Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Синаптичне і контактне передавання
Синаптичне передавання - Функціональна анатомія
Типи синапсів
Анатомічна будова синапсів у різних частинах нервової системи ссавців дуже різноманітна. Кінцеві відділи пресинаптичних нейронів у більшості випадків потовщені, тобто утворюють термінальні розширення (синаптичні пухирці) (див. рис. 4-1). Закінчення розміщені переважно на дендритах (рис. 4-2) і часто на дендритних шипах - невеликих синаптичних пухирцях, які виступають з дендритів (рис. 4-3). У деяких випадках терміналі аксонів пресинаптичних нейронів утворюють кошики, або сітки, навколо тіла постсинаптичної клітини (кошикові клітини мозочка й автономних вузлів). В інших ділянках вони переплітаються з дендритами постсинаптичної клітини (ліаноподібні волокна мозочка) або закінчуються безпосередньо на дендритах (верхівкові дендрити пірамідних клітин кори) чи аксонах (аксо-аксонні закінчення). У середньому кожний аксон утворює понад 2000 синаптичних закінчень. Оскільки в центральній нервовій системі (ЦНС) містяться 1011 нейронів, то кількість синапсів становить приблизно 2x1014. Без сумніву, кількість шляхів, якими може проходити нервовий імпульс за наявності такої складної мережі, надзвичайно велика.
В спинному мозку пресинаптичні закінчення тісно прилягають до тіл і проксимальних кінців дендритів постсинаптичних нейронів. Кількість синаптичних пухирців, що контактують з окремим мотонейроном спинного мозку, становить приблизно 10 000, у тому числі з тілом клітини - 2 000 і з дендритами - 8 000. Нейрон утворює настільки багато контактів, що він ніби інкрустований дендритними шишками. Площа мембрани нейрона, що контактує з окремим синаптичним пухирцем, невелика, проте ці пухирці настільки численні, що в цілому ділянка нейрона, яка контактує з ними, часто становить 40% від поверхні мембрани тіла нейрона і 75% від поверхні мембрани дендритів. Підраховано, що в корі великого мозку 98% усіх синапсів створюються з дендритами і лише 2% - з тілами клітин. Велика кількість збудливих закінчень у корі великого мозку і мозочка закінчуються на дендритних шипиках. Багато з них мають вужчі шийки, ніж головки, і їхні електричні властивості зумовлені співвідношенням ділянки головки до ділянки шийки. Обчислено, що співвідношення кількості синапсів до кількості нейронів у передньому мозку людини становить 40 000:1.
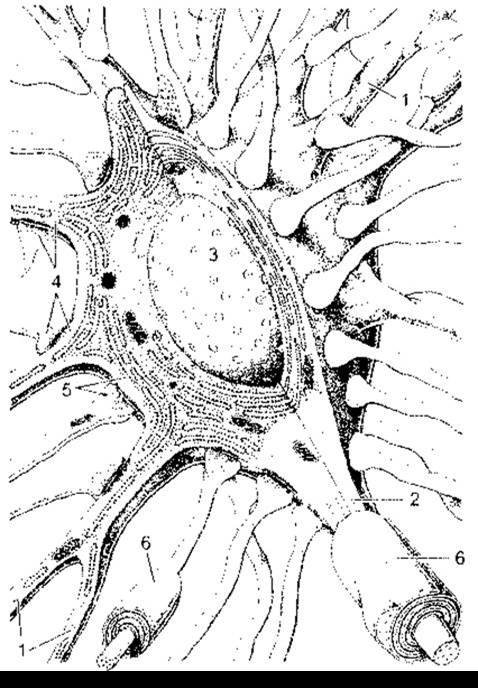
Рис. 4-1. Синапси на типовому мотонейроні. Нейрон має дендрити (1), аксон (2) і велике ядро (3). Зверніть увагу, що шорстка ендоплазматична сітка поширюється в дендрити і не поширюється в аксон. Багато різних аксонів сходяться до нейрона, і їхні термінальні розширення контактують з ним, утворюючи аксодендритні (4) й аксосоматичні (5) синапси; 6 - мієлінова оболонка (відтворено за дозволом з KrsticRV: Ultrastructure of the Mammalian Cell. Springer, 1979).
