Фізіологія людини - Вільям Ф. Ґанонґ 2002
Дихання
Регулювання дихання
Нервове регулювання дихання
Системи регулювання
Нервове регулювання дихання забезпечують дві окремі системи механізмів. Одна відповідає за довільне, а інша - за мимовільне (автоматичне) регулювання. Довільна система охоплює в себе структури, розташовані у корі великих півкуль, і надсилає імпульси у дихальні мотонейрони через кортикоспінальні провідні шляхи. Автоматична система міститься у мості та довгастому мозку, еферентні вихідні імпульси з неї прямують до дихальних мотонейронів через білу речовину, що розташована між латеральним і вентральним кортикоспінальними шляхами. Нервові волокна, завдяки яким розпочинається вдихання, конверґують на діафрагмальних мотонейронах, що розміщені у задніх рогах від СЗ до С5, а зовнішні міжреберні мотонейрони - у передніх рогах грудного відділу спинного мозку. Нервові волокна, що пов’язані з видиханням, конверґують перш за все на міжреберних мотонейронах у грудному відділі спинного мозку.
Мотонейрони, що йдуть до експіраторних дихальних м’язів, гальмуються, коли такі, що забезпечують інспіраторні м’язи, активні і навпаки. Хоча, спінальні рефлекси сприяють реципрокній іннервації (див. Розділ 6), та вона виникає загалом унаслідок активності в низхідних провідних шляхах. Імпульси у низхідних шляхах збуджують агоністи та гальмують антагоністи. Винятком у реципрокному гальмуванні є короткотривала незначна активність у діафрагмальних аксонах після вдихання. Значення такого постінспіраторного вихідного імпульсу полягає у перериванні еластичної тяги легень і створенні дихання плавним.
Система довгастого мозку
Ритмічна імпульсація нейронів довгастого мозку та мосту забезпечує автоматичність дихання. Перетинання стовбура мозку нижче від довгастого мозку (розтин D на рис. 36-1) припиняє дихання, а в разі рострального перетинання мосту (розтин А на рис. 36-1) автоматичне дихання є нормальним. Дихальні нейрони стовбура головного мозку бувають двох типів: одні виявляють активність під час вдихання (інспіраторні, І-нейрони), а інші - під час видихання (експіраторні, Е-нейрони). Більшість активної імпульсації посилює частоту під час вдихання, у випадку І-нейронів, або під час видихання у випадку Е-нейронів. Деяка імпульсація послаблює частоту, а інша імпульсація перебуває на однаковому високому рівні під час вдихання або видихання. Однак видихання є пасивним під час спокійного дихання, і Е-нейрони тоді перебувають у стані спокою; вони стають активними, якщо легенева вентиляція збільшується.
Ділянку довгастого мозку, пов’язану з диханням, називають дихальним центром; у ньому розрізняють дві окремі групи нейронів (див. рис. 36-1). Дорсальна група нейронів міститься у або біля ядра поодинокого шляху. Вентральна група - це довгий стовпець нейронів, що простягається через подвійне та задньоподвійне ядро у вентролатеральній частині довгастого мозку. Дорсальну групу, головно, утворюють І-нейрони, деякі з яких виступають і моносинаптично поєднуються з діафрагмальними мотонейронами. До них, вірогідно, проектуються від дихальних шляхів та каротидних і аортальних клубочків аферентні шляхи, які закінчуються у ядрі поодинокого шляху. Вентральна група містить Е-нейрони у каудальній частині дихального центру, І-нейрони - у його середній частині, та Е-нейрони - на ростральному кінці. Деякі з цих нейронів проникають у дихальні мотонейрони; ті ж, що містяться на ростральному кінці групи, відповідають за гальмування І-нейронів під час видихання.
Головні компоненти генераторного комплексу регулювання дихання, відповідального за автоматичність дихання, містяться у довгастому мозку, тому спонтанне дихання триває, хоча деколи нерегулярно і з затрудненями, після перетинання стовбура головного мозку по нижньому краю мосту (розтин С на рис. 36-1).
Ні дорсальна, ні вентральна групи дихальних нейронів не є обов’язковою частиною генераторного комплексу, оскільки вибіркове ураження однієї або іншої зменшує амплітуду дихання, однак не припиняє його. Хоча після цього дихання буде ритмічним, ініційованим невеликою групою синаптично поєднаних пейсмейкерних клітин у комплексі пре-Буттзінґера на одному з двох боків довгастого мозку між подвійним і латеральним ретикулярним ядром (рис. 36-2). Ці нейрони імпульсують ритмічно й утворюють ритмічну імпульсивну активність у діафрагмальних мотонейронах, яка зникає в разі перетинання між комплексом пре-Буттзінґера і цими мотонейронами.
