БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
14. ЭКОЛОГИЯ ПОПУЛЯЦИЙ И РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Эта глава посвящена развитию и сложению растительности. Растительное сообщество, существующее в определенном местообитании, в конечном счете является результатом сложных взаимодействий естественно-исторических и современных процессов с абиотической средой (климат и исходный субстрат, см. гл. 12 и 13), но его нельзя понять без учета процессов, протекающих внутри самого сообщества, и влияния внешних воздействий (рис. 14.1). Таким образом, друг с другом тесно переплетаются несколько уровней комплексности:
— популяции особей отдельного вида;
— виды растений;
— отдельные единицы растительности;
— мозаика единиц растительности в ландшафте;
— растительные формации;
— климатически обусловленные зоны растительности и высотные пояса.
Рис. 14.1. Схема становления растительных сообществ под влиянием ступенчато проявляющихся внешних воздействий или конкретных условий, а также внутренней динамики и их взаимодействия
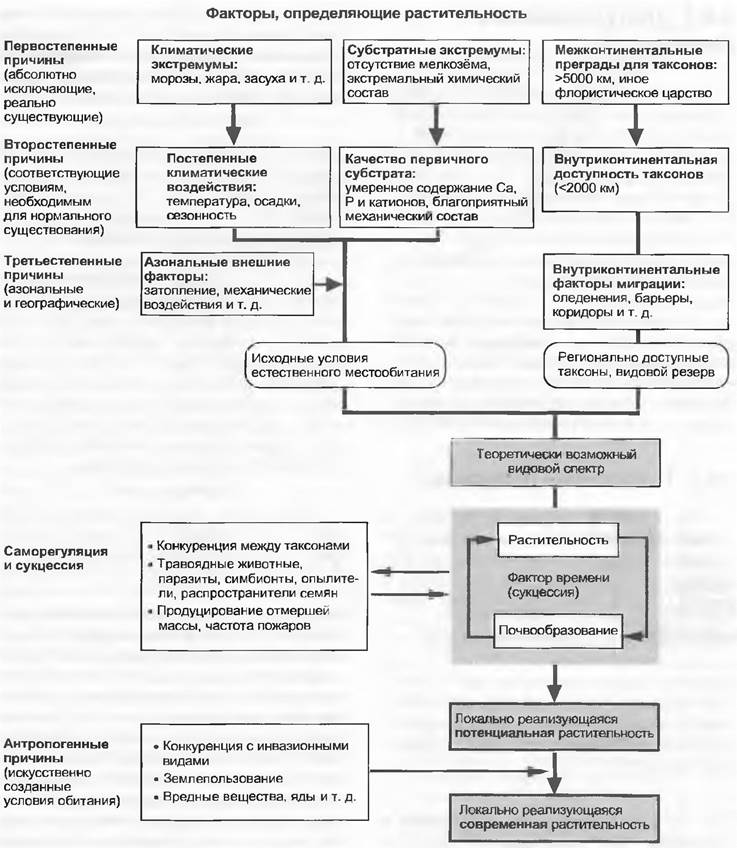
Соответственно, перечисленные элементы укладываются в большую эволюционно обусловленную систему сообществ — растительный покров Земли. От того факта, произрастает ли некий вид на определенном местообитании и может ли закрепиться там, зависит успешное существование его потомства. Поэтому исходным пунктом этой главы являются основы популяционной экологии, которые непосредственно соотносятся с гл. 10. Появление, пребывание и исчезновение видов в определенном жизненном пространстве как бы вливаются в более крупную площадь, занимаемую ареалом вида или группы видов, это тема раздела 14.2. В разделе 14.3 речь пойдет о наблюдаемом в определенное время локальном результате этих процессов — растительном сообществе.
14.1. Популяционная экология
Внешняя среда воздействует на судьбу индивида и тем самым на динамику популяций и их величину самыми разнообразными способами. Популяционная экология занимается регистрацией этой динамики и выяснением биотических и абиотических причин, которые ее обусловливают. Процессы, происходящие в рамках одного вида, в особенности генетическое (эволюционное) дальнейшее развитие популяций рассматривались в гл. 10. Вопросы биологии цветения и распространения диаспор — предмет раздела 11.2 (цветение; распространение плодов и семян). Здесь рассмотрим вопросы развития популяций и конкуренции, а также репродукционную экологию и стратегию размножения растений.
14.1.1. Развитие популяций
Как и для многих особей, наличие на данной плошали отдельного растения (N) в течение определенного времени (t) есть результат его рождения (В, англ. birth) и смерти (D, англ. death). В открытых системах существует еще возможность, что индивидумы, существующие на определенном базовом местообитании, переселились извне или ушли из него (импорт или экспорт диаспор). Это выражается определенным уравнением для изменения величины популяций в промежутке между моментами времени t и t + 1:
![]()
I и Е — обозначения для иммиграции и эмиграции, могут быть выражены как ∆М, или чистая миграция. В дальнейшем для упрощения понимания не будем учитывать миграцию. Поскольку растение изначально закреплено корнем, его мобильность (в отличие от большинства животных) близка к нулю. Для популяционной экологии растений этот факт имеет далеко идущие последствия. Захват пространства является окончательным (пространственно структурированные фитоценозы) и способен изменяться только путем репродукции, причем потомки и родительские особи часто произрастают скученно. Некоторые клонообразующие виды (см. 14.1.3) и свободноплавающие водные растения ограниченно мобильны. Изменение величины популяции![]() вытекает из уравнения
вытекает из уравнения
![]()
причем t обычно выражается в годах. Величина![]() может быть больше, меньше или равна единице. При
может быть больше, меньше или равна единице. При![]() величина популяции остается стабильной. Если
величина популяции остается стабильной. Если![]() в течение длительного времени больше 1, популяция экспоненциально увеличивается, что может быть описано экспоненциальной моделью роста. Примем определенную единицу времени (например, год), обозначим степень появления всходов как b, а степень отмирания как d, отсюда следует, что скорость, с которой происходит изменение числа индивидумов в эту единицу времени (скорость роста или сокращения), г = b — d или, выражая изменение числа индивидумов в единицу времени,
в течение длительного времени больше 1, популяция экспоненциально увеличивается, что может быть описано экспоненциальной моделью роста. Примем определенную единицу времени (например, год), обозначим степень появления всходов как b, а степень отмирания как d, отсюда следует, что скорость, с которой происходит изменение числа индивидумов в эту единицу времени (скорость роста или сокращения), г = b — d или, выражая изменение числа индивидумов в единицу времени,
![]()
Отсюда для определенного отрезка времени t, отсчитывая от нулевого момента, рост популяции можно выразить как
![]()
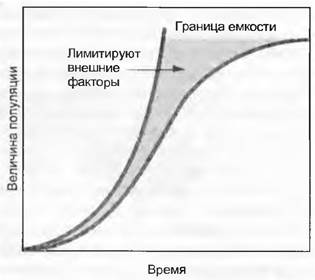
Причем г— абсолютный процент прироста (англ. intrinsic growth rate); t — длительность периода наблюдения; е = 2,718. Эта модель справедлива для популяций с перекрывающимися поколениями (в противоположность однолетникам, которые каждый год формируют новые популяции). Популяция индивидумов, подчиняющаяся данному правилу (тогда r остается постоянным), растет в геометрической прогрессии (рис. 14.2), это значит, что число ее индивидумов удваивается с постоянной скоростью. Росту препятствуют естественные ограничения (наличие питательных веществ или просто места), для которых установлено понятие максимальной емкости К (англ. carrying capacity), что означает максимальное число видов на единицу площади. В случае данной самой простой модели биомассы индивидумов не принимаются во внимание, хотя на самом деле они различаются. Степень роста популяции будет ослабевать благодаря фактору К - N/K до тех пор, пока при значении К = N не станет нулевой:
![]()
Это уравнение описывает сигмовидную модель роста (от греч. Σ, см. рис. 14.2). Она на самом деле намного ближе к реальности, чем геометрическая модель, поскольку принимаются во внимание пределы роста, но исходя из ряда упрощенных положений она наилучшим образом подходит для моновидовых клеточных культур в однородной среде.