Пластичність дендритних шипів
Численні шипи містяться на дендритах не лише в корі великого мозку (див. рис. 4-3), а й у мозочку. Вони також є досить лабільними структурами. їхня кількість збільшується, наприклад, у складних умовах зовнішнього середовища in vivo, внаслідок подразнення ділянок морського коника (гіпокампа).
Морфологічні зміни в шипах можна виявити упродовж секунд; вони залежать від актину і міозину. Вірогідно теж, що принаймні в певних умовах ці зміни зумовлені спонтанним вивільненням пресинаптичними нейронами глютамату і його дією на АМПА-рецептори (див. нижче). Дендритні шипи підпадають і під вплив гормонів; наприклад, естрадіол сприяє ростові шипів у культурі клітин гіпокампа.

Рис. 4-2. Електронна мікрофотографія синаптичного пухирця (S), що контактує з дендритом (D) у центральній нервовій системі; Р - постсинаптичне потовщення; М - мітохондрія. (збільшення у 56 000 разів; з люб’язного дозволу DM McDonald).
Пре- та постсинаптичні структури і їхня функція
Між пре- і постсинаптичною частинами синапсу міститься синаптична щілина 20-40 нм завширшки. Вздовж синаптичної щілини в постсинаптичній мембрані є багато нейротрансмітерних рецепторів і постсинаптичні потовщення (див. рис. 4-2,4-4). Скупчення рецепторів зумовлені зв’язувальними білками, які утримують рецептори і відповідні йонні канали.
Усередині пресинаптичної частини є велика кількість мітохондрій, а також багато вкритих мембраною синаптичних пухирців, що містять нейротрансмітери. Налічують три типи синаптичних пухирців: маленькі прозорі синаптичні пухирці, що містять ацетилхолін, гліцин, ГАМК або глютамін (див. нижче); маленькі електроннощільні пухирці, що містять катехоламіни; великі електроннощільні пухирці, що містять нейропептиди. Пухирці і білки, що містяться в них, утворюються в комплексі Ґольджі тіла нервової клітини і мігрують аксоном до закінчень завдяки швидкому аксоплазматичному транспортуванню. Нейропептиди, що містяться у великих електроннощільних пухирцях, теж утворюються в тілі клітини. Однак маленькі прозорі і маленькі електроннощільні пухирці рециклізуються в закінченнях. Вони там наповнюються нейротрансмітером, унаслідок екзоцитозу виділяють свій вміст у синаптичну щілину, внаслідок ендоцитозу повертаються в ендосоми, згодом відділяються від ендосом, наповнюються нейротрансмітером, і цей цикл повторюється знову (рис. 4-5).

Рис. 4-3. Шипи верхівкових дендритів великих пірамідних нейронів кори великого мозку людини. Зверніть увагу, що кількість шипів швидко збільшується від народження до восьмимісячного віку. У випадку синдрому Дауна шипи тонкі і невеликі (модифіковано з Shepherd GM: Neurobiology, 2nd ed. Oxford Univ Press, 1988).
Зазначимо, що великі електроннощільні пухирці містяться всюди в пресинаптичній частині, утвореній термінальною гілочкою аксона, і виділяють свої нейропептиди внаслідок екзоцитозу в усіх ділянках терміналі. Маленькі пухирці, локалізовані поблизу синаптичної щілини, зливаються з мембраною і виділяють свій вміст дуже швидко в синаптичну щілину в ділянках синаптичних потовщень, які називають активними зонами (див. рис. 4-4). Активні зони містять ряди кальцієвих каналів.
Молекула Са2+ є ключовою в процесах злиття синаптичних пухирців з пресинаптичною мембраною і виділення трансмітера в синаптичну щілину. Потенціал дії, що надходить до закінчення пресинаптичного нейрона, відкриває потенціалозалежні Са2+-канали, і надходження Са2+ зумовлює вивільнення трансмітера. Концентрація Са2+ усталюється на початковому рівні внаслідок їхнього швидкого видалення з клітини, головно внаслідок Ca2+-Na+ антипорту.
