транспирации 16 мкл • ч-1 и потребности в воде роста 5 мкл • ч-1. Следовательно, величина объемного тока воды в ксилеме равна точной сумме величин тока во флоэме + интенсивности транспирации + потребности в воде роста. Гуттации не происходит.
Выделение сока (пасоки) у различных древесных растений при повреждении ксилемы ранней весной («плач растений») также объясняется действием корневого давления.
Количества жидкости, вытекающей из ран, могут быть весьма значительны: за 24 ч виноградная лоза может потерять около 1 л жидкости, береза — до 5 л.
Анализы показали, что содержимое водопроводящих путей, как и гуттационная жидкость, и весенняя пасока растений, представляет собой не чистую воду, а разбавленный (0,1—0,4%-й) раствор неорганических веществ, сахаров, органических кислот, аминокислот, витаминов, гормонов и т.д. Известно, например, высокое (в среднем 2,5%) содержание сахаров (преимущественно сахарозы) в пасоке сахарного клена (Acer saccharum), который в Северной Америке используется для приготовления кленового сиропа (англ. — maple syrup). Сильное дерево поставляет в середине марта около 4 л пасоки в день и приблизительно 2 — 3 кг сахара в течение весны. Впрочем, к этому времени клен развивает не только корневое давление, но и избыточное давление в стволе, что показано на спиленных деревьях.
Однако по выделению жидкости после повреждения ксилемы нельзя заключить, что в проводящих путях деревьев, способных к плачу, в период сокодвижения, т.е. когда они лишены листьев, благодаря корневому давлению поддерживается интенсивный ток воды. В действительности в необлиственном стволе движение воды столь мало (проводилось термоэлектрическое измерение водного тока, рис. 6.37), что вряд ли могло бы удовлетворить потребности в воде или восполнить ее потери. Вероятно, значение пасоки и избыточного давления в ксилеме древесного растения, лишенного листвы, можно усмотреть в том, что с распусканием почек ранней весной, т.е. прежде чем новые листья сами начинают активно фотосинтезировать, уже имеется потребность в питательных веществах, которая частично покрывается за счет содержимого ксилемы.
Рис. 6.37. Термоэлектрическое измерение скорости тока в ксилеме; схема опытной установки. Проволочная петля нагревается электричеством в течение короткого периода (1—3 с), и прибытие тепловой волны «вниз по течению» регистрируется термоэлементом из сплава медь (Сu)-константан (Ко) на определенном расстоянии от места нагрева. Движущийся фронт нагретого ксилемного сока достигает сначала ближайшего места измерения и вызывает отклонение стрелки гальванометра. Когда сок проходит второе место измерения, и оно становится теплее первого, то происходит отклонение стрелки гальванометра в обратную сторону. Это возвратное отклонение служит явным доказательством прохода некоторого объема нагретой воды
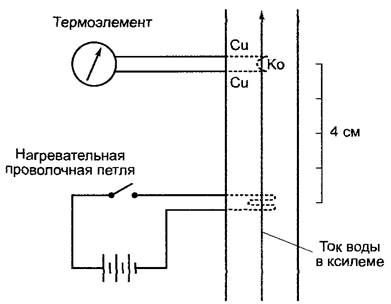
Происхождение отрицательного гидростатического давления (присасывающего действия транспирации) в апопласте транспирирующих листьев было объяснено в разделе 6.3.4.1. Отсюда по водопроводящим путям ксилемы до корня насквозь тянутся сплошные водные тяжи, которые вследствие присасывающего действия транспирации движутся черeз растение (транспирационный ток). Водопроводящие пуги — трахеи и/или трахеиды (см. 3.2.4.2) (отмершие проводящие элементы) — представляют довольно незначительное сопротивление движущейся воде благодаря отсутствию в них протопластов.
Слой цитоплазмы между вакуолью и клеточной стенкой (включая тонопласт и плазмалемму) одной-единственной клетки хары (Chara) обладает водопроницаемостью лишь около 10-4см c-1 МПа 1, что соответствует величине для 600 м сосновой древесины в продольном направлении и 3 мм в радиальном направлении
Общая площадь поперечного сечения водопроводящих элементов, сформированных в оси побега растения на грамм сырого веса листьев, обеспечиваемых водой, зависит от экотипа: растения влажных биотопов (ограниченная транспирация) показывают меньшие величины, чем растения сухих местностей (табл. 6.13). Даже внутри кроны одного дерева эти величины в отдельных сучьях или ветвях неодинаковы: например, верхний побег обеспечивается водой однозначно предпочтительнее.
