Фізіологія людини - Вільям Ф. Ґанонґ 2002
Ендокринна система, метаболізм і репродуктивна функція
Гормональне регулювання метаболізму кальцію та фізіологія кістки
Вітамін D та гідроксихолекальцифероли
Хімія
Активне транспортування Са2+ та РО43 з кишки збільшується завдяки метаболіту вітаміну D. Термін вітамін D використовують для позначення групи близьких стеринів, утворених під дією ультрафіолетового світла на деякі провітаміни (рис. 21-7). Вітамін D3, який ще називають холекальциферолом, виникає в шкірі ссавців з 7-дегідрохолестеролу під дією сонячного світла. Реакція полягає у швидкому утворенні провітаміну D3, який згодом повільніше перетворюється у вітамін D3 (холекальциферол). Вітамін D3 та його гідроксильовані похідні транспортуються в плазмі зв’язано з глобуліном вітамін D-зв’язувальним білком (DBP - від англ. vitamin D-binding protein), який також відомий як GC-БІЛОК і зв’язує G-актин (див. Розділ 1). Крім того, він підсилює стимулювання хемотактичної активності нейтрофілів, індукованих комплементом. Спорідненість DBP до провітаміну D3 низька, а до вітаміну D3 висока, отже, DBP переносить вітамін D3 зi шкіри в кров’яне русло. Вітамін D3 також надходить з харчовими продуктами.
Метаболізують вітамін D3 ензими, що є членами надродини цитохрому Р450 (CYP) (див. Розділи 17 і 20). У печінці вітамін D3 перетворюється в 25-гідроксихолекальциферол (кальцидіол, 25-OHD3), а той у клітинах проксимальних канальців нирок - в активніший метаболіт 1,25-дигідроксихолекальциферол, який ще називають кальцитріолом, чи 1,25-(OH)2D3. Також 1,25-дигідроксихолекальциферол утворюється в плаценті, у кератиноцитах шкіри та в макрофагах. У пацієнтів з саркоідозом 1,25-дигідроксихолекальциферол утворюють легеневі альвеолярні макрофаги, очевидно, в разі стимулювання у-інтерфероном. Нормальний рівень у плазмі 25-гідроксихолекальциферолу становить близько 30 нг/мл, а рівень 1,25-дигідроксихолекальциферолу - 0,03 нг/мл (100 пмоль/л). Менш активний метаболіт - 24,25-дигідроксихолекальциферол - теж утворюється в нирках (див. рис. 21-7).
Вітамін D3 та його похідні є секостероїдами, тобто стероїдами з одним відкритим кільцем, у нашому випадку це В-кільце (див. рис. 21-7). Крім того, 1,25-дигідроксихолекальциферол є гормоном, оскільки утворюється в організмі, впливає на клітини-мішені, і його переносить кров.
Механізм дії
Оскільки 1,25-дигідроксихолекальциферол є стероїдом, то не дивно, що він діє через рецептор, який належить до надродини рецепторів, через які стероїди, тиреоїдні гормони та багато інших сполук зумовлюють зміни в експресії генів (див. Розділ 1). Зв’язування стероїду з рецептором впливає на ДНК-зв’язувальну ділянку, і результатом цього є посилення транскрипції одних мРНК та пригнічення - інших.
Дії
Унаслідок дії 1,25-дигідрохолекальциферолу утворюються мРНК, які контролюють утворення білків родини кальбіндинів-D. Вони є представниками надродини тропоніну С - Са2+-зв’язувальних білків, що також містять кальмодулін (див. Розділ 1). Кальбіндин-Ds знайдений у кишці, мозку та нирках людини, а також у багатьох різних тканинах щурів. В епітелії кишки та інших тканинах індукуються два кальбіндини: кальбіндин D9K, який має молекулярну масу 9000 та зв’язує 2 Са2+, і кальбіндин D28K з молекулярною масою 28 000, який зазвичай зв’язує 4 Са2+, хоч і має 6 Са2+-зв’язувальних ділянок. У кишці збільшення рівнів кальбіндину DgK та кальбіндину D28K залежать від транспортування Са2+ однак точний шлях, яким вони сприяють проходженню Са2+ через епітелій кишки, не з’ясований. Є також докази того, що 1,25-дигідроксихолекальциферол збільшує кількість Са2+-Н+-АТФ-азних молекул у клітинах кишок; вони необхідні для нагнітання Са2+ в інтерстиційний простір.