Як зазначено в Розділі 1, утворення пухирців, їхнє злиття з мембраною, виділення їхнього вмісту і подальша рецик- лізація їхньої мембрани є фундаментальними процесами, що відбуваються в усіх клітинах. У комплексі Ґольджі і клітинній мембрані вони відбуваються внаслідок екзо- і ендоцитозу. Такого типу механізми мають свої особливості в конкретних умовах. Ці тонко регульовані процеси, що стосуються виділення вмісту синаптичних пухирців і їхньої рециклізації, відображені нарис. 4-5. Деталі процесу, завдяки якому синаптичні пухирці зливаються з клітинною мембраною усе ще з’ясовують, проте відомо, що до цього процесу причетний v-петлеподібний білок мембрани пухирців - синаптобревін, що зв’язується з t-петлеподібним білком клітинної мембрани - синтаксином (рис. 4-6).
Клінічний інтерес становить факт, що деякі смертоносні токсини, які блокують виділення нейротрансмітера - це цинквмісні ендопептидази, які розщеплюють і, отже, інактивують білки комплексу, що забезпечує процеси злиття й екзоцитозу. Правцевий токсин і ботулінічні токсини В, D, F і G діють на синаптобревін, ботулінічний токсин С - на синтаксин, а ботулінічні токсини А та В - на SNAP-25 (див. рис. 4-6). Клінічно правцевий токсин спричинює спастичний параліч, блокуючи виділення в ЦНС пресинаптичного трансмітера, а в разі ботулізму внаслідок блокування виділення в місці нервово-м’язового сполучення ацетилхоліну виникає млявий параліч. Позитивне значення має те, що місцеве введення невеликих доз ботулінічного токсину ефективне в лікуванні широкого спектра хвороб, що супроводжуються м’язовою гіперактивністю. Прикладом може слугувати ін’єкція в нижній стравохідний сфінктер з метою полегшення стану, що виникає в разі ахалазії, або ж ін’єкція в мімічні м’язи для усунення зморшок.
Конверґенція і диверґенція
Лише в окремих випадках синаптичні пухирці, що контактують з постсинаптичним нейроном, є закінченням певного одного пресинаптичного нейрона. Переважно ж вплив на клітину множинний. Наприклад, на спинномозковому мотонейроні закінчуються нейрони заднього корінця, нейрони довгих низхідних спинномозкових шляхів і велика кількість інтернейронів - коротких проміжних нейронів спинного мозку. Отже, багато пресинаптичних нейронів конверґують, тобто сходяться, на кожному поодинокому постсинаптичному нейроні. І навпаки, аксони більшості пресинаптичних нейронів розділені на велику кількість гілок, що диверґують, закінчуючись на багатьох постсинаптичних нейронах. Конверґенція і диверґенція - анатомічні субстрати процесів підсилення, оклюзії і реверберації (див. нижче).
Однонапрямлене передавання
Синапси забезпечують проходження нервових імпульсів лише в одному напрямі - від пресинаптичного нейрона до постсинаптичного. Імпульс, що його передає аксон переднього корінця антидромно, загасає після деполяризації в тілах клітин спинномозкових мотонейронів. Оскільки аксони можуть передавати нервовий імпульс у будь-якому напрямі, то однонапрямлений процес у синапсах необхідний для впорядкування функцій нервової системи. Хімічне передавання в синаптичних контактах забезпечує такий односторонній процес.
