ОСНОВЫ МИКРОБИОЛОГИИ - В. М. Самыгин - 2015
ГЛАВА 4. МОРФОЛОГИЯ МИКРООРГАНИЗМОВ
Все известные одноклеточные и многоклеточные организмы по морфологическому признаку делятся на две большие группы - прокариоты и эукариоты. К прокариотам относятся бактерии и цианобактерии, к эукариотам - грибы, водоросли, простейшие, зеленые растения и животные. Клетки прокариот не имеют оформленного ядра. Иными словами, генетический материал (ДНК) прокариот находится прямо в цитоплазме и не окружен ядерной мембраной. У эукариот (греч. eu - истинный, karion - ядро) имеется настоящее ядро, то есть у них генетический материал окружен двойной мембраной (ядерной оболочкой) и образует вполне определенную клеточную структуру, которую очень легко узнать.
4.1. Методы изучения морфологии микроорганизмов
Исследованиями микробной клетки занимаются цитология, цитохимия, биохимия, биофизика, молекулярная биология, генетика, микроскопия и другие научные дисциплины. Они изыскивают различные методы, стараясь «заглянуть» внутрь клетки, изучить ее строение и связанные с ним жизненные процессы.
Основным методом изучения морфологии микробной клетки является микроскопия, то есть наблюдение за живыми или мертвыми клетками с помощью микроскопа. Вначале это был световой микроскоп, позднее появились фазово-контрастный, стереоскопический, ультрафиолетовый, люминесцентный, поляризационный и, наконец, электронный микроскопы.
Световые микроскопы подразделяются на студенческие, рабочие, лабораторные и исследовательские, различающиеся по конструкции и комплектации.
Микробиологическое исследование начинается обычно с микроскопии нативного материала (биологический материал, продукты питания и др.). Результаты микроскопии в подавляющем большинстве случаев недостаточны для идентификации микроорганизма, но дают возможность получить ориентировочное представление о количестве содержащихся в материале микроорганизмов и их принадлежности к палочковидной или кокковой микрофлоре. Для определения формы микроорганизмов с помощью светового микроскопа чаще всего используют препараты фиксированных (убитых) клеток, которые окрашивают различными красителями, подразделяемые по их способности выявлять определенные структурные образования микробных клеток. Обычно различают 4 большие группы красителей:
- основные (или ядерные) красители, избирательно окрашивающие ядро и базофильные (от лат. basis - основной) структуры бактериальных клеток;
- кислые (или цитоплазматические) красители, которые окрашивают преимущественно цитоплазму, реже клеточные стенки;
- нейтральные красители, избирательно окрашивающие отдельные компоненты цитоплазмы, например, Судан III или нильский синий окрашивают капельки жира;
- флюорохромы - группа красителей, способных флюоресцировать при той или иной длине возбуждающего света.
Существуют методы окраски у микробов ядерного вещества (по Романовскому-Гимзе) спор (методы Ожешко, Пешкова), капсул (метод Бурри), клеточной стенки (метод Гутштейна, Циля-Нильсена), жгутиков (метод Морозова), зерен волютина (полифосфата, метод Раскиной).
Уже в XIX веке ученые установили, что разрешающая способность оптических микроскопов ограничена длиной световой волны. Самые мелкие бактерии находятся около предела видимости наиболее совершенного оптического микроскопа. Фазово-контрастный микроскоп не преодолел нижней границы наблюдаемых размеров, но с его помощью можно наблюдать живые клетки микроорганизмов. В фазово-контрастном микроскопе благодаря изменению длины пути световых волн возникает «фазовый сдвиг на одну четвертую длины волны». В результате усиливается рельеф, что позволяет увидеть некоторые малые элементы структуры клеток.