Рис. 14.2. Кривые роста. Число индивидумов в популяции только при условии неограниченности пространства и ресурсов возрастает геометрически; сигмовидная кривая насыщается до граничных значений емкости систем
Для более высокоорганизованных организмов нереально предположить, что все особи постоянно размножаются с одной и той же скоростью. В действительности дочерние организмы (семена) продуцируются только на отдельном определенном жизненном отрезке, а число выживающих, способных к репродукции особей, получившихся из этих семенных популяций, в конечном счете ничтожное, у стабильных популяций теоретически это число должно быть равно количеству отмерших особей, которые достигли репродуктивного возраста, т. е. из преобладающей части семян никогда не разовьется репродуктивная особь. Таким образом, размер популяции определяется всеми фазами жизненного цикла (lifecycle) отдельно взятой особи, а не только продукцией диаспор, которая представляет собой лишь один этап репродукционного цикла. На каждой фазе существования имеются лимитирующие рост обстоятельства, которые действуют особенно сильно (в развитии популяции, это так называемая фаза бутылочного горлышка) и которыми в наибольшей степени объясняются факты произрастания и обилия видов. Данный аспект часто упускают, ориентируясь на большую часть экофизиологических исследований за поведением растущего растения, не всегда уяснив при этом, обусловливает ли этот жизненный этап преуспевание рассматриваемого вида.
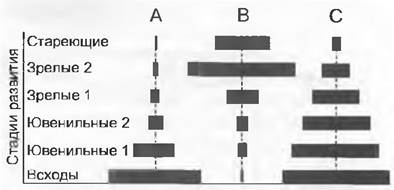
Популяция одного вида охватывает особи на всех индивидуальных стадиях развития и всех возрастных классов. Демография описывает количественное соотношение этих жизненных этапов и возрастной состав популяции, так называемую структуру популяции (рис. 14.3). Для этого необходимо определение возраста отдельных особей. У деревьев, произрастающих в регионах с сезонным климатом погоды, фактический возраст в годах можно установить по годичным кольцам (аналогично возрастной структуре популяций человека). Обычно для определения демографической структуры учитываются характерные стадии развития (число или процент особей, приходящиеся на каждую возрастную стадию). Для описания структуры популяций за неимением других сведений также служат данные по размерам отдельных особей (например, высота, диаметр, масса).
Рис. 14.3. Возрастная структура популяций, представленная в форме пирамид возрастов. Ширина горизонтальных широких полос отражает число особей (или их долю в общем числе особей всей популяции) определенных возрастных классов. Примеры гипотетические и символизируют: А — популяцию с аномально большим потомством, но с малым количеством старых особей, что может отражать либо начало экспансии, либо высокую смертность более старых особей; В — плохое воспроизведение (отсутствующие особи самых молодых классов возрастов) с повышенным риском вымирания; С — пропорциональную возрастную структуру с равномерно распределенным риском вымирания. У долгоживущих видов (деревьев) отсутствие на данный момент всходов и молодых растений не обязательно может быть интерпретировано как индикатор вымирания, поскольку многие виды размножаются периодически. Аналогично и для короткоживущих видов, у которых может быть неизвестно количество покоящихся, но жизнеспособных семян в семенном банке
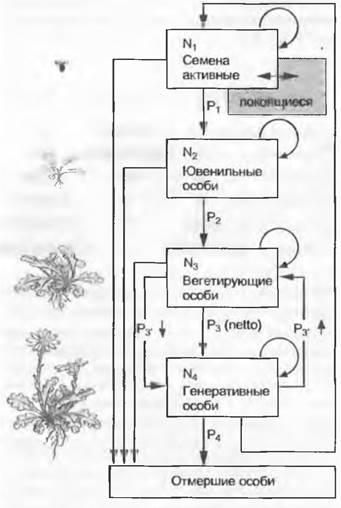
Увидеть динамику развития популяций можно путем повторного сравнения отдельных демографических описаний. Тогда можно оценить, с какой вероятностью особи переходят из одной жизненной фазы (или класса размера) в другую (рис. 14.4, 14.5). Вероятность перехода между отдельными жизненными фазами определяет форму демографической пирамиды популяции и ее рост. Таблицы вероятностей переживания отдельных возрастных фаз и фаз развития для видов называют «life table» — таблицами выживания.