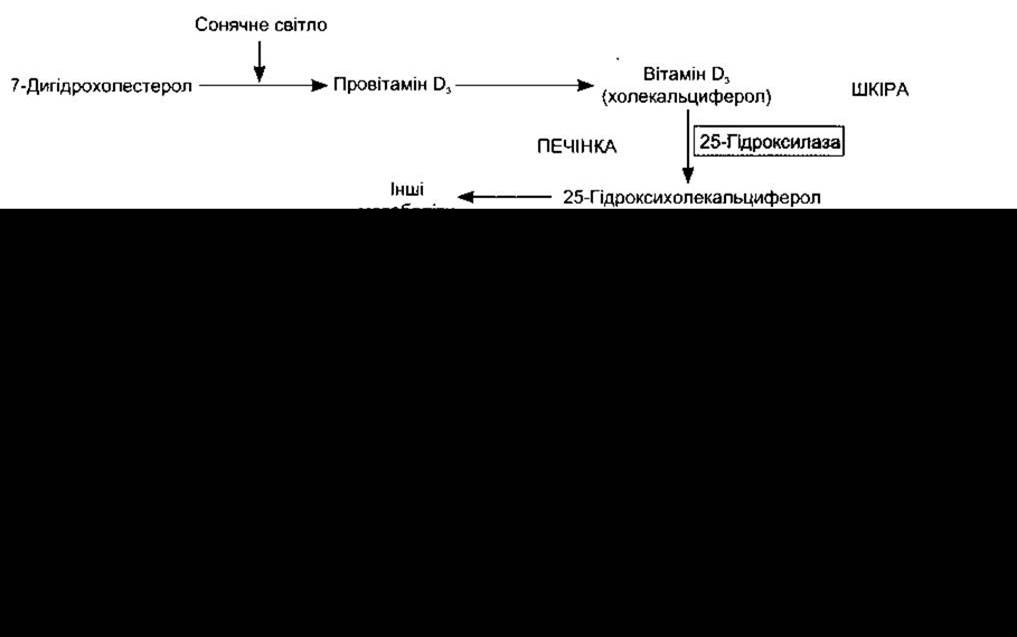
Рис. 21-7. Утворення та гідроксилювання вітаміну D3; 25-гідроксилювання відбувається в печінці, а інші типи - головно в нирках. Наведені також формули 7-дигідроксихолестеролу, вітаміну D3 та 1,25-дигідроксихолекальциферолу.
Окрім збільшення абсорбції Са2+ з кишки, 1,25-дигідроксихолекальциферол сприяє реабсорбції Са2+ в нирках. Він діє на кістки, де мобілізує Са2+ та РО43 , збільшуючи кількість зрілих остеокластів, а також стимулює остеобласти, проте кінцевим результатом знову є мобілізація Са2+.
Рецептори 1,25-дигідроксихолекальциферолу виявлені в багатьох тканинах, окрім кишки, нирок та кісток. До них належать шкіра, лімфоцити, моноцити, скелетні та серцевий м’язи, молочна залоза і передня частка гіпофіза. Щораз більше з’являється доказів того, що 1,25-дигідроксихолекальциферол стимулює диференціацію клітин імунної системи та кератиноцитів у шкірі. Щодо цього простежується збільшення частоти інфекцій у пацієнтів з нестачею вітаміну D. Крім того, з’ясовано, що 1,25-дигідроксихолекальциферол задіяний у регулюванні росту та утворенні факторів росту. Однак його точну роль ще треба дослідити.
