Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції травної системи
Травлення та всмоктування
Всмоктування води та електролітів
Вода, натрій, калій, хлор
Весь водний баланс у шлунково-кишковому тракті підсумовано у табл. 25-3. Щодня в кишці є близько 2000 мл спожитої рідини і 7000 мл різних секретів, що їх утворюють слизова оболонка шлунково-кишкового тракту і приєднані до нього залози. Із цієї рідини всмоктується 98%, а денні втрати з випорожненнями становлять 200 мл. Лише невелика кількість води проходить через слизову оболонку шлунка, тоді як у тонкій та товстій кишці вода рухається у двох напрямах відповідно до осмотичного Градієнта. Частина Na+ дифундує до або назовні від тонкої кишки згідно з концентраційним градієнтом. Оскільки люмінальні мембрани всіх ентероцитів у тонкій та ободовій кишці проникні для Na+, а базолатеральні мембрани містять Na+-K+-ATФ- азу, то Na+ також активно всмоктується у тонкій та товстій кишці. У тонкій кишці вторинне активне транспортування Na+ важливе для постачання під час всмоктування глюкози, деяких амінокислот (див. вище) та інших речовин, тоді як наявність глюкози в кишковому просторі полегшує реабсорбцію Na+. Цей факт є фізіологічною основою для лікування втрат Na+ і води в разі діареї per os за допомогою різноманітних розчинів, що містять NaCl і глюкозу. Пластівці зернових, що містять вуглеводи, теж використовують для лікування діареї. Такий тип лікування доцільний і у випадку терапії холери - хвороби, що супроводжується вираженою, а якщо не лікувати, то часто фатальною діареєю.
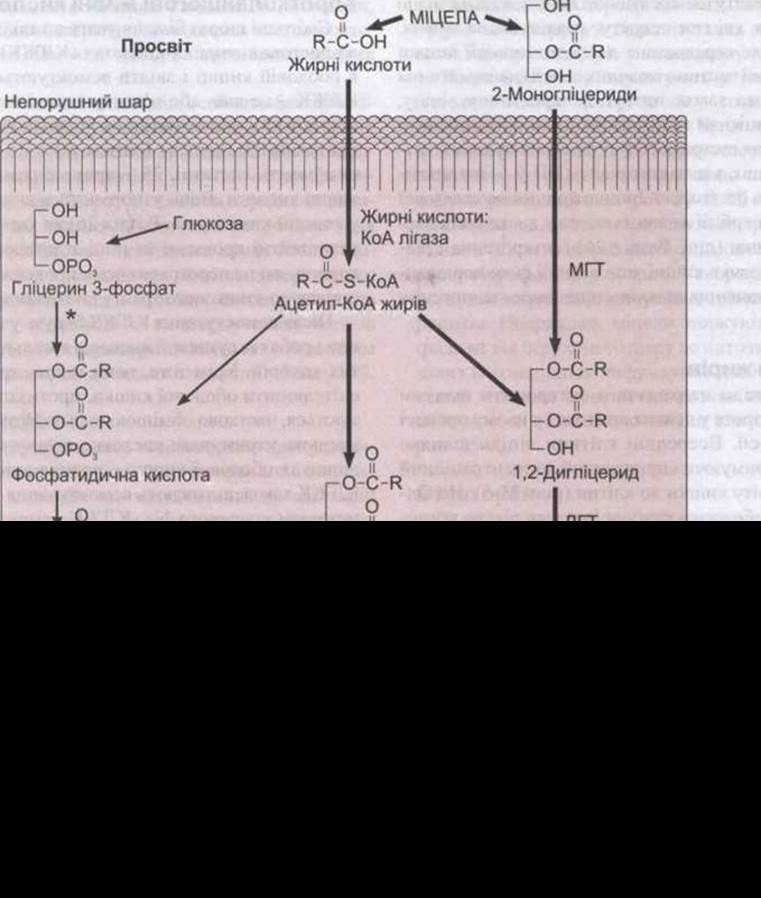
Рис. 25-5. Всмоктування жирів. Тригліцериди утворюються у слизових клітинах з моногліцеридів та жирних кислот. Деякі з гліцеридів також синтезуються з глюкози через фосфатидичну кислоту. Згодом тригліцериди перетворюються у хіломікрони, і їх вивільняє екзоцитоз. З позаклітинного простору вони потрапляють у лімфу. Товсті лінії позначають головні шляхи; * - реакцію інгібують моногліцериди; МГТ - моноацилгліцеридацилтрансфераза; ДГТ - діацилгліцеридацилтрансфераза.
За нормальних умов Сl з інтерстиційної рідини входить в ентероцити через Na+-K+ -2Cl- котранспортери базолатеральних мембран (рис. 25-7). Згодом він секретується в просвіт кишки через канали, регульовані численними білковими кіназами. Один з них активує протеїнкіназа А і згодом цАМФ. У випадку холери концентрація цАМФ збільшується. Холерний вібріон перебуває в просвіті кишки, однак він виробляє токсин, що зв’язується з GM-1 гангліозидним рецептором, а це дає змогу частині А-субодиниці (А1-пептиду) токсину увійти в клітину. Пептид А1 зв’язує аденозиндифосфат рибози до а-субодиниці Gs, що гальмує ГТФ-азну активність (див. Розділ 1). Отже, конститутивно активований G-білок зумовлює тривале стимулювання аденілатциклази і помітне підвищення внутрішньоклітинної концентрації цАМФ. Поряд зі зростанням секреції Сl- функція носія слизової оболонки для Na+ зменшується, і, відповідно, зменшується абсорбція NaCl. Результатом стає збільшення водного об’єму та електролітного вмісту у кишці і, як наслідок, поява діареї. Крім того, Na+-K+-АТФ-аза і Na+/глюкоза котранспортер не активовані, тому й подвійна опосередкована реабсорбція глюкози і Na+ теж неповноцінна.
