Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції травної системи
Механізми регулювання шлунково-кишкових функцій
Загальні положення
Організація
Принцип організації структур, з яких утворена стінка шлунково-кишкового тракту від задньої глотки до відхідника, показано на рис. 26-1. Є незначні відмінності, однак назагал вона містить чотири шари (від кишкового просвіту: слизовий, підслизовий, м’язовий та серозний). У підслизовому шарі розташовані волокна гладких м’язів, а у м’язовому зовнішньому - два шари гладких м’язів: поздовжній та внутрішній косий. Уздовж травний тракт вистелений слизовою оболонкою, за винятком стравоходу та дистальної частини прямої кишки, які вкриті серозною оболонкою. Ця оболонка, відповідно, продовжена мезентерієм, який містить нервові волокна, лімфатичні та кровоносні судини, що оточують травний тракт.
Кровообіг у шлунково-кишковому тракті
Плин крові до шлунка, кишок, підшлункової залози та печінки забезпечує серія паралельних кіл, з яких кров від кишки та підшлункової залози потрапляє через портальну вену до печінки. Фізіологія цієї важливої ділянки системи кровообігу описана у Розділі 32.
Ентерична нервова система
Усередині шлунково-кишкового тракту є дві мережі нервових волокон: міентеричне сплетення (Ауербахівське сплетення) між зовнішнім поздовжнім і середнім циркуляторним м’язовими шарами, та підслизове сплетення (Мейснерівське сплетення) між середнім циркуляторним шаром і слизовою оболонкою (див. рис. 26-1). Разом ці нейрони утворюють ентеричну нервову систему. Така система у людей містить понад 100 мільйонів сенсорних нейронів, інтер- та мотонейронів, тобто більше, ніж є в цілому спинному мозку, тому, мабуть, найліпше її розглядати як «переміщену» частину ЦНС, що відповідає за регулювання функцій травного тракту. Ця система пов’язана з ЦНС за допомогою парасимпатичних та симпатичних волокон, однак може діяти автономно без цих зв’язків (див. нижче). Міентеричне сплетення іннервує поздовжній і циркуляторний гладкі м’язові шари та відповідає головно за регулювання кишкової секреції. До нейротрансмітерів ентеричної нервової системи належать ацетилхолін, аміни - норадреналін та серотонін, амінокислоти - ГАМК, пурин АТФ, гази NO й СО та багато інших пептидів і поліпептидів (табл. 26-1). Деякі з цих пептидів також впливають у паракринний спосіб, а інші потрапляють у кровоплин і діють як гормони. Відомо також, що більшість з них міститься і в головному мозку.
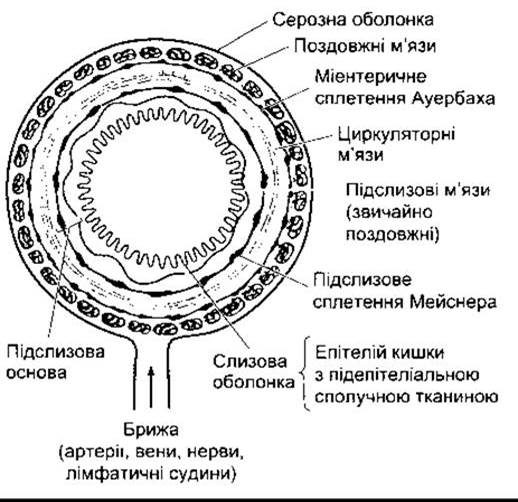
Рис. 26-1. Схематичне зображення шарів стінки шлунка, тонкої та ободової кишки. Будова стравоходу і дистальної частини прямої кишки подібна, за винятком того, що в них нема серозної оболонки або брижі. Крім того, м’язи верхньої чверті стравоходу поперечно-посмуговані, у перехідній частині - змішані: як гладкі, так і поперечно-посмуговані, а в дистальній - лише гладкі (відтворено за дозволом з Bell GH, Emslie-Smith D, Paterson CR: Textbook of Physiology and Biochemistry, 9th ed. Churchill Livingstone, 1976).
Зовнішня іннервація
Кишка забезпечена подвійною зовнішньою іннервацією: як парасимпатичною, холінергічною (зі збільшенням активності гладких м’язів кишки), так і симпатичною, норадренергічною (з протилежними ефектами і скороченнями сфінктерів травної системи). Прегангліонарні парасимпатичні волокна складаються з понад 2000 еферентних волокон блукаючого нерва та інших волокон, що є у складі крижових нервів. Назагал вони закінчуються в холінергічних нейронах міентеричних та підслизових сплетеннях. Симпатичні волокна - це постгангліонарні, однак багато з них закінчується в постганліонарних холінергічних нейронах, де власний секретований норадреналін гальмує ацетилхолінову секрецію внаслідок активування а2-пресинаптичних рецепторів. Деякі симпатичні волокна закінчуються безпосередньо на гладких м’язових клітинах кишки. Біоелектричні властивості інтестинальних гладких м’язів описані у Розділі 3. Інші волокна іннервують кровоносні судини кишки і спричинюють їхню вазоконстрикцію. Вони характерні для тих кровоносних судин кишки, де є подвійна іннервація (зовнішня адренергічна та внутрішня іннервація ентеричною нервовою системою). ВІП та NO належать до нейротрансмітерів внутрішньої іннервації та відповідають за появу гіперемії, що супроводжує перетравлення їжі. Досі остаточно не з’ясована локалізація додаткової холінергічної іннервації кровоносних судин.