Рис. 14.4. Жизненный цикл («история жизни») растения, состоящий из характерных отрезков, которые с определенной долей вероятности (между 0 и 1) переходят на следующие стадии. Эта вероятность перехода (Р) в большой степени зависит от конкретной жизненной стадии и внешней среды («просеивание» внешними факторами). N — количество индивидумов одного возрастного класса. Развитие между N3 и N4 может идти в обоих направлениях
Количественное соотношение растений отдельных стадий развития или возрастных классов и вероятности переходов, вытекающие из повторных сравнений, заносятся в таблицу (матрицу). Подобные матрицы переходов (англ. transition matrix) дают возможность смоделировать дальнейшее развитие популяции. При этом сблокированные группы особей одной возрастной фазы будут переходить в другую, подчиняясь вероятности, свойственной именно данному этапу перехода. В течение многих циклов это приводит к изменению во времени величины и структуры популяции. Поскольку каждая отдельно взятая вероятность перехода зависит от воздействий среды, а переходы между двумя жизненными фазами также ранжированы во времени и размыты в результате внутривидовых взаимодействий, такая модель сразу становится очень сложной. Если еще включить в нее межвидовые взаимодействия и менять условия внешней среды, она еще больше усложняется. По этим понятным причинам в большинство популяционных моделей воздействия внешней среды включаются в недостаточном объеме. Эмпирически полученные вероятности переходов включают действие всех внешних факторов в качестве «черного ящика». Кроме того, развитие популяций находится в тесной зависимости от саморегуляции (см. 14.1.2).
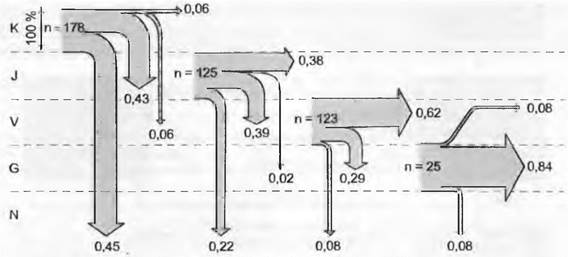
Рис. 14.5. Судьба одной разновозрастной исходной популяции Ranunculus acris на лугу, один год после первого измерения (n — изначальное число особей на 10 м2 каждого возрастного класса). Стрелки показывают, с какой вероятностью перехода (ширина стрелки и показатель в десятичных долях) особи оказываются в соответствующем возрастном классе (показаны, как «этажи»), К — всходы; J — ювенильные растения; V — сформировавшие вегетативные розетки; G — генеративные (цветущие) растения; N — более не существующие (отмершие) растения
Ценность такой модели наряду с отягощенным большим количеством неопределенностей прогнозированием дальнейшего развития популяции на основе достаточно постоянных вероятностей перехода заключается также в имитации возможного развития при изменяющихся вероятностях перехода («если предположить в дальнейшем»). Определенной один раз в полевых условиях демографической ситуации недостаточно, поскольку она не может прояснить динамики развития популяции, но она все же передает общую картину наличия молодых растений на данный момент и по ней можно заблаговременно определить, грозит ли популяции опасность вымирания (отсутствие или неудовлетворительное состояние подроста) или идет ли экспансия (инвазия) вида.
Судьба семян оказывается под влиянием разнообразных факторов, что хорошо иллюстрируется вероятностями переходов (рис. 14.6, 14.7). Основная часть особей по самым разным причинам теряется на том пути, который проходят семена от момента своего продуцирования (после чего они попадают в семенной банк в почве) до сформировавшейся популяции сеянцев.
Рис. 14.6. Судьба семенной популяции участка луга размером 10 м2. Схема иллюстрирует приемлемый, разумеется, гипотетический сценарий, так как в практике невозможно дать количественную оценку судьбы всех этих семян. Полная совокупность (семенной пул) могла бы быть определена на конкретный момент только приблизительно путем подсчета семян в выборочных почвенных пробах под микроскопом и последующего их проращивания
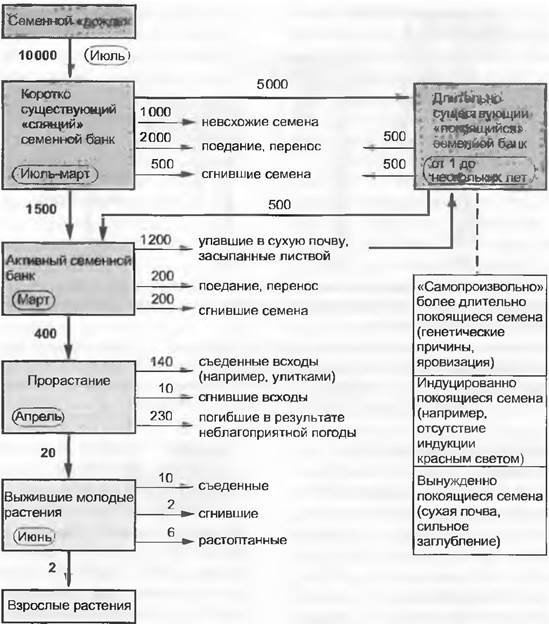
В примере с сорным видом растения, имеющим мелкие семена (см. рис. 14.6), конечная вероятность перехода семени в укоренившееся молодое растение составляет 0,0002, т. е. этот переход полностью совершают только 2 семени из 10000. Из 400 семян, которые к следующей весне будут готовы к прорастанию, большая часть погибнет из-за неблагоприятных погодных условий (например, иссушение почвы до того момента, как разовьются настоящие корни). Из оставшихся 150 сеянцев в этой модели 140 в первую же неделю будут съедены улитками. Каковы же были бы последствия для популяции, если только не было бы пожирателей улиток (например, появления одного такого охотника, как еж или крот) или если яйца улиток не поражались бы паразитами?
Поскольку растения имеют модулярное строение, каждый индивидум можно рассматривать как популяцию модулей (например, всех фитомеров, см. 4.2.1). Такие под
ходы очень успешно применяются при анализе клонообразующих растений, у которых особи, или раметы (англ. ramets) представляют различные возрастные группы генетически единого индивидума (англ. genets). Возрастная структура рамет отражает динамику роста клонов. Также листья одного растения или ветви деревьев могут рассматриваться как структурированные в возрастном отношении популяции. Как индивидумы листья будут «рождаться», «умирать» и проходить при этом определенные жизненные фазы. Результата изучения демографии листьев (например, жизненного цикла листьев пшеницы) существенны для продукционной биологии и многократно превосходят по достоверности результаты физиологических измерений во взрослых листьях. Без их знания данные по продуктивности (продуктивности фотосинтеза) не могут быть интерпретированы с точки зрения продукции (длительность жизни листьев, см. 13.6.3, 13.7.3). Кроме того, подобные данные можно получить без технического оснащения, путем маркировки и повторных измерений. Для большинства биотопов Земли они неизвестны, несмотря на их высокую экологическую значимость. Продуктивность таких естественных, несезонных злаковников, какие имеются в тропических горах, может быть определена только с применением листовой демографии.