Скорости транспорта в ксилеме можно определить термоэлектрическим методом (см. рис. 6.37). Измеренные величины значительно различаются у отдельных видов в зависимости от строения их древесины. По этому показателю были выделены три больших типа растений (голосеменные, рассеяннопоровые и кольцепоровые покрытосеменные), у которых заметно отличались максимальные и средние скорости транспирации (табл. 6.14).
Таблица 6.13 Площадь поперечного сечения водопроводящей системы у различных растений (мм2/г сырого веса листа)
Растение |
Срез, мм2 г 1 |
Кувшинка (черешки листьев) |
0,02 |
Травы подлеска |
0,01-0,80 |
Хвойные деревья |
0,30-0,61 |
Лиственные деревья |
0,25-0,79 |
Пустынные растения |
1,42-7,68 |
Таблица 6.14 Наибольшие полуденные скорости транспирационных токов различных типов растений, измеренных термоэлектрическим методом
Объект |
Скорость, м ч 1 |
Мхи |
1,2-2,0 |
Вечнозеленые хвойные деревья |
1,2 |
Лиственница |
1,4 |
Средиземноморские жестколистные растения |
0,4-1,5 |
Летнезеленые рассеяннопоровые лиственные деревья |
1-6 |
Кольцепоровые лиственные деревья |
4-44 |
Травянистые растения |
10-60 |
Лианы |
150 |
Применение закона Хагена —Пуазейля (уравнение 6.32) позволяет на основании измеренных скоростей тока определить величину разрежения (отрицательного гидростатического давления), которое при данной геометрии сосудов ксилемы и вязкости ее содержимого (приближена к вязкости воды, 103 МПа с) требуется для перемещения столба жидкости с соответствующей скоростью тока. Для средней скорости 16 м ч-1 и среднего радиуса сосуда 30 мкм (диаметр 60 мкм) получается величина-0,02 МПа м-1 (рис. 6.38). Поскольку свойства трахей и трахеид отличаются от идеальных капилляров, эта величина может быть минимальной. Отклонения реальных гидравлических проводимостей ксилемы от идеальных значений для различных растений приведены в табл. 6.15. Следовательно, реальные значения для требуемых разрежений могут в зависимости от вида лежать гораздо выше. Однако кольцепоровые деревья (дуб, лианы — см. табл. 6.15) по значениям гидравлических проводимостей удивительно близки к идеальным капиллярам. Следует также учитывать работу подъема против силы тяжести (столб воды высотой в 1 м оказывает на свое основание давление 0,01 МПа = 0,1 бар), так что в совокупности величина пониженного давления должна составлять минимально -0,03 МПа • м-1. Для самых высоких видов деревьев — североамериканские секвойядендроны (Sequoiadendron) и австралийские эвкалипты (Eucalyptus), достигающие высоты 100 — 120 м, — создается, таким образом, отрицательное гидравлическое давление минимум от -3 до -4 МПа, позволяющее поднимать воду из корня до верхушки дерева. Правда, эти разрежения легко достигаются посредством возникновения матричных потенциалов вследствие испарения (см. рис. 6.32). Однако и осмотический потенциал в клетках листовой ткани достигает величин от -3 до -4 МПа (см. 6.3.2.1; рис. 6.28), и этого вполне достаточно для обеспечения подъема воды даже у самых высоких деревьев из корней до концов верхних побегов и в отсутствие транспирации.
