Фізіологія людини - Вільям Ф. Ґанонґ 2002
Кровообіг
Регулювання серцево-судинної системи
Механізм системного нервового регулювання
Нервові механізми регулювання
Хоча артеріоли та інші судини опору мають найщільнішу іннервацію, усі кровоносні судини, за винятком капілярів і венул, містять гладкі м’язи й іннервовані руховими волокнами із симпатичного відділу автономної нервової системи. Волокна, що проходять до судин опору, регулюють тканинний кровообіг і рівень артеріального тиску; волокна, що проходять до ємнісних судин, регулюють об’єм крові, яка депонується у венах. Більшість вен має незначну іннервацію, однак вени внутрішніх органів - багату. Звуження вен відбувається під дією тих самих подразників, які активують судинозвужувальні нерви артеріол. Унаслідок цього зменшується ємність венозних судин, збільшується кількість крові, яка надходить до серця, і кров перерозподіляється в артеріальний відділ системи кровообігу.
Іннервація кровоносних судин
Норадренергічні волокна закінчуються в судинах усіх ділянок організму (рис. 31-5). За функцією вони судинозвужувальні. Крім вазоконстрикторної іннервації, судини опору скелетних м’язів мають судинорозширювальні волокна, які хоча й проходять з симпатичними нервами, належать до холінергічних (симпатична судинорозширювальна система). Є дані, які свідчать про наявність холінергічної іннервації у кровоносних судинах серця, легень, нирок і матки. Пучки норадренергічних і холінергічних волокон утворюють сплетення у зовнішньому шарі стінки артеріол. Волокна з численними варикозними розширеннями поширюються від цих сплетень до середньої оболонки і закінчуються на зовнішній поверхні гладких м’язів, не проникаючи в них. Трансмітери проникають у внутрішні ділянки середньої оболонки шляхом дифузії, і біопотенціал поширюється від однієї гладком’язової клітини до іншої через щілинні контакти.

Рис. 31-5. Норадренергічні нервові волокна, що іннервують кровоносні судини очеревини щурів. Стрілками позначено напрям плину крові (відтворено за дозволом з Furness JB, Marshall JM: Correlation of the directly observed responses of mesentric vessels of the rat to nerve stimulation and noradrenaline with the distribution of adrenetic nerves. J Physiol 1974;75: 239).
Тонічні імпульси не характерні для судинорозширювальних волокон, однак вазоконстрикторним волокнам більшості судинних лож притаманна деяка тонічна активність. У випадку перерізання симпатичних нервів (симпатектомії) кровоносні судини розширюються. У більшості тканин розширення судин зумовлене зменшенням рівня тонічної активності вазоконстрикторних нервів. Однак у скелетних м’язах судини можуть розширюватися також унаслідок активування симпатичної судинорозширювальної системи (табл. 31-3).
Багато кровоносних судин іннервовані волокнами, в яких утворюються поліпептиди. Холінергічні нерви містять також ВІП, який спричинює розширення судин. У норадренергічних постганглюнарних симпатичних волокнах формуються також нейропептид Y, який є вазоконстриктором. У чутливих нервах, які закінчуються біля кровоносних судин, містяться речовина Р і CGRPoc, які зумовлюють розширення судин.
Таблиця 31-3. Фактори, які впливають на діаметр артеріол
Звуження |
Розширення |
Місцеві фактори Місцеве зниження температури Авторегулювання |
Збільшення вмісту СО2 і зменшення О2 Збільшення вмісту К+, аденозину, лактату та ін. Місцеве зниження pH Місцеве підвищення температури |
Ендотеліальні фактори Ендотелін-1 Місцево виділений серотонін тромбоцитів Тромбоксан А2 |
NО Кініни Простациклін |
Гормони, що циркулюють у крові Адреналін (за винятком скелетних м’язів і печінки) Норадреналін Аргініновий вазопресин (АВП) Ангіотензин II Циркулювальний інгібітор Nа+-К+-АТФ-ази Нейропептид Y |
Адреналін у скелетних м’язах і печінці CGRPa Речовина Р Гістамін ANP ВІП |
Нервові фактори Посилена імпульсація норадренергічних вазомоторних нервів |
Послаблена імпульсація норадренергічних вазомоторних нервів Активування холінергічних судинорозширювальних волокон, що проходять до скелетних м’язів |
Аференіні імпульси в чутливих нервах від шкіри переходять антидромно вниз по гілках чутливих нервів, які іннервують кровоносні судини. Ці імпульси призводять до виділення речовини Р з нервових закінчень. Речовина Р спричинює розширення судин і підвищення проникності капілярів. Цей місцевий нервовий механізм називають аксонним рефлексом (див. рис. 32-17). Інші серцево-судинні рефлекси відбуваються за участю центральної нервової системи.