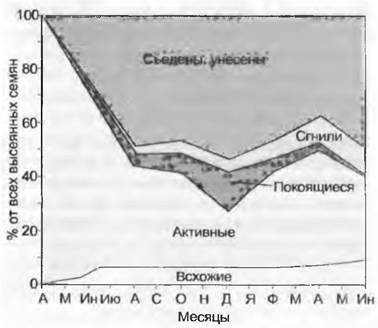
Рис. 14.7. Судьба семян лютика едкого (Ranunculus acris). На одном участке луга было посеяно, не считая естественного самосева, по 100 всхожих семян, на тестовых пробных площадках, после чего некоторые тестовые площадки раскапывались через регулярные промежутки времени, состав и качество семян анализировались под микроскопом
Бокс 14.1. Метапопуляция последствия фрагментации местообитания для выявления видов
Индивидумы одного вида редко распределены по пространству равномерно, но произрастают отдельными популяциями в подходящих для них местообитаниях, различными способами обмениваясь друг с другом диаспорами или пыльцой. Благодаря такой пространственной структуре динамика и генетическая структура популяций являются продуктом не только локальных условий, но и тех процессов, которые действуют на региональном уровне. Концепция метапопуляции принимает в расчет эту пространственную размерность(мета означает сверхпопуляцию. т. е. по сути более объемную). По С. А. Левину (1970), метапопуляция — популяция из субпопуляций, которые локально могут отмирать и опять возвращаться на место. При этом часть занятых местообитаний, которые свойственны рассматриваемому виду, является результатом процессов отмирания и колонизации. Подобным образом метапопуляция занимает структурированный ландшафт с многочисленными мелкими островками экотопов среди окружающего пространства с неподходящими для него условиями. Метапопуляция может существовать долгое время только при условии, что число вновь локально возникших субпопуляций превышает число локально отмерших. Это положение, само по себе банальное, позволяет установить связи динамики метапопуляции со структурой окружающей среды, в основном с величиной и изолированностью пригодных местообитаний. Благодаря этой идее в последние годы возник большой интерес к моделям метапопуляций, особенно в природоохранной области. Для изучения метапопуляций в наибольшей степени подходят растения в силу их прикрепленное к субстрату, выраженной пространственной структуры и ограниченной способности к расселению. До нашего времени проведены лишь немногие исследования конкретных метапопуляций, поскольку трудно определить такие важные параметры, как степень смертности и возникновения, а также происходящие миграции. При этом долгосрочное выживание многих видов может иметь столь же важное значение, как и обычная популяционная регуляция на локальном уровне.
14.1.2. Конкуренция
Субпопуляции, произошедшие от одних и тех же родительских растений, обозначаются обычно терминами, происходящими от названий древнеримских военных подразделений. В пределах каждой популяции представлены продуцируемые семена, взошедшие семена и взрослые молодые растения, все обозначенные одним и тем же термином — когорты.Когорты разных видов одного возраста иногда объединяют в легионы. Семенная когорта попадает на еще не заселенное место, и многие семена всходят одновременно на небольшом пространстве, формируется когорта проростков, внутри которой при увеличении размеров особей возникает проблема территории. В синхронных популяциях взаимная конкуренция за свет и почвенные ресурсы приводит в действие массированные процессы внутривидового отбора. Демографические процессы всегда тесно взаимосвязаны, в изначально возникшей инициальной популяции они находят отражение не только в смертности (что будет описано в следующем разделе), но и в плодовитости (фертильности). Обычно из каждой семенной когорты вырастают лишь немногие особи, часто далеко отстоящие друг от друга и попавшие в пустые пространства между особями своего и других видов (см. рис. 14.6). В результате повторения этого процесса во времени возникают популяции, структурированные по возрасту или размерам, в них конкурируют не только индивидумы одновозрастных когорт, но и появляются асинхронные популяции, находящиеся в комплексном взаимодействии с существующей растительностью. Однако основные трудности остаются у синхронных популяций, что лучше всего видно судя по обычным пожарам и катастрофическим нарушениям в инициальных сообществах на свежеобразованных субстратах или на землях сельскохозяйственного или лесохозяйственного использования.
Возьмем за исходную точку когорту семян (для наглядности — семян деревьев), которые собраны в семенном банке в почве, и синхронно возникшую из нее без конкуренции с другими видами представительную популяцию всходов. В высшей степени невероятно, что все эти сеянцы вырастут во взрослые деревья (для этого просто не хватит места) и что все они растут совершенно идентично (с одинаковой скоростью и одинаковой формы). Самая маленькая разница в размере семени и ходе развития (например, прорастание несколькими часами раньше) порождает сначала едва различимую разницу в размерах растения, которая потом быстро увеличивается (сопоставимо с «эффектом суммируемых процентов», рис. 14.8). Это внутривидовое неравенство — причина возникающих процессов естественного отбора за счет угнетения, называемого самоизреживанием (англ. self-thinning). Благодаря своему фундаментальному значению в биологии они стали предметом сотен публикаций, но до сих пор их механизм окончательно не прояснен.
Рис. 14.8. Внутривидовая конкуренция в синхронно высеянной монокультуре быстро усиливается за счет эффекта суммируемых процентов и ведет к неравномерному росту особей, который обусловливает процесс самоизреживания
Самоизреживание протекает удивительным образом закономерно или совершенно различным образом у каждого вида, но следуя повторяющемуся соотношению, «правилу самоизреживания -3/2» (минус три вторых), согласно которому густота особей с увеличением средней массы одной особи линейно снижается, подчиняясь зависимости двух логарифмических функций, а именно с уклоном -1,5 раза (рис. 14.9). В зрелых сообществах, которые достигли своей окончательной высоты (и постоянной итоговой массы), это соотношение выравнивается до -1 Подъем так называемой кривой самоизреживания, который происходит на самых поздних стадиях этого процесса, означает для густо посеянных, экранированных от света всходов трав возможность вырваться из-под гнета, так же, как и для лесных посадок или молодой поросли после естественного пожара. При этом изменяется точка пересечения с абсциссой (параллельное смещение).
Рис. 14.9. Самоизреживание в синхронно взошедшей монокультуре следует «правилу самоизреживания по -3/2»
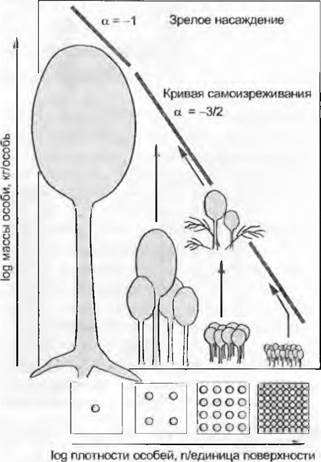
В лесном хозяйстве самоизреживание предупреждается своевременным прореживанием, в сельском хозяйстве — соответствующей подготовкой семенного материала.