Регулювання синтезу
Утворення 25-гідроксихолекальциферолу, здається, не регульоване надто суворо. Проте формування 1,25-дигідроксихолекальциферолу в нирках, яке каталізує 1а-гідроксилаза, регульоване за механізмом зворотного зв’язку вмістом Са2+ та РО43 у плазмі (рис. 21-8). Його утворення прискорює паратгормон, і коли рівень Са2+ в плазмі низький, то секреція паратгормону збільшується. Якщо ж рівень Са2+ у плазмі високий, то утворюється мало 1,25-дигідроксихолекальциферолу, і нирки, натомість, продукують порівняно неактивний метаболіт 24,25-дигідроксихолекальциферол. Цей вплив Са2+ на утворення 1,25-дигідроксихолекальциферолу є механізмом, що забезпечує адаптацію абсорбції Са2+ з кишки (див. вище). Утворення 1,25-дигідроксихолекальциферолу також пришвидшує дія низьких, а сповільнює дія високих рівнів РO43 у плазмі, безпосередньо пригнічуючи вплив РО43 на 1а-гідроксилазу. Додаткове контролювання за утворенням 1,25-дигідроксихолекальциферолу відбувається за допомогою безпосереднього негативного зворотного зв’язку метаболіту на 1а-гідроксилазу, позитивної зворотної дії на формування 24,25-дигідроксихолекальциферолу та безпосередньої дії паращитоподібної залози - інгібування утворення мРНК для паратгормону. Пролактин посилює активність 1а-гідроксилази, і кількість 1,25-дигідроксихолекальциферолу, що циркулює, збільшена під час лактації. Естроген збільшує загальну кількість 1,25-дигідроксихолекальциферолу, що циркулює, однак цей вплив, імовірно, відбувається внаслідок збільшення секреції його зв’язувального білка без помітної зміни вільного 1,25-дигідроксихолекальциферолу. Гіпертиреоїдизм пов’язаний зі зменшенням 1,25-дигідроксихолекальциферолу, що циркулює, та збільшеною частотою остеопорозу. Утворення 1,25-дигідроксихолекальциферолу пригнічуване метаболічним ацидозом. Гормон росту, ХСл та кальцитонін також стимулюють утворення 1.25-дигідроксихолекальциферолу.
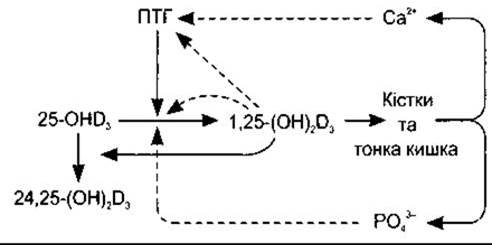
Рис. 21-8. Механізм зворотного контролювання за утворенням 1,25-дигідроксихолекальциферолу (1,25-[OH]2D3) з 25-гідроксихолекальциферолу (25-OHD3) в нирках. Суцільні стрілки позначають стимулювання, штрихові - пригнічення.
Рахіт та остеомаляція
Нестача вітаміну D спричинює порушення кальцифікації кісткового матриксу та хворобу, що названа рахітом у дітей та остеомаляцією в дорослих. У різко вираженому стані в дітей рахіт зумовлює слабість та вигин опорних кісток, зубні дефекти та гіпокальцинемію. В дорослих цей стан менш помітний і найчастіше спричинений недостатньою кількістю сонячного світла в задимлених містах, однак тепер він все щораз є наслідком недостатнього споживання провітамінів, на які діє сонячне світло в шкірі. В таких випадках треба вживати вітамін D. Названі захворювання також можуть бути зумовлені інактивувальними мутаціями в гені ниркової 1а-гідроксилази, у цьому випадку немає відповіді на вітамін D, проте є нормальна відповідь на 1,25-дигідроксихолекальциферол (тип І вітамін-D-резистентного рахіту). В рідкісних випадках хвороба може виникати внаслідок інактивувальних мутацій гена рецептора 1,25-дигідроксихолекальциферолу (тип II вітамін D-резистентного рахіту), за якого простежується недостатня відповідь на вітамін D та 1,25-дигідроксихо- лекальциферол.