Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Слух та рівновага
Волоскові клітини
Будова
Усі волоскові клітини внутрішнього вуха мають подібну будову (рис. 9-8). Кожна волоскова клітина розміщена в оточенні опорних, або підтримувальних, клітин. За винятком зовнішніх волоскових клітин завитки, базальна частина цих клітин охоплена дендритами аферентних нейронів. Від апікальної поверхні волоскових клітин відходить 30-150 паличкоподібних відростків, або волосків. За винятком завитки, один з цих відростків - кіноцилія - є справжньою, проте нерухомою війкою, що містить дев’ять периферійних і одну центральну пару мікротрубочок (див. Розділ 1). Це один із найдовших відростків з булавоподібним розширенням на кінці. Кіноцилії втрачаються волосковими клітинами завитки у дорослих ссавців. Інший тип відростків - стереоцилії - наявні в усіх без винятку волоскових клітинах. Вони мають серцевину, утворену паралельними актиновими філаментами, що вкриті різноманітними ізо-формами міозину. Сукупність відростків кожної волоскової клітини є впорядкованою: висота стереоцилій поступово збільшується у напрямі до кіноцилії; у перпендикулярному напрямі, в межах одного і того ж ряду, стереоцилії мають однакову висоту.
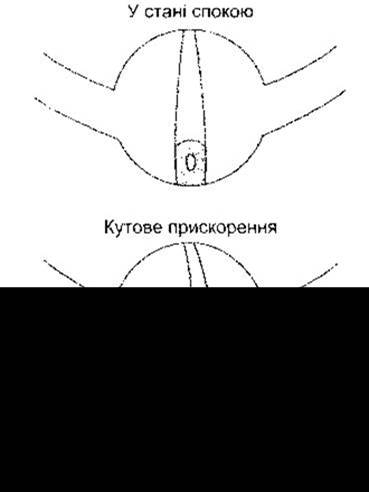
Рис. 9-6. Схематичне відтворення ампульних гребенів. Купол на верхівці гребеня перекриває ампулу і може згинатися. Унаслідок інерційності ендолімфа під час кутового прискорення зміщується у напрямі, протилежному до напряму обертання. Це зумовлює згинання відростків і, як наслідок, зміну проникності та мембранного потенціалу волоскових клітин.
Електричні відповіді
Мембранний потенціал волоскових клітин у спокої становить близько - 60 мВ. Коли стереоцилії рухаються у напрямі до кіноцилії, то мембранний потенціал зменшується до - 50 мВ. Зі зміщенням стереоцилій у протилежному напрямі клітини гіперполяризуються. Зміщення відростків у напрямі, перпендикулярному до означеної осі, не зумовлює змін мембранного потенціалу. Зміщення у косому напрямі спричинює де- або гіперполяризацію волоскових клітин, пропорційно та залежно від того, до чи від кіноцилії рухаються стереоцилії. Отже, волоскоподібні відростки реалізують механізм трансформації напряму і розміру відхилень у зміни мембранного потенціалу волоскових клітин.
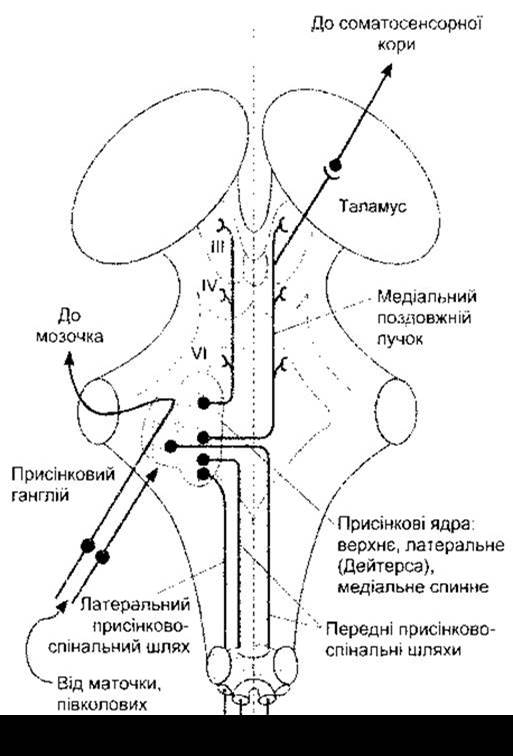
Рис. 9-7. Головні присінкові шляхи мозкового стовбура, вигляд ззаду. Мозочок та кора мозку видалені.
Походження потенціалів дії в аферентних нервових волокнах
Як зазначено вище, відростки волоскових клітин оточені ендолімфою, тоді як їхню базальну частину омиває пери- лімфа. Таке співвідношення потрібне для нормального утворення потенціалів дії. Перилімфа утворюється щонайбільш з плазми крові. Хоча транспортування манітолу та сахарози з плазми до перилімфи барабанних сходів сповільнене порівняно з транспортуванням цих речовин до перилімфи вестибулярних сходів та є інші дрібні відмінності між рідинами у цих двох порожнинах, проте обидва різновиди перилімфи нагадують позаклітинну рідину. Ендолімфа, яку продукує судинна смужка, має високу концентрацію К+ і низьку концентрацію Na+ (рис. 9-9). Для клітин судинної смужки характерний високий рівень Na+- К+-АТФ-ази. Окрім того, цим клітинам притаманна унікальна потенціалозалежна К+-помпа, яка забезпечує різницю потенціалів, що дорівнює 85 мВ, між середніми та присінковими або барабанними сходами.