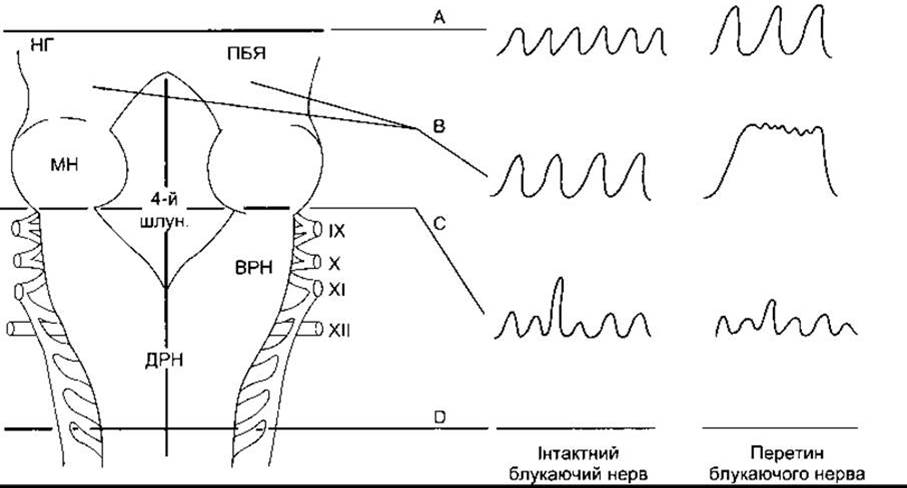
Рис. 36-1. Респіраторні нейрони у стовбурі головного мозку. Дорсальний вигляд стобура головного мозку; мозочок видалений. Зображені також впливи різноманітних уражень та перетини стовбура головного мозку. Записи спірометрії праворуч відображають глибину і частоту дихання. ДРН - дорсальна група респіраторних нейронів; ВРН - вентральна група респіраторних нейронів; ПБЯ - парабрахіальне ядро (пневмотаксичний центр); 4-й шлун. - четвертий шлуночок; НГ - нижній горбик, МН - середні мозкові ніжки. Римські цифри позначають черепні нерви (модифіковано та відтворено за дозволом з Mitchell RA, Berger A: State of the art: Review of newral regulation of respiration. Am Rev Respir Dis 1975; 111:206).
Впливи мосту і блукаючих нервів
Ритмічна імпульсація нейронів довгастого мозку стосується спонтанності дихання, проте її змінюють нейрони мосту й аферентні волокна блукаючого нерва від рецепторів у дихальних шляхах та легенях. У ділянці, відомій як пневмотаксичний центр, у медіальних і парабрахіальних ядрах та ядрі Куллікера-Фузе у дорсолатеральному мості містяться І- та Е-нейрони, і нейрони, активні у обох фазах дихання. Якщо зруйнувати цю ділянку (розтин В на рис. 36-1), то дихання стане повільнішим, а дихальний об’єм збільшиться; у разі перетинання блукаючого нерва в анестезованих тварин настає пролонгований інспіраторний спазм, подібний до затримки дихання. Дихання припиняється на фазі вдихання (апнейзис). Функція пневмотаксичного центру у нормі досі остаточно не з’ясована, однак він може відігравати роль перемикача між вдиханням та видиханням, регулятора оптимального співвідношення тривалості вдихання, видихання і дихальної паузи. Розтягнення легень під час вдихання ініціює імпульсацію в аферентних легеневих вагусних волокнах. Ці імпульси гальмують інспіраторну імпульсацію. Власне тому збільшується глибина вдихання після ваготомії (див. рис. 36- 1); апнейзис же виникає, якщо блукаючі нерви перетято після руйнування пневмотаксичного центру. Як зображено на рис. 36-3, активність вагусного зворотного зв’язку не змінює рівня зростання нейрональної активності у дихальних мотонейронах, проте без них активність триває.
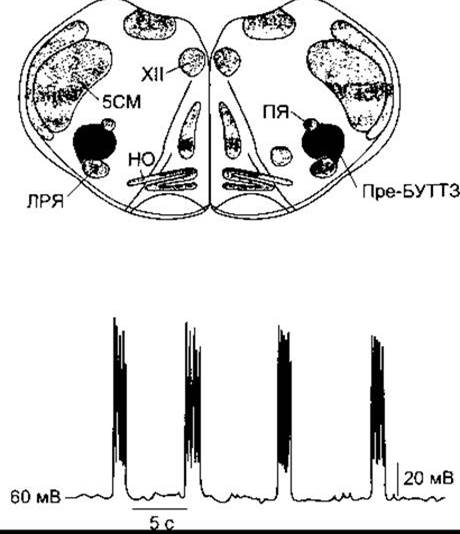
Рис. 36-2. Ритмічна імпульсація (унизу) нейронів комплексу пре-Буттзінґера (затемнена ділянка, пре-БУТТЗ) у препараті головного мозку щура в неонатальному періоді; НО - нижні оливи; ЛРЯ - латеральне ретикулярне ядро; ПЯ - подвійне ядро; XII - ядро дванадцятого черепного нерва; 5СМ - спинномозкове ядро трійчастого нерва (модифіковано з Smith JC et аі: Рrе-Вуttrіngеr complex: A brainstem region that may generate respiratory rhythm in mammals. Science 1991;254:726).
В інтактних тварин зі збільшенням активності інспіраторних нейронів частота і глибина дихання зростають. Глибина дихання збільшується, оскільки легені розтягуються до більшого ступеня, перш ніж потужність інгібіторної активності вагусного та пневмотаксичного центру стане достатньою для подолання інтенсивнішої імпульсації в інспіраторних нейронах. Частота дихання посилюється тому, що післяімпульсація у блукаючих нервах та, вірогідно, в пневмотаксичних аферентних волокнах до довгастого мозку швидко переважає.