Рис. 6.38. Зависимость необходимого градиента гидростатического давления (∆р) от диаметров капилляров при различных скоростях тока, по Хагену—Пуазейлю (см уравнение 6 32)
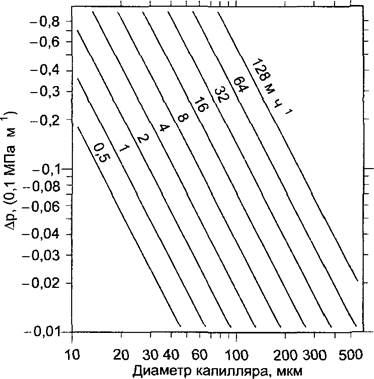
Таблица 6.15. Гидравлическая проводимость ксилемы различных растений (% от теоретических значений для идеальных капилляров с такими же диаметрами)
Растение |
Гидравлическая проводимость, % |
Виноград (лиана) Дуб (древесина корня) Пихта Береза (древесина корня) Тополь (древесина ствола) Различные травы и кустарники |
100 53-84 26-43 34,8 21,7 12-22 |
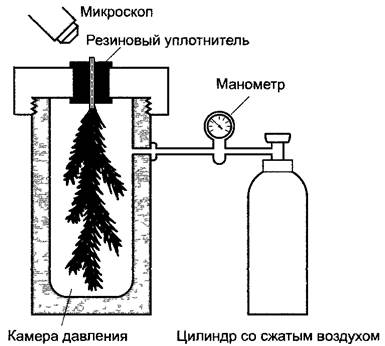
Экспериментальное доказательство наличия пониженного давления в ксилеме может быть получено с использованием камеры давления Сколандера. Для этого определяется избыточное давление, необходимое для того, чтобы у отрезанных частей растений на поверхностях среза снова показались мениски, которые при перерезании водных нитей втянутся внутрь водопроводящих сосудов под действием разрежения в ксилеме (рис. 6.39). Сначала таким способом определяется средний водный потенциал всего органа, заключенного в камере давления. Водный потенциал ксилемы в большинстве случаев приблизительно соответствует этому значению, поскольку осмотический потенциал ксилемного сока очень мал и поэтому водный потенциал ксилемного содержимого приблизительно равен отрицательному гидравлическому давлению водного столба в ксилеме, которая к тому же находится в очень близком контакте с прилегающими тканями.
Рис. 6.39. Камера давления для измерения отрицательного гидростатического давления в ксилеме частей растений
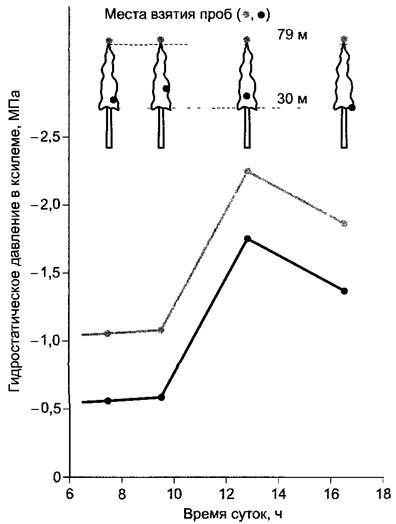
При помощи этого метода у высоких хвойных деревьев действительно было доказано наличие градиента давления в требуемом порядке величин (несколько более 0,01 МПа м-1) (рис. 6.40). Абсолютные значения пониженного давления показывали четкую суточную динамику с максимально отрицательными значениями во время самой интенсивной транспирации. Отсюда следует, что доставка воды не всегда «идет в ногу» с ее расходованием (см. 6.3.6 — Водный баланс). Суточные ритмы отрицательных гидростатических давлений приводят к тому, что у деревьев вовремя более интенсивной транспирации (в полдень) отчетливо уменьшаются диаметры по сравнению с условиями более слабой транспирации или при ее отсутствии (например, ночью).
Рис. 6.40. Градиенты давления в ксилеме лжетсуги (Pseudotsuga) в течение дня, измеренные с применением камеры давления. Кружками отмечены положения ветвей, каждый раз использованных для измерения
Столбы воды в проводящих путях могут противостоять «напряжению» лишь в том случае, если адгезия со стенками сосудов и когезия водных молекул выдерживают это усилие. «Напряжение», при котором когезия водных молекул нарушается, можно рассчитать теоретически или измерить опытным путем. Первое определение такого рода осуществили на естественной системе, изучая разрыв наполненных водой клеток кольца спорангия папоротника (см. 8.4; рис. 8.37). Он происходит при величинах между -22 МПа (насыщенный раствор сахарозы) и -36 МПа (насыщенный раствор поваренной соли). Чисто физическими методами создают еще более отрицательные величины (ниже -100 МПа). Следовательно, нет опасности, что при разрежениях, преобладающих в проводящих сосудах, когезия воды будет нарушена (когезионная теория проведения воды).