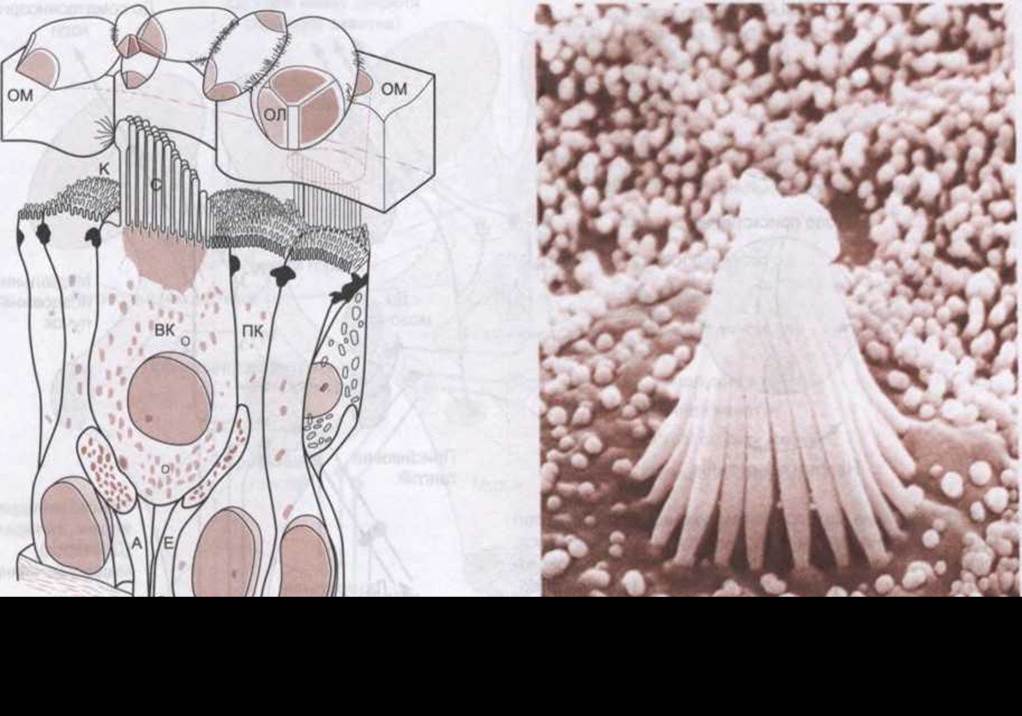
Рис. 9-8. Ліворуч: структура волоскової клітини мішечка жаби, яка відображає її взаємозв'язок із отолітовою мембраною (ОМ); К - кіноцилія; С - стереоцилія; ВК - волоскова клітина з аферентними (А) та еферентними (Е) нервовими волокнами; ОЛ - отоліт; ПК - підтримувальна клітина (відтворено за дозволом з Hillman DE: Morfology of peripheral and central vestibular systems. Linas R, Precht W [editors]: Frog Neurobiology. Springer, 1976). Праворуч: сканована електронна мікрофотографія відростків волоскової клітини плями мішечка жаби. Отолітова мембрана видалена. Короткі відростки навколо волоскової клітини - мікроворсинки підтримувальних клітин (AJ Hudspeth).
Дуже тонкі відростки, так звані верхівкові з’єднання (рис. 9-10), сполучають верхівку кожної стереоцилії з бічною поверхнею сусідньої вищої стереоцилії, яка у ділянці цього контакту містить чутливі до механічних подразнень катіонні канали. Коли нижчі стереоцилії зміщуються у напрямі до вищих, то час перебування цих каналів у відкритому стані збільшується. Вважають, що стан напруження кожного каналу регулює «адаптаційний мотор», утворений міозином вищої стереоцилії. Зміщення стереоцилій у протилежному напрямі зменшує час перебування каналу у відкритому стані. Функцію цього каналу може виконувати а-субодиниця Na+-каналу епітеліоцитів, яка сама по собі утворює низькоселективний катіонний канал (див. Розділ 1), а амілорид приєднаний до ділянки контакту між коротшою та довшою стереоциліями. Означені канали є порівняно низькоспецифічними катіонними каналами, та оскільки їх омиває ендолімфа з підвищеним вмістом йонів К+, то йони проникають всередину волоскової клітини через відкриті канали, зумовлюючи деполяризацію мембрани. Проникнення Ca2+ зумовлює викидання нейротрансмітера, який спричинює деполяризацію мембрани одного або кількох аферентних нейронів, що контактують з волосковою клітиною. Хоча нейротрансмітер до цього часу не ідентифіковано, правдоподібно, що ним може бути глютамат.
Йони К+, що проникають усередину волоскової клітини через механочутливі катіонні канали, рециклізують (див. рис. 9-9). Спершу вони потрапляють до підтримувальної клітини, звідки через щільні контакти дифундують до інших підтримувальних клітин. Нарешті вони досягають судинної смужки, звідки, завершуючи цикл, відбувається їхня секреція до ендолімфи.