Іннервація серця
Імпульси, що надходять по норадренергічних нервах до серця, зумовлюють збільшення частоти (хронотропний ефект) і сили серцевих скорочень (інотропний ефект). Вони пригнічують також ефекти стимулювання блукаючих нервів, можливо, завдяки виділенню нейропептиду Y, який є ко трансмітером у симпатичних нервових закінченнях. Імпульси, які надходять до серця по холінергічних волокнах блукаючих нервів, зменшують частоту серцевих скорочень. У стані спокою для симпатичних нервів серця характерна помірна тонічна активність. У людей та великих тварин їй протидіє більше виражена тонічна імпульсація блукаючих нервів (тонус блукаючих нервів). У тварин в експерименті з перерізаними блукаючими нервами частота серцевих скорочень збільшується. Після введення парасимпатолітичних препаратів, таких як атропін, частота серцевих скорочень у людей збільшується з 70 (у стані спокою в нормі) до 150-180 ударів за 1 хв. Це пояснюють тим, що парасимпатичний тонус перестає протидіяти симпатичному. У людей в ран одночасної блокади норадренергічної і холінергічної систем частота серцевих скорочень становить приблизно 100 ударів за 1 хв.
Вазомоторний контроль
Симпатичні нерви, які спричинюють звуження артеріол і вен, а також збільшують ударний об’єм, мають тонічну активність (постійно посилюють нервові імпульси). Регулювання рівня артеріальною тиску відбувається внаслідок зміни цієї тонічної активності (рис. 31-6). Артеріальний тиск залежить від активності рефлексів спинного мозку, однак передусім його контролює група нейронів довгастого мозку, яку називають вазомоторною ділянкою, або вазомоторним центром.
Нейрони, які забезпечують посилення симпатичного впливу на кровоносні судини і серце (рис. З 1 -7), проходять до симпатичних прегангліонарних нейронів у складі проміжнобічного (11 Б) стовпа спинного мозку. З обох боків тіла цих нейронів містяться ближче до м’якої оболонки в росіральній вентролагеральній ділянці довгастого мозку (РВЛД). Їхні аксони проходять дорсально і медіально, а пOТІм опускаються в складі бічного стовпа спинного мозку до ПБ стовпа.
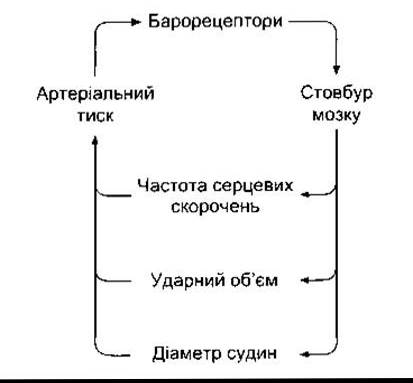
Рис. 31-6. Контроль рівня артеріального тиску за принципом зворотного зв’язку
Імпульси, які надходять із довгастого мозку, діють також на частоту серцевих скорочень через блукаючі нерви. Нейрони, від яких починаються волокна блукаючих нервів, містяться у дорсальному ядрі блукаючого нерва і подвійному ядрі (рис. 31-8).
З посиленням вазоконстрикторного впливу простежується звуження артеріол і підвищення артеріального тиску. Здебільшого ці зміни супроводжуються звуженням вен і зменшенням об’єму депонованої венозної крові, хоча не завжди у ємнісних судинах відбуваються зміни, аналогічні до змін у судинах опору. Частота серцевих скорочень і ударний об’єм крові теж збільшуються з посиленням активності симпатичного відділу. Одночасно збільшується хвилинний об’єм серця. У цьому разі, як звичайно, послаблюється тонічна активність волокон блукаючих нервів, що іннервують серце. І навпаки, у випадку послаблення активності вазомоторного центру судини розширюються, артеріальний тиск знижується, депонування крові у ємнісних судинах посилюється. Здебільшого одночасно зменшується частота серцевих скорочень. Однак цей ефект головно зумовлений посиленням впливу блукаючих нервів на серце.
Аферентні шляхи до вазомоторного центру
Аферентні волокна, які сходяться у вазомоторному центрі, наведені у табл. 31-4. До них належать не тільки дуже важливі волокна, що проходять від барорецепторів артерій і вен, а й волокна від інших ділянок нервової системи, а також від каротидних та аортальних хеморецепторів. Крім того, деякі стимули діють безпосередньо на вазомоторний центр.
Є низхідні шляхи до вазомоторного центру, які проходять від кори головного мозку (зокрема, лімбічної кори) і прямують до гіпоталамуса. Ці волокна беруть участь у підвищенні артеріального тиску і розвитку тахікардії під впливом емоційних факторів, таких як статеве збудження та гнів. Зв’язки між гіпоталамусом і вазомоторним центром є рецинрокними, а аферентні імпульси зі стовбура мозку замикають петлю.