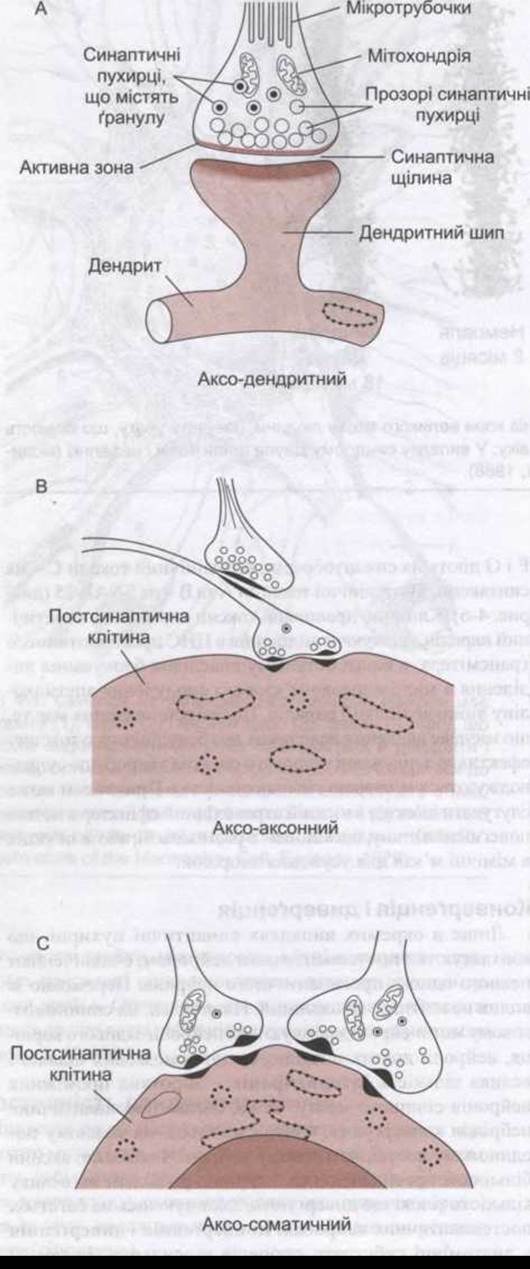
Рис. 4-4. Аксо-дендритний, аксо-аксонний і аксо-соматичний синапси. Багато пресинаптичних нейронів закінчуються на дендритних шипах, як показано вгорі, проте деякі - безпосередньо на тілах дендритів. Зверніть увагу на наявність у закінченнях прозорих синаптичних пухирців, а також на скупчення прозорих пухирців біля активної зони. А - поздовжній переріз; В, С - поперечний переріз.
Хімічний трансмітер міститься в синаптичних пухирцях пресинаптичних волокон, проте його нема в постсинаптичних мембранах. Тому імпульс, що надходить антидромно до постсинаптичної мембрани, не зумовлює вивільнення синаптичного трансмітера. Поширення імпульсу можливе лише за умови, що потенціал дії досягає пресинаптичних закінчень і зумовлює виділення накопиченого в них хімічного трансмітера.
Розвиток синапсів
Цікавим є питання про те, як протягом розвитку нейрони знаходять один одного і налагоджують відповідні синап- тичні контакти. Під час росту організму на верхівках аксонів містяться конуси наростання, що забезпечують міграцію крізь тканини. Ці конуси діють за участі атрактантів і репелентів тканин і, якщо нема принаймні однієї з цих речовин, то аксон відмирає. Деталі розвитку в цій книзі не описані. Однак треба звернути увагу на важливість у цьому процесі білків, які називають семифоринами. Окремі представники цієї великої родини тісно пов’язані з трансмембранними доменами, тоді як інші здатні до переміщення. В хребетних описано понад 20 семифоринів. Репеленти відштовхують конуси наростання, а атрактанти, навпаки, притягують їх. Проте вплив цих білків залежить теж від концентрації вторинних месенджерів, що містяться в конусі наростання, який перебуває в ділянці впливу семифоринів. Рецептори для семифоринів називають нейрофілінами. Крім того, багато нейронів налагоджують синаптичні контакти з великою кількістю інших нейронів або скелетними м’язами, надалі невідповідні контакти ліквідуються і залишаються лише ті, які згодом функціонують у дорослому організмі. Багато нейронів відмирають під час розвитку (апоптоз; див. Розділ 1), залишаються ж лише найактивніші нейрони і їхні синаптичні контакти. Між нейронами є також певна конкуренція за утворення синаптичних сайтів, і нервові закінчення сусідніх нейронів вростають у ділянки, що позбавлені іннервації.