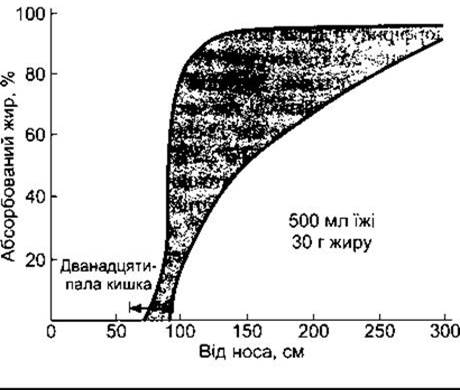
Рис. 25-6. Динаміка всмоктування жирів у людей, що ґрунтується на вимірюваннях після приймання жирної їжі. Стрілка позначає дванадцятипалу кишку (відтворено за дозволом з Davenport HW: Physiology of the Digestive Tract, 2nd ed. Year Book, 1966).
Вода рухається в кишку та у зворотному напрямі тільки тоді, коли осмотичний тиск інтестинального вмісту та плазми однакові. Залежно від спожитих харчів осмоляльність дуоденального вмісту може бути гіпертонічною або гіпотонічною, однак на момент надходження в голодну кишку стає близькою до осмоляльності плазми. Така ж осмоляльність утримується в міжтравний період у кишці. Осмотично активні частинки, що формуються під час травлення, всмоктуються, і вода рухається пасивно назовні від кишки за новоутвореним осмотичним градієнтом. В ободовій кишці відбувається активне випомповування Na+ назовні, вода рухається пасивно разом з ним знову за осмотичним градієнтом. Мінеральні послаблювальні засоби, такі як магнезії сульфат, що належить до осмотично активних солей, які погано абсорбуються, утримують воду в кишці для відновлення осмотичної рівноваги. Унаслідок збільшення інтестинального об’єму відбувається послаблювальний ефект.
Таблиця 25-3. Щоденний баланс води у шлунково-кишковому тракті1, мл
Спожито |
2000 |
|
Ендогенні секрети |
7000 |
|
Слинні залози |
1500 |
|
Шлунок |
2500 |
|
Жовч |
500 |
|
Підшлункова залоза |
1500 |
|
Кишка |
1000 |
|
7000 |
||
Загальна кількість |
9000 |
|
Реабсорбовано |
8800 |
|
Голодна кишка |
5500 |
|
Клубова кишка |
2000 |
|
Ободова кишка |
1300 |
|
8800 |
||
Вміст у калових масах |
200 |
1 Дані з Moore EW: Physiology of Intestinal Water and Electrolyte Absorption, American Gastroenterological Society, 1976.
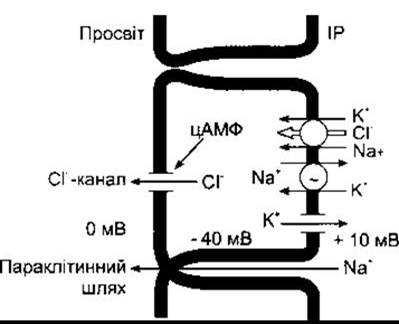
Рис. 25-7. Рух йонів через ентероцити у тонкій кишці: Cl- потрапляє у клітину через Nа+-К+-2Сl- -котранспортер базолатеральної поверхні і секретує у кишковий простір через Cl- - канали, деякі з яких активовані цАМФ; К+ рециклює в інтестинальну рідину (IP) через базолатеральні К+-канали (відтворено за дозволом з Field М, Roa MC, Chang ЕВ: Intestinal electrolyte transport and diarrheal disease. N Engl J Med 1989;321: 800).
У просвіт кишки секретує незначна кількість К+ спеціально як компонент слизу, а більша частина надходить завдяки руху К+ через шлунково-кишкову слизову оболонку шляхом дифузії. З іншого боку, К+ теж секретує в ободову кишку, оскільки К+-канали є як у люмінальній, так і у базолатеральній мембранах ентероцитів ободової кишки. Рух К+ відбувається пасивно за електрохімічним градієнтом.
Акумуляція К+ в ободовій кишці частково компенсована Н+-К+-АТФ-азою у люмінальній мембрані клітин дистального відділу ободової кишки з кінцевим активним транспортуванням К+ до клітин. Незважаючи на наявність такого механізму, втрати рідини в разі хронічних діарей можуть призводити до вираженої гіпокаліємії.
У випадках тривалого надмірного харчового приймання К+ збільшується секреція альдостерону, і більше К+ надходить до ободової кишки. Це зумовлене частково появою більшості Nа+-К+-АТФ-азних помп у базолатеральних мембранах клітин і, як наслідок, зростанням внутрішньоклітинного вмісту К+ і дифузії К+ через люмінальні мембрани клітин.