Плотность колосоносных стеблей на поле не увеличивается после определенного граничного значения количества семенного материала. Более плотный посев дал бы только увеличение стерильных стеблей и снижение кущения на зерно (большее число побегов, но с мелкими колосьями). Говоря о постоянном конечном урожае, имеют в виду некое количество биомассы на определенную площадь, которое при данной степени плодородия почвы не может больше увеличиться. При более высокой плотности урожай зерна может вообще стремиться к нулю, хотя общая продуцируемая биомасса на единицу площади остается неизменно высокой. В спелом лесном насаждении биомасса на единицу площади может не зависеть от плотности, остающейся почти постоянной, т. е. годичная продукция биомассы только компенсирует опад (включая те особи, которые теряются в процессе самоизреживания).
Геометрия естественного высева (пространственная структура популяции) имеет определенное влияние на развитие сообщества. Плотное, примерно регулярное распределение семян (при заданной плотности расстояния между отдельными особями сходны) более содействует высокой урожайности и силе конкурентности в отношении сорняков, чем рядовой посев (с неодинаковыми расстояниями между особями и между рядами; no J. Weiner). Это также существенный экологический фактор, и возникает вопрос, почему так часто встречается групповое произрастание видов там, где межвидовая конкуренция сильно повышается. На такие вопросы можно дать ответ, если известны все действующие в процессе эволюции факторы риска. При групповом произрастании (как и в косяках рыб) увеличиваются шансы на индивидуальное выживание при выборочном прессе со стороны травоядных, даже если ценой этого будет более медленный рост особи. Групповое произрастание является неизбежным следствием выборочного распространения источников диаспор, стохастического размещения и гетерогенно (англ. patchy) распределенных условий существования.
Высоту подъема -3/2 обычно объясняют исходя из того факта, что растения прикреплены к субстрату и при этом поверхность, на которой разрастается популяция, фиксирована, а высота морфологически и статически лимитирована, чем также ограничен максимальный возможный объем, находящийся в распоряжении одной особи. Биомасса же одной особи занимает место, равное ее объему. Как в ящике для кубиков, в данном объеме могут поместиться только либо много маленьких, либо немного больших кубиков-растений, отношение следует показателю одной кубической и одной квадратичной функции длины ребра (как раз в логарифмической форме 3/2). Но за конкретным положением «кривой самоизреживания» должна также скрываться такая биологическая «константа», как потребность в свете, необходимом для фотосинтеза, а также автотрофно-гетеротрофные соотношения (листовая биомасса/нелистовая биомасса) в самом растительном организме. Лес из толстых и плотно стоящих стволов, но с небольшим количеством листьев на макушках невозможно себе представить. Попытки экспериментальным образом изменить «кривую самоизреживания» показали ее выполаживание при сильном затенении.
Пример так называемой асимметричной конкуренции, самоизреживания популяции, зависящего от ее плотности, — угнетенность и последующее отмирание отстающих в росте особей в пользу более высоких, которые получают большее количество солнечного света. Конкуренция называется асимметричной, поскольку возможности использовать световые ресурсы распределяются между индивидуальными растениями асимметрично (свет — векторно направленный ресурс) и соревнование между ними приводит к сильной асимметрии в индивидуальных размерах. Напротив, используемый растениями запас питательных веществ в почве распределен скорее диффузно, у корней (по крайней мере, теоретически) имеются одинаковые возможности использовать питательные вещества, поэтому данную ситуацию можно назвать симметричной конкуренцией. В реальности между этими двумя крайними случаями есть множество переходных. Взаимоотношения наземных побегов, как правило, в большей степени асимметричны, чем корневых систем. Для интерпретации результатов конкуренции всегда требуется анализ процессов как в надземной, так и в подземной сфере (например, удаление соседних особей, пересадка, «садовый» или «горшечный» эксперимент, на рис. 14.10).
Рис. 14.10. Надземная и подземная конкуренция за ресурсы на примере вьюнка (Ipomoea tricolor). Светлые столбцы демонстрируют действие на биомассу одного растения (среднюю); больше влияет подземная конкуренция, меньше — надземная. Темные столбцы показывают вариабельность (асимметрию) биомассы, обусловленную различными условиями роста в каждом опыте: больше влияет надземная конкуренция, меньше — подземная
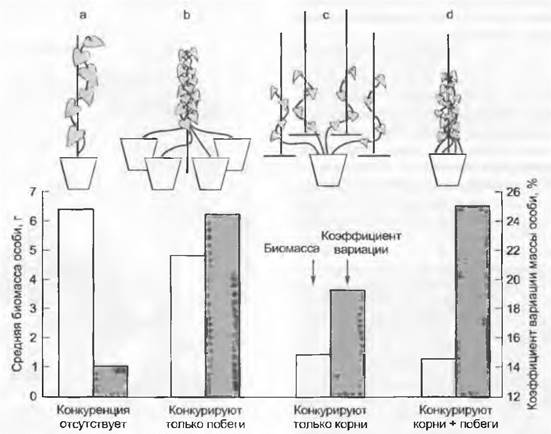
Представленный на рис. 14.10 опыт иллюстрирует различающееся значение конкуренции побегов и корней для вьюнка. Значение биомассы одного растения отражает «весомость» обоюдных ограничений; вариабельность этих значений показывает, насколько воздействие асимметрично. Высокая вариабельность означает большие различия между самым мелким и самым крупным растением каждой группы, указывая этим на тенденции угнетения. В то время как наземная конкуренция (ср. а и b) мало снижает биомассу, она усиливает асимметрию в индивидуальных размерах растений. Только подземная конкуренция (с) ведет к резкому уменьшения биомассы (что и не может быть иначе в силу уменьшения пространства, используемого корнями), увеличение коэффициента вариации между биомассой особей с 14 % почти до 19% также невелико и статистически несущественно. Совместное действие обоих типов (d) ведет к дальнейшему снижению индивидуальной биомассы, а вариабельность возрастает до 25%, т. e. до того же уровня, как и при одной только конкуренции побегов, причем биомасса составляет только 1/5 от последнего случая. Это означает, что потери биомассы в данном эксперименте в первую очередь происходят из-за корневой конкуренции, но асимметрия биомасс одного растения обусловлена преимущественно конкуренцией побегов.