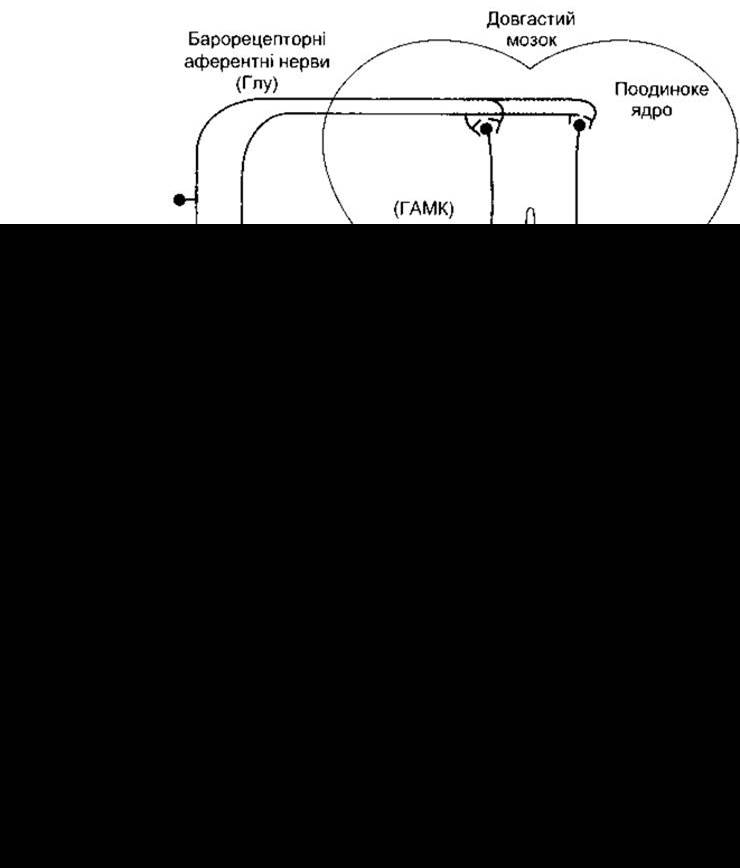
Рис. 31-7. Головні шляхи, які беруть участь у регулюванні довгастим мозком рівня артеріального тиску Показані еферентні шляхи блукаючого нерва, які сповільнюють роботу серця У круглих дужках зазначені можливі нейротрансмітери нервових шляхів; Глу - глутамат; ГАМК - у-аміномасляна кислота; Ах - ацетилхолін; НА - норадреналін; ПБ - проміжнобічний стовп; ПоЯ - поодиноке ядро; КВЛД, ПВЛД, РВЛД - каудальна, проміжна і ростральна вентролатеральні ділянки довгастого мозку; IX і X - язикогорловий і блукаючий нерви.
Наповнення легень повітрям приводить до розширення судин і зниження артеріального тиску. Ця реакція відбувається за участю аферентних волокон блукаючого нерва, що проходять від легень і спричинюють послаблення активності вазомоторного центру. Біль, як звичайно, зумовлює підвищення артеріального тиску завдяки аферентним імпульсам, що переходять від ретикулярної формації до вазомоторного центру. Однак тривалий біль може спричиняти розширення судин і запаморочення.
Соматосимпатичний рефлекс
Біль зумовлює підвищення артеріального тиску. Аферентні імпульси від скелетних м’язів, що працюють, можливо, спричинюють подібний пресорний ефект. Ця реакція відбувається за участю СІ нейронів ростральної вентролатеральної ділянки довгастого мозку. Звуження судин у відповідь на стимулювання соматичних аферентних нервів називають соматосимпатичним рефлексом. Нервові шляхи, які беруть участь у цій реакції, зображено на рис. 31 -9.