Таблиця 26-1. Головні пептиди ентеричної нервової системи

Перистальтика
Перистальтика - це рефлекторна відповідь, що розпочинається після розтягнення стінки кишки її власним вмістом і простежується уздовж усього шлунково-кишкового тракту - від стравоходу до прямої кишки. Розтягнення розпочинає циркуляторне скорочення м’язів вище від місця подразнення і в ділянці розслаблення перед ним. Хвиля скорочення, яка йде від ротової порожнини у каудальному напрямі, проштовхує вміст просвіту вперед зі швидкістю від 2 до 25 см/с. Перистальтичну активність кишки збільшують або зменшують автоматично вхідні імпульси, однак вона також залежить від зовнішньої іннервації. Крім того, прогресивний рух вмісту не блокований видаленням і повторним ушиванням сегмента кишки в її попередньому положенні, а гальмований лише тоді, коли сегмент у зворотному напрямі повернеться у попереднє положення. Перистальтика є прикладом інтегративної активності ентеричної нервової системи. Вона виявляється у вивільненні серотоніну в разі місцевого розтягнення, що активує сенсорні нейрони, які, відповідно, збуджують міентеричне сплетення. Холінергічні нейрони проходять у цих сплетеннях у ретроградному напрямі й активують нейрони, що вивільняють речовину Р та ацетилхолін, спричинюючи скорочення гладких м’язів. У цей час холінергічні нейрони, що проходять в антеградному напрямі, активують нейрони, що секретують NО, ВІП і АТФ, зумовлюючи розслаблення розташованих вище ділянок від дії подразника.
Базальна електрична активність та регулювання моторики
Гладкі м’язи шлунково-кишкової системи, крім стравоходу та проксимальної ділянки шлунка, мають спонтанну біоелектричну активність. Коливання мембранного потенціалу сягають від -65 до -46 мВ. Цей вихідний базальний електричний ритм (БЕР) спричинюють інтестинальні клітини Кажала, зіркоподібні пейсмейкерні клітини мезенхіми з властивостями гладких м’язів, які простягають довгі множинно-розгалужені відростки до гладких м’язів кишки. У шлунку та тонкій кишці ці клітини містяться у зовнішньому циркуляторному м’язовому шарі біля міентеричного сплетення, у товстій кишці - біля підслизового краю циркулярного м’язового шару. У шлунку та тонкій кишці простежується низхідний градієнт пейсмейкерної частоти, тоді як у серці пейсмейкер домінує, звичайно, з більшою частотою. Сам по собі БЕР зрідка спричинює м’язові скорочення, розряди пікових потенціалів (або спайків, з англ. spike) нашаровуються над більшістю деполяризаційних частин хвиль БЕР, що збільшують м’язове напруження (рис. 26-2). Деполяризаційна частина кожного пікового потенціалу відповідає надходженню Са2+, а реполяризаційна - виділенню К+. На БЕР впливають численні поліпептиди та нейротрансмітери. Наприклад, ацетилхолін збільшує кількість пікових потенціалів та напруження гладких м’язів, тоді як адреналін - зменшує як кількість спайків, так і напруження гладких м’язів (див. рис. 26-2). У шлунку рівень БЕР становить 4/хв, у дванадцятипалій кишці - 12/хв, у дистальній частині клубової кишки знижується до 8/хв. У товстій кишці БЕР збільшується від 9/хв для сліпої кишки до 16/хв для сигмоподібної. БЕР координує перистальтику та інші види моторної активності, і скорочення відбуваються лише під час деполяризаційної частини хвиль. Наприклад, після ваготомії або перетинання стінки шлунка перистальтика в шлунку стає іррегулярною та хаотичною.

Рис. 26-2. Базальний електричний ритм (БЕР) гладких м’язів шлунково-кишкового тракту. Угорі: морфологія та зв’язок із м’язовим скороченням. Унизу: стимулювальний вплив ацетилхоліну та інгібувальний вплив адреналіну (модифіковано та відтворено за дозволом з Chang ЕВ, Sitrin MD, Black DD: Gastrointestinal Hepatobiliary and Nutritional Physiology. Lippincott-Raven, 1996).
Міґрувальний моторний комплекс
У міжтравний період відбувається модифікація електричної та моторної активності гладких м’язів шлунково-кишкової системи. Це призводить до того, що цикли моторної активності мігрують від шлунка до дистальної частини клубової частини. Кожний цикл, або міґрувальний моторний комплекс (ММК), починається періодом спокою (фаза І), що переходить у період іррегулярної та механічної активності (фаза II) і закінчується «вибухом» регулярної активності (фаза III) (рис. 26-3). Міграція ММК в аборальному напрямі відбувається зі швидкістю близько 5 см/хв і настає через кожні 90 хв. Їхня функція остаточно не з’ясована, однак відомо, що шлункова секреція, жовчевиділення та панкреатична секреція збільшуються під час кожного ММС. Можливо, вони сприяють евакуації вмісту зі шлунка і тонкої кишки, готуючи їх до наступного приймання їжі. Під час їди ці комплекси негайно припиняють активність і повертаються до перистальтики й інших форм БЕР та пікових потенціалів.
Інші аспекти м’язових скорочень кишки є високоспецифічними в кожному відділі й описані у частинах, присвячених цим відділам.