В случае длительного существования такой конкурентной ситуации всегда происходят подавление и последующее отмирание более слабых индивидумов или видов. Почему же тогда вообще имеются разнообразные популяции и многовидовые растительные сообщества? Это одна из центральных проблем изучения сосуществования видов и биоразнообразия (см. 14.2.4).
Многократно утверждалось (см. 14.2.4.1), что длительное сосуществование видов или генотипов в пределах одного вида возможно только тогда, когда их потребность в ресурсах в качественном отношении неодинакова, поэтому они хотя бы частично уклоняются от конкуренции (функциональная дифференциация ниш, по Г. Ф. Гаузе). Отсутствие дифференциации ниш приводит к конкурентному исключению (англ. competitive exclusion). Если ограничиться такими классическими ресурсами, как питательные вещества, вода и свет, то концепция ниш в настоящее время потеряла значение, поскольку большая часть растений на одном и том же местообитании нуждается в одних и тех же ресурсах. Определенная дифференциация ниш, однако, все же возможна путем пространственной и временной дифференциации жизненной активности (использование различных почвенных горизонтов, позиции в сообществе, разного времени года). В расширенную концепцию ниш включают также устойчивость к патогенным микроорганизмам и травоядным животным или даже дифференцированные мутуалистические связи с микоризными грибами и опылителями, тогда понятие «ниши» становится синонимом значения суммы всех свойств растения.
Было бы нетрудно приписать отсутствующее зачастую доказательство функциональной дифференциации ниш просто недостаточной точности и тщательности аналитики (самого определения ниш). Тут новые решающие возможности открывает математическое моделирование популяций и сообществ видов, в которое включаются нестабильные условия внешней среды. Теоретическая модель имеет здесь огромное преимущество, так как она не лимитирована временем, что всегда свойственно эксперименту. Смоделируем на компьютере классическую конкурентную ситуацию (например, вид растения, который в борьбе за свет всегда растет быстрее, чем остальные виды), тогда вывод будет таким, что из двух видов один всегда остается лишним. Добавим элемент нарушения, например, постоянное удаление 50% особей каждой популяции, тогда вопрос о том, какой из двух видов одержит победу, будет решаться дольше, но в конце концов из них все же останется один. Поставим в условие вместо двух шесть видов, произрастающих вместе, а в качестве нарушения — соразмерное изъятие особей на нерегулярном расстоянии друг от друга, тогда они на какое-то время останутся неограниченно сосуществующими, поскольку речь идет о видах, рост популяций которых происходит относительно медленно, и поскольку нарушение происходит не часто.
Хотя математическая модель не может воспроизвести всю неоднородность реального мира, имитация показывает, что виды могут сосуществовать и без дифференциации ниш, если они испытывают постоянные нарушения. Нарушение в комбинации со специфическими для каждого вида реакциями на это нарушение может обеспечивать сосуществование видов со значительно перекрывающимися экологическими нишами (см. 14.2.4.1) Растительные сообщества всегда испытывают какие-то нарушения: естественные злаковники стравливаются, средиземноморские сухие кустарники выгорают через определенные промежутки времени, девственные леса переживают постоянное изреживание и смыкание сообществ из-за падающих деревьев. Вызванная этим так называемая динамика лесных окон в значительной степени определяет высокое биоразнообразие тропических лесов (см. 14.3.1).
14.1.3. Экология возобновления
В этом разделе речь идет о различных жизненных стратегиях растений, выработанных ими для успеха воспроизведения (эволюционная биология, биология цветения и распространение диаспор, см. гл. 10 и раздел 11.2, семенные растения).
Обеспечение сохранности и дальнейшего развития своего генома в пространстве в течение времени — жизненная функция, превалирующая над всеми другими. Для ее осуществления имеются различные пути, или жизненные стратегии (англ. life strategies или life history strategies). Какие из них имеют успех, зависит в сильной степени от условий окружающей среды и сложившейся конкурентной ситуации. Основная проблема, которая стоит перед каждым растением, каким образом (и в какой временной последовательности) распределить ассимиляты между процессами размножения и вегетативного роста (англ. reproductive allocation). Если они вкладывают их в один процесс, то они не могут в той же мере одновременно снабдить другой; говорят о попеременном использовании, а также о компромиссе при условии ущерба для соответствующего альтернативного процесса. Эта инвестиционная стратегия тесно связана с длительностью жизни растения и с течением его жизненного цикла (англ. life cycle).
Некоторые растения могут завершить свой полный жизненный цикл за 6 недель (например, Arabidopsis thafiana), другим до достижения репродуктивной зрелости требуется от I до 3 лет (многие корневищные травянистые растения), после чего они отмирают, а некоторые виды деревьев могут оставаться в репродуктивной стадии более 2000 лет (Sequoiadendrongiganteum, Cryptomeriajapónica). Классическое подразделение растений на однолетние, двулетние и многолетние виды для такого континуума не будет справедливым. Некоторые однолетние проходят за один год несколько жизненных циклов, другие плодоносят только один раз в жизни (так называемые монокарпики или гапаксантные виды, см. 10.1.3.4), у них до первого (и в то же время последнего) цветения проходит от 20 до 30 лет, после чего они истощаются и отмирают (например, Agave americana).
Привести доказательства того, что именно и сколько растение вкладывает в репродукционные процессы, очень трудно. Нет четких границ между двумя положениями:
1) к ним относится только масса семян;
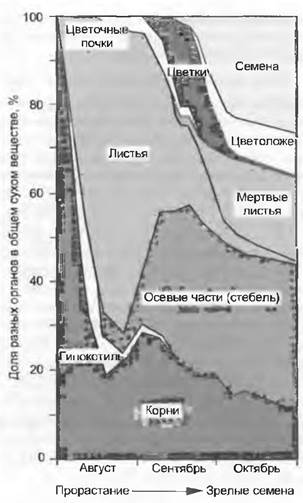
2) для обеспечения потомства служит вся продуцируемая растением биомасса. Обычно в «затраты на воспроизводство» включают все метаболические расходы, относящиеся к соцветию вместе с его осевыми частями, нектару и пыльце, плодам и семенам. Количественно это едва ли можно оценить, поэтому на практике за размеры часто принимают только общую массу плодов или семян, хотя она представляет собой лишь часть действительных затрат на воспроизводство (рис. 14.11). У некоторых короткоживущих травянистых растений и хлебных злаков (эго называют индексом урожая или harvest-индексом) биомасса диаспор составляет около 50% от общей продуцируемой биомассы. У многолетних растений это значение может падать до 1 % или вообще быть нулевым долгое время жизни.
