МЕДИЧНА БІОЛОГІЯ, АНАТОМІЯ, ФІЗІОЛОГІЯ ТА ПАТОЛОГІЯ ЛЮДИНИ - Я.І.Федонюк 2010
БІОЛОГІЯ
РОЗДІЛ 1. БІОЛОГІЧНІ ОСНОВИ ЖИТТЄДІЯЛЬНОСТІ ЛЮДИНИ
1.3. МОЛЕКУЛЯРНО-ГЕНЕТИЧНИЙ І КЛІТИННИЙ РІВНІ ОРГАНІЗАЦІЇ ЖИТТЯ
1.3.3. Спадковий апарат еукаріотичних клітин і його функціонування - молекулярний рівень
Реплікація ДНК
Реплікація (син: самоподвоєння, самовідтворення, авторепродукція молекул ДНК) - синтез ДНК на матриці ДНК, який відбувається в S-періоді інтерфази; забезпечує точне копіювання генетичної інформації. Здатність до самоподвоєння - унікальна властивість ДНК, яка притаманна лише ДНК і непритаманна ніякій іншій хімічній речовині. Схему реплікації первинно запропонували Дж.Уотсон і Ф.Крік (рис. 1.40). За цією схемою, біспіраль материнської молекули ДНК розділяється на два ланцюги і кожен з них слугує матрицею для синтезу за принципом комплементарності нового ланцюга. У результаті з однієї материнської молекули ДНК утворюються дві дочірні, які є точними копіями материнської. Оскільки в кожній новоутвореній дочірній молекулі один ланцюг старий (материнський), а інший - новий, такий механізм реплікації одержав назву напівконсервативного. Процес забезпечується координованою роботою ряду ферментів. Синтез ДНК каталізує фермент ДНК- полімераза. Існує декілька типів ДНК-полімераз. Фермент ДНК-полімеразу вперше в 1957 р. виділив з бактерії кишкової палички (Escherichia coli) А.Корнберг. З'ясувалося, що ДНК-полімераза не здатна почати синтез ланцюгів ДНК de novo, тому для її активності необхідна наявність у системі вже існуючої ДНК (так званої ДНК-затравки), тобто ланцюга ДНК, що має вільний У-ОН кінець. ДНК-полімераза забезпечує приєднання до 3-ОН кінця ДНК-затравки дезокси-рибонуклеотидів. Наявність у цій системі ДНК-матриці визначає, які саме нуклеотиди повинні приєднатися до ДНК-затравки. За наявності ДНК-затравки і ДНК-матриці ДНК-полімераза забезпечує синтез дочірнього ланцюга ДНК, комплементарного одному з ланцюгів материнської ДНК. У системі in vivo роль ДНК-затравки виконує РНК-затравка (РНК- праймер).

Рис. 1.40. Реплікація ДНК за схемою Дж.Уотсона і Ф.Кріка: 1,2- материнські ланцюги ДНК; 1a, 2а - дочірні ланцюги ДНК.
Реплікація починається в точці огі (англ. origin - початок), де відбувається розкручування біспіралі ДНК, локальне розведення її ланцюгів, стабілізація однониткових ДНК, поява з двох розведених ланцюгів Y-подібної структури (реплікативної вилки), синтез РНК-праймера. Синтез нових ланцюгів відбувається лише в напрямку 5' →3'. Ініціації синтезу передує утворення РНК-затравок (РНК- праймерів), до 3'-ОН кінця яких фермент ДНК-полімераза приєднує дезоксинуклеозидмонофосфати, що утворюють новий ланцюг. Синтез РНК-праймерів каталізує фермент РНК-полімераза - праймаза. Оскільки ланцюги ДНК антипаралельні (5→3 і 3→5) а ДНК-полімераза приєднує вільні нуклеотиди лише до З'-ОН кінця, синтез двох дочірніх ланцюгів здійснюється за різними механізмами. Один ланцюг лідируючий) синтезується безперервно від РНК- праймера в напрямку руху реплікативної вилки, а другий (відстаючий) - переривчасто з утворенням коротких фрагментів, так званих фрагментів Оказакі (Р.Оказакі - японський учений, який вперше їх відкрив), у напрямку, протилежному руху реплікативної вилки (рис. 1.41). Кожен з фрагментів починається з відповідного РНК-праймера. Ці короткі ділянки новосинтезованого полінуклеотидного ланцюга пізніше з'єднуються між собою ферментом ДНК-лігазою в один ланцюг. Після завершення реплікації РНК-праймери видаляються, а на це місце вбудовуються нуклеотиди, комплементарні ДНК-матриці.
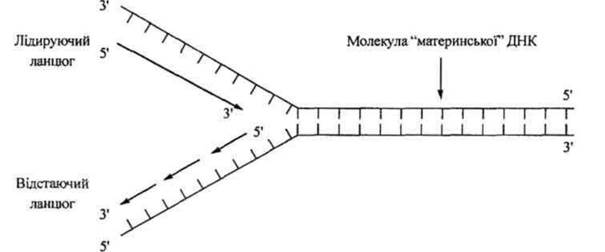
Рис. 1.41. Реплікація ДНК за схемою Р.Оказакі.
Ділянка ДНК, яка знаходиться під контролем однієї точки огі, називається одиницею реплікації (репліконом). У ДНК прокаріотів - одна точка огі і вся молекула подвоюється як один реплікон. У ДНК еукаріотів - багато точок огі і відповідно багато репліконів, що є необхідною умовою для того, щоб велетенські молекули ДНК еукаріотів встигли подвоїтися за період одного клітинного циклу.