Фізіологія людини - Вільям Ф. Ґанонґ 2002
Ендокринна система, метаболізм і репродуктивна функція
Гормональне регулювання метаболізму кальцію та фізіологія кістки
Кальцитонін
Походження
У собак перфузія щитоподібно-паращитоподібної ділянки розчинами, що містять високі концентрації Са2+, веде до зниження рівня периферійного Са2+ у плазмі, після ушкодження ж цієї ділянки інфузії Са2+ спричинюють збільшення Са2+ у плазмі порівняно з контрольними тваринами. Ці та інші спостереження привели до відкриття того, що Са2+-знижувальний та Са2+-підвищувальний гормони секретовані утворами в шиї; Са2+-знижувальний гормон названо кальцитоніном. У хребетних нессавців джерелом кальцитоніну є ультимобранхіальні тільця - пара залоз, що мають ембріологічне походження з п’ятих зябрових дуг. У ссавців ці тільця здебільшого є в щитоподібній залозі, де ультимобранхіальна тканина поширюється навколо фолікулів як парафолікулярні клітини, відомі ще як світлі, або С-клітини (рис. 18-2 та 21-15).
Будова
Кальцитонін людини має молекулярну масу 3500 і містить 32 амінокислотні залишки (рис. 21-16). Велика кількість мРНК, транскрибованої з гена кальцитоніну, зазнає процесинґу в інші мРНК у нервовій системі, завдяки чому утворюється кальцитонін гензалежний пептид (CDRP - з англ. calcitonin gene-related peptide), а не кальцитонін (див. Розділ 4). Кальцитоніни інших досліджених видів також містять 32 амінокислотні залишки, проте амінокислотний склад значно змінюється. Цікавим є кальцитонін лосося, оскільки його активність у 20 разів перевищує людський кальцитонін.
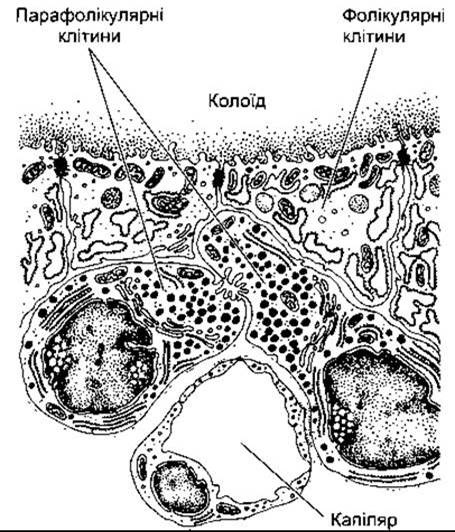
Рис. 21-15. Парафолікулярні клітини в щитоподібній залозі (модифіковано з Poirier J, Dumas JLR: Review of Medical Histology. Saunders, 1977).
Секреція та метаболізм
Секреція кальцитоніну збільшується, коли щитоподібну залозу перфузують розчинами, що містять високі концентрації Са2+. Вимірювання кальцитоніну, що циркулює, радіо- імунологічним аналізом свідчить про те, що він не секретується доти, доки рівень кальцію в плазмі не досягне приблизно 9,5 мг/дл, вище цього рівня у плазмі кальцитонін прямо пропорційний до кальцію. Бета-адренергічні агоністи, дофамін та естрогени також стимулюють секрецію кальцитоніну, як і гастрин, ХЦК, глюкагон та секретин, однак гастрин має найсильніший вплив (див. Розділ 26). Рівень кальцитоніну в плазмі підвищений у разі синдрому Золлінґера-Еллінсона (див. Розділ 26) та перніціозної анемії, за якої також підвищується рівень гастрину в плазмі. Проте доза гастрину, потрібна для стимулювання секреції кальцитоніну, значно збільшує концентрацію гастрину в плазмі, ніж їжа, тому ще рано робити висновок, що кальцій в кишці ініціює секрецію кальційзнижувального гормону до своєї абсорбції.
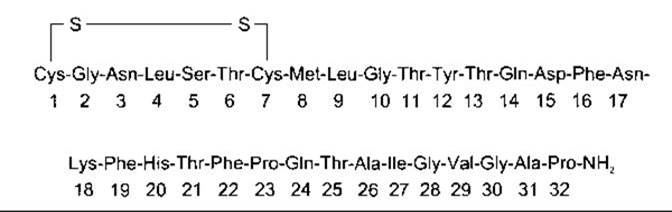
Рис. 21-16. Людський кальцитонін.
Період півжиття людського кальцитоніну - менше 10 хв.
Дії
Серпентинові рецептори для кальцитоніну виявлені в кістках та нирках. Кальцитонін знижує рівні кальцію та фосфату, що циркулюють. Він спричинює свій кальційзнижувальний ефект, пригнічуючи резорбцію кістки. Ця дія пряма, кальцитонін інгібує активність остеокластів in vitro, а також підсилює екскрецію Са2+ з сечею.
Точна фізіологічна роль кальцитоніну невизначена. Вміст кальцитоніну в щитоподібній залозі людини низький, тому після її видалення густина кісток та рівень кальцію в плазмі є нормальними, якщо паращитоподібні залози інтактні. Окрім цього, є лише короткочасні аномалії метаболізму кальцію, якщо його вводять після видалення щитоподібної залози. Це частково можна пояснити секрецією кальцитоніну з інших, крім щитоподібної залози, тканин. Проте панує загальна думка, що гормон має слабкий довготривалий вплив на рівень кальцію в плазмі у дорослих тварин та людини. Крім того, хворі на медулярні карциноми щитоподібної залози мають дуже високі рівні кальцитоніну, що циркулює, однак не мають симптомів, що могли б безпосередньо стосуватися гормону, а їхні кістки є, по суті, нормальними. У разі нестачі кальцитоніну не було описано жодних симптомів. У молодих осіб секретується більше гормону, і він може відігравати роль у формуванні скелета, а також захищати від післяобідньої гіперкальціємії. Кальцитонін захищає кістки матері від надмірної втрати кальцію під час вагітності. Утворення кісток немовляти та лактація є головними споживачами запасів кальцію, і рівні 1,25-дигідроксихолекальциферолу під час вагітності підвищені. Вони б спричинили втрату кісткової маси в матері, якби резорбцію кісток одночасно не пригнічувало збільшення рівня кальцитоніну в плазмі.
Клінічні паралелі
Кальцитонін корисний для лікування хвороби Педжета, стану, за якого збільшена остеокластична активність спричинює компенсаторне утворення неорганізованої нової кістки. Він також має лікувальну дію у разі важкої гіперкальціємії, однак гормон треба вводити у вигляді ін’єкцій і його вплив зазвичай втрачається.
Підсумок
Тепер можна підсумувати дію трьох головних гормонів, що регулюють концентрацію Са2+ в плазмі. Паратгормон збільшує концентрацію кальцію в плазмі, мобілізуючи цей йон з кісток. Він збільшує реабсорбцію Са2+ нирками, однак це може бути наслідком зростання відфільтрованого Са2+, а також посилює утворення 1,25-дигідроксихолекальциферолу, який підсилює абсорбцію Са2+ з кишки, мобілізує йон з кісток та збільшує реабсорбцію кальцію нирками. Кальцитонін пригнічує резорбцію кісток та збільшує кількість Са2+ в сечі.