Барорецептори
Барорецептори - це рецептори, подразнення яких відбувається під час розтягування стінок серця і кровоносних судин. Рецептори сонної пазухи (каротидного синуса) і дуги аорти беруть участь у регулюванні артеріального кровообігу. Барорецептори містяться також у стінках правого і лівого передсердь у місцях впадіння верхньої та нижньої порожнистих і легеневих вен, а також у судинах легень, їх у відділах малого кола кровообігу з низьким тиском називають кардіопульмональними рецепторами. Оскільки подразнення барорецепторів відбувається в разі розтягування ділянок, у яких вони локалізовані, то підвищення тиску у цих структурах призводить до сильнішої імпульсації від барорецепторів. Імпульси від барорецепторів надходять по аферентних волокнах язикогорлового і блукаючого нервів до довгастого мозку. Більшість цих волокон закінчується у ядрі поодинокого шляху (ЯПШ), збуджувальним трансмітером названих волокон є, можливо, глутамат. Від ЯПШ відходять збуджувальні глутамінергічні волокна до каудальної і проміжної вентролатеральних ділянок довгастого мозку, де вони стимулюють гальмівні нейрони, які виділяють ГАМК. Ці нейрони надходять у ростральну вентролатеральну ділянку. Є також збуджувальні, можливо багатонейронні, шляхи від ЯПШ до моторних нейронів блукаючого нерва у дорсальному ядрі і до подвійного ядра. Отже, посилення імпульсації від барорецепторів пригнічує тонічну активність судинозвужувальних симпатичних нервів і стимулює дію блукаючих нервів на серце, розширення артеріол і вен, що призводить до зменшення хвилинного об’єму і зниження артеріального тиску.
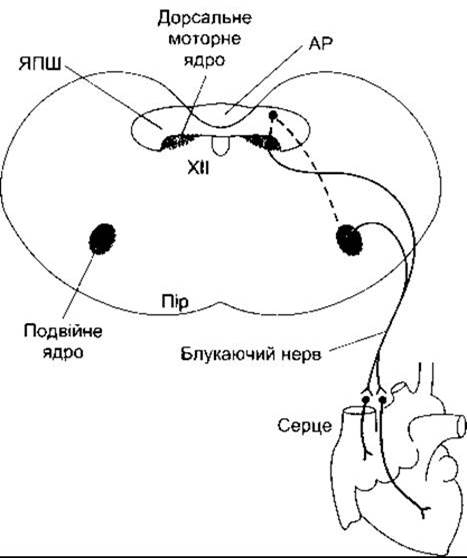
Рис. 31-8. Головні шляхи, які беруть участь у регулюванні довгастим мозком частоти серцевих скорочень за допомогою блукаючих нервів. Нейрони ЯПШ (штрихова лінія) проходять до прегангліонарних парасимпатичних нейронів дорсального моторного ядра блукаючого нерва і подвійного ядра. Постгангліонарні холінергічні нейрони іннервують передсердя і шлуночки; Пір - піраміда; XII - ядро під’язикового нерва (модифіковано з Standish R, Enquist LH, Schwaber JS: Innervation of the heart and its central medullary origin defined by viral tracing. Science 1994;263:232).
Таблиця 31-4. Фактори, що впливають на активність вазомоторного відділу довгастого мозку


Рис. 31-9. Соматосимпатичний рефлекс. Імпульси від рецепторів шкіри та м’язів проходять до РВЛД довгастого мозку, аксони клітин РВЛД - безпосередньо до проміжнобічного стовпа (ПБ), де вони закінчуються на прегангліонарних симпатичних нейронах; НМН - нижня мозочкова ніжка; НОЯ - нижнє оливне ядро; ППП - присередній поздовжній пучок; ПЯ - подвійне ядро; ПоЯ - поодиноке ядро; П - піраміда; ЯСШТН - ядро спинномозкового шляху трійчастого нерва (відтворено за дозволом з Reis DJ, Ruggerio DA, Morrison SF: The C, area of the rostal ventrolateral medulla oblongata-A critical brainstem region for control of resting and reflex integration of arterial pressure. Am J Hypertens 1989;2:363S).
Сонна пазуха і дуга аорти
Сонна пазуха - це незначне розширення внутрішньої сонної артерії відразу над місцем біфуркації загальної сонної артерії на зовнішню і внутрішню гілки (рис. 31- 10). У ділянці цього розширення містяться барорецептори, що локалізовані також у стінці дуги аорти. Рецептори розміщені у зовнішньому шарі судинної стінки. Вони є дуже розгалуженими, шишкоподібними, закрученими закінченнями мієлінових нервових волокон, які нагадують сухожильний орган Гольджі. У деяких видів подібні рецептори містяться у різних ділянках великих артерій грудної клітки і шиї. Аферентні волокна, які проходять від сонної пазухи і сонного клубочка, утворюють окрему гілку язикогорлового нерва - гілку сонної пазухи. Однак волокна від дуги аорти формують окрему гілку блукаючого нерва лише у кролів. Нерви сонної пазухи і гілки блукаючого нерва, які проходять від дуги аорти, часто називають буферними нервами.
Активність буферних нервів
За нормального артеріального тиску волокна буферних нервів мають низький рівень імпульсації (рис. 31-11). Підвищення тиску в сонній пазусі та дузі аорти призводить до посилення імпульсації, а зниження - до її послаблення.