Рис. 14.11. Доля репродуктивных структур в общей биомассе жизненного цикла крестовника обыкновенного (Senecio vulgaris)
На основании этого выделяют два противоположных типа жизненных стратегий: r-стратегию и К-стратегию (рис. 14.12). У растений, относящихся к r-стратегии, быстрое созревание многочисленных семян превалируют над продуцированием других органов и общей длительностью жизни. Это пионерные растения сильно нарушенных местообитаний, например, рудеральные растения (см. 12.5.1.3), а также виды, свойственные ранним стадиям сукцессий (см. L4.3.2). r-Стратеги вполне приспособлены к высокой степени риска смертности. Для К-стратегов более высокий приоритет имеют вегетативный рост и устойчивость (обеспечение длительности жизни). Они занимают однажды завоеванное пространство на как можно более длительное время, достигая этой цели за счет в целом консервативной (с минимальным риском гибели) стратегии роста и развития. По типологии Грамма (J.P.Grime) — это «конкуренты» (competitors), относящиеся к поздним стадиям сукцессий (см. 12.5.1.3). Большинство видов растений относится к промежуточным типам между этими двумя крайними стратегиями.
Рис. 14.12. На разных фазах сукцессионного процесса доминируют растения, относящиеся к разным жизненным и репродукционным стратегиям. Древесные растения: А — молодые заросли распространяемых ветром ив и тополей на щебнистой отмели, В — 300-летний коренной хвойный лес (Pseudotsuga menziesii = дугласия, Орегон). Травянистые растения. С — рудеральное сообщество на намывной поверхности; D — зрелое климаксовое горно-луговое сообщество тысячелетнего возраста (Caricetum curvulae, 2 500 м, Западные Альпы). На А и С изображены виды с быстрым размножением и высокой семенной продукцией, на В и D — растения с медленным вегетативным ростом, на D — растения с клоновым расселением

Бросается в глаза тот факт, что свойства диаспор характерным образом корреспондируют этим стратегиям. Большей частью r-стратегам свойственны многочисленные мелкие, «дешевые» семена, часто снабженные приспособлениями для распространения на дальние расстояния, обычно они способны к длительному нахождению в покоящемся состоянии (затягивание прорастания, крупные покоящиеся семенные банки). У однолетних пионерных растений семена могут сохранять всхожесть более 100 лет (сообщение о всхожих семенах из раскопок возрастом 1 600 лет — это экстремальный случай). Известно также, что пустынные так называемые эфемеры сохраняются в почве в виде семян много лет, и только при очень редко случающихся здесь сильных дождях пустыня неожиданно становится морем цветов.
К-стратеги обнаруживают тенденцию к продуцированию менее многочисленных, но более тяжелых семян, богатых запасными веществами. Последнее объясняется тем фактом, что на более поздних стадиях сукцессии прорастающие семена испытывают сильный лимит света. Семя должно нести в себе все то количество ресурсов, которое необходимо проростку, чтобы в достаточной мере углубиться в почву и существовать в условиях светового дефицита до тех пор, пока молодое растение не начнет фотосинтезировать. Максимальный период покоя семян у таких видов редко длится более 2 лет. Как правило, семена этих видов всходят после короткого периода покоя в следующем сезоне, благоприятном для прорастания (в умеренной зоне следующей весной или самое позднее через одну весну). Вместо семенного банка у К-стратегов часто образуется банк проростков, которые могут долго выжидать, пока наличие свободного пространства нс обеспечит им возможность дальнейшего развития (это типично для девственных влажно-тропических лесов).
Малолетние растения в большинстве случаев бывают мелкими, а многолетние часто крупными, из чего следует, что существует корреляция между средним размером растения и средним размером семени, хотя это правильно не во всех случаях (табл. 14.1). Самые легкие семена, впрочем, у орхидных — примерно 1 мк, а самые тяжелые у сейшельской пальмы Lodoicea maldivica (= L. sechellarum = L. callypige) — 18 — 27 кг. Так как семена орхидей не могут прорастать без симбиоза с грибами, нельзя сказать, что в данном случае мелкий размер семян связан с рудеральным характером. Масса семян — очень консервативный признак. При неблагоприятных условиях среды редуцируется в первую очередь число семян, но не их размеры. У травянистых растений гумидных областей средний размер семени, например, не изменяется при подъеме в более высокие горные пояса, в то время как среднее число семян у одной особи падает. Примечательно, что столь малое варьирование массы семян Сеratonia siliqua (рожковое дерево) привело даже к тому, что масса одного его семени стала мерой массы (карат).
Таблица 14.1. Масса семени коррелирует с размером растения
Средняя масса семени, мг |
||
Жизненная форма |
Великобритания |
В целом по миру |
Травянистые растения |
2 |
7 |
Кустарники |
85 |
69 |
Деревья |
653 |
328 |
Следствием различий отдельных диаспор является то, что у К-стратегов часто продуцируются диаспоры, привлекающие травоядных животных. Если бы таких диаспор постоянно образовывалось мало, животные могли бы приноровиться к этому и препятствовать размножению. Поэтому у наиболее выраженных К-стратегов наблюдается тенденция к длинным паузам в размножении, следующим за так называемыми урожайными годами, в которые имеющимся популяциям травоядных пищи более чем достаточно (например, у бука, дуба, многих хвойных пород). Это требует наличия специального бюджетного резерва питательных веществ. Альтернативным для них является вырабатывание в семенах ядовитых веществ или образование таких плодов, как ягоды или костянки, содержащиеся в которых семена, распространяются животными (часто после прохождения через желудочно-кишечный тракт).
У бука урожайные годы повторяются через каждые 6 — 7 лет. Они часто соответствуют более слабому росту годовых колец (иногда до 2 лет), что является доказательством конкуренции между процессами размножения и вегетативным ростом внутри одного индивидума. У хвойных, такое сокращение наблюдалось только в годы массовых плодоношений. Крупноплодные дубы, растущие в естественном густом лесу, продуцируют даже в хорошие годы обычно не более 2 000 желудей на одном дереве в год. У более рудеральных видов деревьев, таких, как береза и сосна обыкновенная, напротив, семена в меньшей степени представляют собой кормовую базу и число их очень мелких семян может достигать на одном дереве от 50000 до 300000. Для травянистой наперстянки (Digitalis purpurea) определено число семян на одном растении — 0,5 млн.
У растений в гораздо большей степени, чем у животных, распространены размножение и распространение путем клонального разрастания в обход рисков, с которыми связано половое размножение (см. 10.1.3.3). Многие травянистые растения в зависимости от конкретных условий жизни могут использовать либо вегетативное, либо генеративное размножение, либо оба одновременно. Значение вегетативного распространения особенно усиливается в том случае, когда условия существования становятся неблагоприятными.