Опасность разрыва сплошных столбов воды вследствие натяжения состоит в гораздо большей степени в том, что в проводящих путях встречаются случаи газовой эмболии, когда при существующих в ксилеме условиях давления даже мельчайшие пузырьки газа занимают большие объемы (отсутствует когезия газовых молекул). С помощью чувствительных ультразвуковых детекторов можно регистрировать акустически звуковые волны, возникающие при «обрыве» столбов воды в стеблях интенсивно транспирирующих растений.
Лишь вопросом времени представляется это прежде всего для проводящих элементов с широкими просветами, когда они утратят способность функционировать в результате эмболии (в большинстве случаев необратимой). У кольцепоровых деревьев, например, у дуба, крупные трахеи, как правило, функционируют лишь в течение одного вегетационного периода, а к началу нового периода роста вся водопроводящая система должна быть заново создана камбием. Это одна из причин, почему у дубов весной так поздно раскрываются почки. Еще не выяснено, каким образом трахеи у лиан, имеющие крупные просветы, сохраняют способность функционировать долгие годы.
Состоящие из трахеид проводящие пути, например, в древесине голосеменных, гораздо менее подвержены эмболии. Если трахеида и выпадает в результате эмболии, то вследствие изменения давления торусы окаймленных пор сразу же необратимо закрывают проводящий элемент, изолируя его от соседних трахеид («принцип переборки», см. 3.2.4.2; рис. 2.75). Обратимое запирание имеет место в том случае, когда содержимое трахеид начинает замерзать и испытывает влияние давления, возникающего вследствие увеличения объема при образовании льда. В трахеидах, теперь герметично закрытых, достаточно замерзания малой части заполняющей воды, чтобы компенсировать существовавшее, возможно, до того разрежение и, таким образом, уменьшить образование газовых пузырьков. Каждое последующее образование льда дополнительно сдавливает оставшуюся жидкую фазу и удерживает газ в растворе, пока вся вода не замерзает. При таянии процессы протекают в противоположном направлении, так что и при возникновении пониженного давления пузырьки газа не образуются. Эта функция окаймленных пор, наряду с другими особенностями строения, обусловливает особую приспособленность голосеменных к заселению холодных областей. Показательно, что торусов в окаймленных порах не имеют лишь те голосеменные, которые не подвергаются опасности морозов (например, Суcas или палеозойские роды Саllixylon и Согdaites).
Живые клетки, соседствующие с проводящими путями, прежде всего с крупными трахеями (паратрахеальная паренхима), могут нести функцию защиты от проникновения пузырьков газа в проводящие элементы; остается неясным, способны ли они также устранять пузырьки, имеющиеся в проводящих путях.
Предполагают, что давление в ксилеме, возрастающее при отсутствии транспирации, содействует растворению пузырьков газа (растворимость газов в жидкостях увеличивается при возрастании давления). В этом может состоять еще одна функция корневого давления.
У больших деревьев ночью (при закрытых устьицах) движение воды в кроне термоэлектрическим методом не обнаруживается. Утром с началом устьичной транспирации начинается движение воды в периферических частях кроны, и затем вода распространяется вниз по стволу. Вечером транспирационный ток прекращается в той же последовательности: сначала в кроне и лишь после этого — в верхних частях ствола; часто ночью он не окончательно останавливается в основании ствола и в корне. Столь длительная работа этих органов требуется для полного восстановления резервов воды.
6.3.6. Водный баланс
Разность между поглощением воды и ее потерей называется водным балансом. Отрицательный водный баланс наблюдается в случае преобладания транспирации над поглощением воды, в обратном случае говорят о положительном водном балансе. При интенсивной дневной транспирации возможно возникновение отрицательного водного баланса, в то время как ночью этот дефицит снова выравнивается. В сухой период полного восстановления не происходит, так что баланс становится все более отрицательным. Вследствие этого все отрицательнее становятся осмотический потенциал и водный потенциал. Различные виды, а также различные экотипы растений внутри видовой принадлежности выдерживают различные степени и различную длительность такого дефицита, т.е. обладают различной засухоустойчивостью.
Водный баланс растения (или органа) часто выражают как процентный дефицит насыщения водой (ДНВ); он показывает, сколько воды требуется ткани для полного насыщения:
![]()
где WН — насыщающее содержание воды; WР — реальное содержание воды (об экологии водного обмена см. 13.5).