Якщо одну сонну пазуху мавпи ізолювати і підтримувати її перфузію та одночасно виконати денервацію інших барорецепторів, то в разі перфузійного тиску до 30 мм рт. ст. імпульси в аферентних волокнах від сонної пазухи не виникають, і артеріальний тиск та частота серцевих скорочень не знижуються. При перфузійному тиску 70-110 мм рт. ст. є лінійна залежність між перфузійним тиском і зниженням артеріального тиску й частоти серцевих скорочень. Якщо перфузійний тиск понад 150 мм рт. ст., то подальшого суттєвого посилення реакції не відбувається (рис. 31-12), очевидно, тому, що імпульсація від барорецепторів і пригнічення вазомоторного центру досягають максимального рівня.
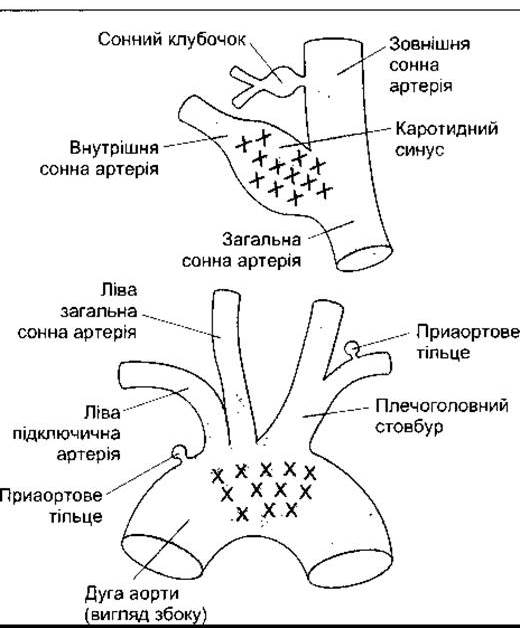
Рис. 31-10. Барорецепторні ділянки каротидного синуса і дуги аорти. Локалізацію рецепторів позначено через X.

Рис. 31-11. Імпульси (вертикальні лінії) в одному аферентному нерві, які надходять від каротидного синуса, за різних значень артеріального тиску; співвіднесені зі змінами аортального тиску в часі (відтворено за дозволом з Berne RM, Levy MN: Cardiovascular Physiology, 3rd ed Mosby, 1977).
Рецептори сонної пазухи реагують як на підтримувальний, так і на пульсовий тиск. Зниження пульсового тиску в сонних артеріях без будь-яких змін середнього тиску веде до меншої імпульсації від барорецепторів і зумовлює підвищення артеріального тиску й тахікардію. Рецептори реагують також як на зміни тиску, так і на стабільний тиск. У випадках коливання тиску деколи за його підвищення імпульси виникають, а в разі зниження - не виникають (див. рис. 31-11). У цьому випадку середній тиск без коливань спричиняв би постійну імпульсацію.
Рецептори дуги аорти вивчали не настільки детально, однак немає підстав уважати, що їхня відповідь суттєво відрізняється від відповіді рецепторів сонної пазухи.
Зі сказаного стає очевидним, що барорецептори артеріального відділу кровообігу, їхні аферентні зв’язки з вазомоторним і кардіоінгібіторним відділами, а також еферентні шляхи від цих відділів утворюють рефлекторний механізм зворотного зв’язку, спрямований на стабілізацію артеріального тиску і частоти серцевих скорочень. Будь-яке зниження системного артеріального тиску послаблює пригнічувальну імпульсацію в буферних нервах і спричинює компенсаторне підвищення артеріального тиску та хвилинного об’єму. Будь-яке підвищення тиску зумовлює розширення артеріол і зменшення хвилинного об’єму до того моменту, доки артеріальний тиск не досягне нормального рівня.
Переустановлення барорецепторів
У випадку хронічної гіпертензії барорецепторний рефлекс переустановлюється на підтримку не нормального, а підвищеного тиску. Під час перфузійних досліджень на експериментальних тваринах з гіпертензією з’ясовано, що підвищення тиску в ізольованій сонній пазусі призводить до зниження підвищеного системного артеріального тиску. Зниження перфузійного тиску веде до зростання підвищеного тиску (рис. 31-12). Переустановлення швидко розвивається в експериментальних тварин, однак мало відомо про те, як і чому це відбувається. Як в експериментальних, так і в клінічних умовах це зворотний процес.

Рис. 31-12. Суцільна лінія: зниження системного артеріального тиску з підвищенням тиску в ізольованому каротидному синусі нормальної мавпи. Штрихова лінія: реакція гіпертензивної мавпи, яка свідчить про переустановлення барорецепторів (стрілка).