Для вегетативного распространения служат столоны, корневища, дочерние луковицы, корневые клубни, отдельные участки побегов, снабженные придаточными корнями участки ползучих побегов или корневые отпрыски. Диаспоры, сопоставимые клональным единицам расселения, — выводковые почки (луковички, см. рис. 4.31), а также клональные семена, образовавшиеся в результате вторичного бесполого размножения, варианта агамоспермии (апомиксис, см 10.1.3.3), например, у Taraxacum officinale. Вегетативно размножаются большинство высокогорных растений, растений дюн, очень успешно — многие растения семиаридных областей (например, Larrea îridentata, креозотовый куст полупустынь Нового Света), а также растения, обитающие в зонах периодического затопления (Salixy Hippophae), и лаже лесные деревья (Popular, многие виды Ficus). Успешно чувствуют себя на часто выкашиваемых газонах такие способные к вегетативному размножению растения, как Bellis ре tenuis. Trifolium repens, также образуют клоны почти все многолетние травянистые однодольные (прежде всего злаки, бамбук, луковичные и корневищные растения). Именно благодаря выраженной тенденции к вегетативному распространению многие пашенные сорняки так трудно истребимы, той же стратегии придерживаются и многие широко распространенные рудеральные виды (например, Solidago canadensis, Epilobmm angustifolium).Все мхи, хвощи и лишайники, как и многие папоротники, разрастаются вегетативно. Лишь немногие многолетники не пользуются вегетативным размножением как альтернативой семенному, кроме (за редкими исключениями) деревьев. Именно по этой причине популяционно-биологические концепции, принятые в зоологии, могут быть перенесены на растения весьма ограниченно.
Почему же вегетативный (клональный) альтернативный тип размножения играет такую большую роль? Собственно говоря, он представляет собой «торможение» эволюционного процесса. Он очень выгоден в том случае, когда успех вида в меньшей степени зависит от числа генеративных особей, чем от возможности длительного выживания. Вегетативное распространение дает возможность виду достичь чрезвычайного пространственного доминирования без протекающих с риском процессов закрепления проростков. Особенно успешные генотипы могут «законсервироваться», а затем продолжиться, как это осуществляется у покрытосеменных в крайней форме агамоспермного апомиксиса (см. 10.1.3.3) и у криптогамных, например, лишайников, чьи вегетативные части, служащие для распространения, могут пересекать сколь угодно широкие пространства. Это можно сравнить с вегетативным воспроизводством однажды удачно полученных генотипов, применяемым в сельском хозяйстве для сортов фруктовых деревьев, винограда или цветов (улучшение породы посредством прививок, размножение черенками, см. 7.3.3).
Вегетативно размножающиеся растения при конкуренции с другими могут выбрать любой из двух путей. Распространение в стороны от себя может идти мелкими шагами, но тесным фронтом, как у растений, образующих клоны в форме гнезд или гирлянд, для чего применяется стратегия фаланги (аналог античной наступательной тактики сплошной стеной). Чаще встречается последовательное внедрение в чуждые популяции путем «разведчиков», так называемая партизанская стратегия. Эта вторая форма распространения способствует более быстрому завоеванию пространства и с помощью пробных тестирований окружающей среды позволяет быстрее отыскать благоприятные микроэкотопы, на которых клональные модули могут укорениться. Можно привести как пример ползучие виды рода Portulaca, или Trifolium repens, у которых такие побеги ориентируются в сторону поверхностей, отражающих красный свет (это указание на «зеленые» территории, уже занятые другими растениями). Многие виды могут в зависимости от конкретной ситуации применять обе стратегии более быстрый (но рискованный) выигрыш пространства и затем утверждение новых «бастионов» вплоть до подавляющего локального доминирования (рис. 14.13).
Рис. 14.13. Клональное распространение по образцу «фаланг». Клоны Festuca orthophylla столетнего возраста «вспахивают» сплошным фронтом высокогорные плато Анд в северо-западной части Аргентины (4 250 м, Кумбре Кальчагуа)

Единая система клонов может так перераспределить ресурсы внутри себя, что обеспечивается выживание модулей, периодически находящихся в невыгодном положении, а «форпосты», которые в особой степени обеспечивают успех занятия территории (например, незанятые места в насаждениях), могут быстро перетягивать в свою сторону минеральные вещества, что можно доказать, применив метод маркировки изотопами. Используя вегетативное распространение, растения достигают высокой степени подвижности и могут лучше использовать гетерогенно распределенный в пространстве запас ресурсов.
Системы клонов могут достигать чрезвычайно большого возраста и потенциально вообще бессмертны. Однако растительность, сложенная преимущественно клонами, вовсе не обязательно генетически однообразна. Несмотря на возможность соматических мутаций у части модулей внутри клонов, не раз было установлено, что такие доминирующие территориально клонообразующие растения, как тростниковые заросли или альпийские осоковые лужайки, обнаруживают удивительное генетическое разнообразие, что указывает на разнообразие исходных особей в начальной фазе заселения поверхности. Разные клоны (генеты) могут переплетаться друг с другом, при этом генетически различающиеся группы побегов (группы рамет) могут располагаться рядом. Используя генетические маркеры, стало возможным картирование таких клонов альпийской осоки (Сагех сurvula) и рассчитать возраст клона, оказавшийся более чем тысячелетним, исходя из его размеров и известной радиальной скорости разрастания.
Цель этого раздела — объяснить то, что популяции полностью охватывают все стадии развития особей, а не только те, когда на определенном жизненном отрезке растение крупное и бросается в глаза. Именно учет растений, находящихся на незаметных фазах развития, не раз позволял определить, является ли популяция растущей или сокращающейся. При этом динамика развития популяций зависит от возможных ресурсов и вместе с тем от плотности особей и конкурентных отношений и решающим образом контролируется нарушениями. Эволюционная реакция на частоту нарушений — скорость роста вместе с кратковременностью существования и высокой семенной продукцией. В стабильном жизненном пространстве реакция противоположна — это большая длительность жизни и сравнительно малый вклад в процессы размножения. Благодаря своему модулярному строению многие растения могут сделать ставку на вегетативный (клональный) тип размножения и расселения, исключив при этом наиболее чувствительные ранние фазы жизненного цикла. Эти процессы определяют не только локальный успех популяции, но и ее способность распространяться на большие площади, что и будет темой следующего раздела.