Наслідки перетискання сонних артерій і перерізання буферних нервів
Двобічне перетискання сонних артерій ближче до сонних пазух зумовлює підвищення артеріального тиску і частоти серцевих скорочень, оскільки в цьому разі знижується тиск у сонних пазухах. Перерізання з кожного боку нервів сонних пазух веде до аналогічного ефекту. В обох випадках підвищення артеріального тиску є помірним, оскільки продовжують нормально діяти барорецептори дуги аорти. Якщо перерізати також аферентні волокна, що проходять від барорецепторів у складі блукаючого нерва, то артеріальний тиск підвищується до 300/200 мм рт. ст. або вище і є нестабільним. Двобічне ушкодження ЯПШ, місця закінчення барорецепторних аферентних волокон, зумовлює виражену гіпертензію, яка може бути летальною. Ці форми експериментальної гіпертензії називають нейрогенною гіпертензією.
Барорецептори передсердь, які реагують на розтягування
Є два типи рецепторів передсердь, що реагують на розтягування: одні, в яких імпульси виникають протягом систоли передсердь (тип А), інші, у яких імпульси виникають наприкінці діастоли, в момент максимального наповнення передсердь (тип В). Імпульсація від барорецепторів типу В посилюється зі збільшенням венозного повернення крові і послаблюється під впливом позитивного тиску під час дихання. Отже, ці барорецептори передусім реагують на розтягування стінок передсердь. Посилення імпульсації від більшості (якщо не всіх) цих рецепторів зумовлює рефлекторні зміни кровообігу і, зокрема, розширення судин та зниження артеріального тиску. Однак у цьому разі частота серцевих скорочень не збільшується, а зменшується.
Роль барорецепторів у гормональному регулюванні об’єму ПКР
Зі зменшенням об’єму ПКР знижується центральний венозний тиск, і послаблення імпульсації від барорецепторів передсердь зумовлює посилене виділення вазопресину (див. Розділ 14). Водночас посилюється активність симпатичного відділу, що призводить до посиленого продукування реніну. Виділення реніну спричинює продукування альдостерону. За значного зменшення об’єму ПКР простежується зниження артеріального тиску і послаблення імпульсації від барорецепторів сонної пазухи та дуги аорти, що теж сприяє посиленню виділення гормонів. Характеристика інших факторів, які посилюють продукування вазопресину і реніну, наведена у Розділах 20, 24 і 39. Кінцевим результатом описаних процесів є затримка води і натрію, спрямована на відновлення об’єму ПКР.
Рефлекс Бейнбріджа
Швидка інфузія крові або фізіологічного розчину анестезованим тваринам деколи зумовлює збільшення частоти серцевих скорочень, якщо попередньо частота серцевих скорочень була малою. Цей ефект описав Бейнбрідж 1915 р., відтоді він відомий як рефлекс Бейнбріджа. Вважають, що це справжній рефлекс, а не місцеве реагування на розтягування, оскільки він блокований двобічною ваготомією. Інфузія рідин тваринам з трансплантованим серцем збільшує частоту скорочення культі передсердь реципієнта, однак не впливає на частоту скорочень трансплантованого серця. У цьому, можливо, задіяні рецептори передсердь, які спричинюють тахікардію (описані раніше). Цей рефлекс протидіє опосередкованому барорецепторами зменшенню частоти серцевих скорочень у відповідь на збільшення об’єму. Рефлекс послаблюється або взагалі не простежується, якщо початкова частота серцевих скорочень була високою. Було багато дискусій про важливість цього рефлексу, однак його остаточна фізіологічна роль не з’ясована.
Рецептори лівого шлуночка
Унаслідок розтягування лівого шлуночка в експериментальних тварин відбувається зниження системного артеріального тиску і частоти серцевих скорочень. Розтягування шлуночка в цьому разі повинно бути значним, і вирішальна фізіологічна роль цієї реакції до кінця не з’ясована. Можливо, рецептори лівого шлуночка, що реагують на розтягування, відіграють роль у підтриманні тонусу блукаючого нерва, який забезпечує низьку частоту серцевих скорочень у стані спокою.
Якщо експериментальним тваринам уводити серотонін, вератридин, капсаїцин, фенілдигуанід і деякі інші речовини у вінцеві артерії, що постачають кров’ю лівий шлуночок серця, то розвивається апное, після чого настає часте дихання, гіпотензія і брадикардія (вінцевий хеморефлекс, або рефлекс Безольда-Яріша). Рецептори, очевидно, є закінченнями С-волокон, а еферентні волокна проходять у складі блукаючого нерва. Ця реакція не виникає після ін’єкцій у судини, що постачають кров’ю передсердя й правий шлуночок. Її фізіологічна роль остаточно нез’ясована. Можливо, у хворих з інфарктом міокарда речовини, що виділяються з ділянок інфаркту, стимулюють рецептори шлуночка, сприяючи гіпотензії, яка нерідко ускладнює це захворювання.
Легеневі рецептори
У випадку введення в легеневу артерію серотоніну, капсаїцину, вератридину та споріднених речовин відбувається активування закінчень С-волокон, що розташовані близько до капілярів легень, унаслідок чого розвивається апное, а потім настає часте дихання, гіпотензія та брадикардія (легеневий хеморефлекс). Цю реакцію (описана в Розділі 36) блокує ваготомія. За суттю вона аналогічна до вінцевого хеморефлексу, який виникає у разі введення речовин в артерії, що постачають кров’ю лівий шлуночок. Реакція розвивається занадто швидко - раніше, ніж речовини досягнуть рецепторів лівого шлуночка.
Клінічні дослідження і стимулювання
Зміни частоти пульсу і рівня артеріального тиску, що простежуються в людей, які встають або лягають (див. Розділ 33), відбувається насамперед за допомогою барорецепторних рефлексів. Про роль рецепторів свідчать зміни частоти серцевих скорочень у відповідь на підвищення артеріального тиску, спричинені введенням а-адреностимулятора фенілефрину. Нормальна відповідь відображена на рис. 31-13. За значень систолічного тиску від 120—150 мм рт. ст. є лінійна залежність між рівнем тиску і зменшенням частоти серцевих скорочень (збільшення інтервалу RR). Функцію рецепторів можна також підтвердити, вивчаючи зміни пульсу та артеріального тиску у відповідь на короткі періоди навантаження (форсоване видихання в разі закритої голосової щілини: маневр Вальсальви). На початку навантаження артеріальний тиск підвищується (рис. 31-14) тому, що підвищення тиску у грудній порожнині сприяє підвищенню тиску в аорті. Потім артеріальний тиск знижується, оскільки високий тиск у грудній порожнині зумовлює звуження вен, зменшення венозного повернення до серця і зменшення хвилинного об’єму. Зниження артеріального і пульсового тиску призводить до послаблення імпульсації від барорецепторів і спричинює тахікардію та збільшення периферійного опору судин. У випадку відкритої голосової щілини тиск у грудній порожнині нормалізується, хвилинний об’єм відновлюється, однак периферійні судини звужені. Простежується підвищення артеріального тиску понад нормальні показники, що призводить до стимулювання барорецепторів і зумовлює брадикардію та зниження артеріального тиску до нормального рівня.

Рис. 31-13. Опосередковане барорефлексом зменшення частоти серцевих скорочень у разі введення фенілефрину людині Зверніть увагу на те, що значення інтервалу RR на ЕКГ, відкладені на вертикальній осі, обернено пропорційні до частоти серцевих скорочень (відтворено за дозволом з Kotrly К et al: Effects of fentanyl-diazeparn-nitrous oxide anaesthesia on arterial baroreflex control of heart rate in man. Br J Anaesth 1986; 58:406).

Рис. 31-14. Діаграма відповіді на маневр Вальсальви у здорово! людини, записана за допомогою голки, введеної у плечову артерію (за згодою М Мсllrоу).
Описані зміни частоти серцевих скорочень простежуються також у хворих після симпатектомії, оскільки барорецептори і блукаючі нерви у них є інтактними. Однак у хворих з вегетативною недостатністю - синдромом, за якого виникає значне порушення функцій автономної нервової системи, - частота серцевих скорочень не змінюється. З незрозумілих причин частота серцевих скорочень і артеріальний тиск не змінюються також у разі нормалізації тиску в грудній порожнині у хворих з первинним гіперальдостеронізмом. Їхнє реагування на маневр Валь- сальви нормалізується після видалення пухлини, що виробляє альдостерон.
Вплив стимулювання хеморецепторів на вазомоторний центр
Аферентні волокна, що проходять від хеморецепторів каротидних та аортальних клубочків, передусім беруть участь у регулюванні дихання, їхня функція описана в Розділі 36. Проте ці волокна доходять також до вазомоторного центру. Вплив стимулювання хеморецепторів на серцево-судинну систему полягає у звуженні периферійних судин і брадикардії. Однак гіпоксія, крім цього, спричинює гіперпноє і посилене виділення катехоламінів із надниркових залоз, зміни, які, відповідно, зумовлюють тахікардію та збільшення хвилинного об’єму крові. У разі кровотечі розвивається гіпотензія, і це призводить до стимулювання хеморецепторів. Цей ефект пояснюють послабленням плину крові у ділянках локалізації хеморецепторів і застійною аноксією цих ділянок (див. Розділ 37). У тварин з гіпотензією імпульсація від барорецепторів послаблюється (див. нижче), і перерізання язикогорлового та блукаючого нервів зумовлює не підвищення, а зниження артеріального тиску. Це пояснюють тим, що нема впливу хеморецепторів на вазомоторний центр. Імпульсація від барорецепторів, імовірно, відіграє деяку роль і у виникненні хвиль Майєра. Ці хвилі не треба плутати з хвилями Траубе-Герінґа - коливанням артеріального тиску, що синхронізоване з диханням. Хвилі Майєра - це повільні регулярні коливання артеріального тиску з частотою одне за 20-40 с під час гіпотензії. За цих умов унаслідок гіпоксії відбувається стимулювання хеморецепторів, що, відповідно, веде до підвищення артеріального тиску. Наслідком підвищення артеріального тиску є посилення плину крові у ділянках локалізації хеморецепторів і послаблення імпульсації від них. Після зниження тиску починається новий цикл. Хвилі Майєра послаблюються, проте повністю не зникають у разі денервації хеморецепторів. Вони деколи простежуються у хребетних тварин. Отже, в розвитку цих хвиль певну роль відіграють спінальні вазопресорні рефлекси.
Прямі впливи на вазомоторний центр
Як гіпоксія, так і гіперкапнія стимулюють вазомоторний центр, діючи прямо на РВЛД, хоча прямий вплив гіпоксії виражається у незначному підвищенні внутрішньочерепного тиску, погіршенні кровопостачання вазомоторного центру, а місцева гіпоксія та гіперкапнія посилюють імпульсацію в ньому. Підвищення артеріального тиску, яке в цьому разі розвивається (рефлекс Кушінга), спрямоване на відновлення плину крові до довгастого мозку. Воно зумовлює рефлекторне зменшення частоти серцевих скорочень через барорецептори артерій (див. нижче). Саме тому у хворих з підвищеним внутрішньочерепним тиском характерний симптом не тахікардія, а брадикардія.
Таблиця 31-5. Фактори, які впливають на частоту серцевих скорочень1

1 Норадреналін має пряму хронотропну дію на серце. Проте в інтактних тварин його вазопресорна дія зумовлює стимулювання барорецепторів і рефлекторне посилення тонусу блукаючого нерва, достатнє для того, щоб перевищити пряму дію і спричинити брадикардію.
Підвищення артеріального РСО2 стимулює вазомоторний центр, однак прямим периферійним ефектом гіперкапнії є розширення судин. Отже, периферійний і центральний ефекти протидіють один одному. Помірна гіпервентиляція, яка суттєво зменшує напруження СО2 у крові, зумовлює звуження судин шкіри та мозку, артеріальний тиск у цьому випадку практично не змінюється. Під впливом високих концентрацій СО2 простежується виражене розширення судин шкіри та мозку, судини ж інших ділянок звужуються, і артеріальний тиск дещо підвищується.
Симпатична судинорозширювальна система
Холінергічні симпатичні судинорозширювальні волокна є частиною регулювальної системи, яка розпочинається у корі головного мозку, проходить до гіпоталамуса і середнього мозку, а звідти, не перериваючись, через довгастий мозок прямує до ПБ сірого стовпа спинного мозку. Прегангліонарні нейрони цієї системи активують постгангліонарні нейрони, які проходять до скелетних м’язів і анатомічно належать до симпатичних, проте виділяють ацетилхолін. Стимулювання цієї системи зумовлює розширення судин скелетних м’язів. Однак посилення плину крові, яке внаслідок цього виникає, супроводжується не збільшенням, а зменшенням споживання О2. Це свідчить про те, що кров тече в обхід капілярів. У випадку стимулювання цієї системи посилюється виділення адреналіну й норадреналіну з мозкової речовини надниркових залоз. Виділений адреналін посилює розширення судин скелетних м’язів. У собак і котів систему активує дія емоційних факторів, таких як страх, тривога і гнів. Її роль у людей детально не вивчена. Вважають, що симпатична судинорозширювальна система бере участь у розвитку запаморочення під дією емоцій. Є прямі докази того, що існує холінергічно опосередкована вазодилатація у скелетних м’язах протягом або навіть на початку фізичного навантаження (див. Розділ 33), хоча відомо, що розширення судин перед фізичним навантаженням не дуже виражене і простежується не завжди.
Регулювання частоти серцевих скорочень
Симпатична і парасимпатична іннервація серця, а також рефлекторні зміни, опосередковані барорецепторами, детально описано у попередніх розділах. Однак у табл. 31 - 5 наведено і систематизовано головні фактори, що впливають на частоту серцевих скорочень. Загалом, подразники, які збільшують частоту серцевих скорочень, одночасно підвищують також рівень артеріального тиску.
Фактори, які зменшують частоту серцевих скорочень, знижують артеріальний тиск, хоча є винятки, наприклад, гіпотензія і тахікардія в разі стимулювання барорецепторів передсердь; а також гіпертензія і брадикардія у випадку підвищення внутрішньочерепного тиску (див. вище).