Наибольшие возможности для морфологических исследований дает электронный микроскоп. Роль световых лучей, благодаря которым в других микроскопах получается увеличенное изображение наблюдаемых объектов, в электронном микроскопе играют пучки электронов. Их движением управляют электромагниты, выполняющие функцию оптических линз. Современный электронный микроскоп дает возможность получать увеличение объекта в несколько сот тысяч раз. Для исследования внутреннего строения клеток используется особый микрохирургический аппарат - ультрамикротом. Он позволяет получать сверхтонкие срезы клеток (0,02 мкм) для просмотра в электронном микроскопе. Однако у электронного микроскопа есть один недостаток - в нем можно наблюдать лишь мертвые клетки. Это связано с тем, что молекулы воздуха представляют для электронов непреодолимое препятствие, поэтому все наблюдения должны проводиться в вакууме, а это приводит к немедленному обезвоживанию и гибели всех живых клеток.
4.2. Морфологические типы бактерий
Для характеристики размеров микробных клеток используют следующие измерения: микрометр (1 мкм = 10-6 м), нанометр (1 нм = 10-9м). При работе с электронным микроскопом используется еще более мелкая единица - ангстрем (А), или 10-10м. Большинство клеток бактерий имеет в среднем величину 1-3 мкм, клетки дрожжевых грибов достигают 5-10 мкм. Вполне понятно, что такие маленькие существа должны быть и необычайно легкими. На один мг веса приходится 5 млрд бактерий, т. е. 1 клетка весит ~0,2 • 109 мг.
По внешнему виду различают три основные формы бактерий - шаровидную, или кокковидную (греч. coccus - ягода), палочковидную, или цилиндрическую, и извитую, или спиралевидную.
Кокки бывают сферические, элипсовидные, бобовидные и ланцетовидные (обоюдоострые лезвия). В зависимости от расположения отдельных клеток по отношению друг к другу кокки делятся на следующие группы:
- микрококки - характеризуются одиночным, беспорядочным расположением клеток (рис. 1, а);
- диплококки - парные кокки (пневмококк - возбудитель пневмонии, менингококк - возбудитель менингита) (рис. 1, б);
- стрептококки - располагаются в виде цепочек (виды, вызывающие сепсис, воспалительные процессы) (рис. 1, в);
- стафилококки - располагаются в виде гроздьев винограда (вызывают фурункулы, абсцессы и так далее) (рис. 1, г);
- тетракокки - состоят из четырех близко лежащих кокков (рис. 1, д);
- сарцины - располагаются пакетообразно по 8-16 и более клеток (рис. 1, е).
Рис. 1. Форма сферических бактериальных клеток

Палочковидные (цилиндрические) формы подразделяют на бактерии, бациллы и клостродии. К бактериям относятся такие палочковидные микроорганизмы, которые, как правило, не образуют спор (кишечная, брюшнотифозная, дизентерийная, туберкулезная и другие). К бациллам (лат. bacillus - палочка) и клостридиям (лат. closter - веретено) принадлежат микробы, в большинстве образующие споры (сенная, сибиреязвенная, столбнячная палочки, возбудители анаэробных инфекций) (рис. 2).
Рис. 2. Палочковидные формы микроорганизмов: а - кишечные палочки б - сибиреязвенные бациллы в - клостридии ботулизма

Общее число палочковидных бактерий значительно больше, чем кокковидных.
Извитые, или спиралевидные формы. К этой группе бактерий относятся (рис 3):
- вибрионы (лат. vibrio - изгибаюсь) - клетки, имеющие вид запятой (возбудитель холеры, водные вибрионы);
- спириллы (лат. spira - изгиб) - извитые формы бактерий, имеющие изгибы с одним или несколькими оборотами спирали;
- спирохеты имеют штопорообразную извитую форму.
Рис. 3. Извитые (спиралевидные) формы микроорганизмов: а - вибрионы: б - спириллы: в - спирохеты
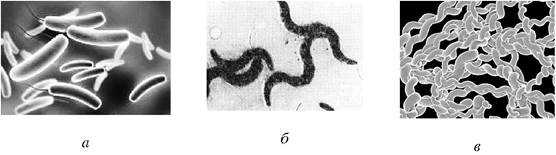
Обнаружены также бактерии, имеющие треугольную, нитевидную, звездообразную и другие формы.
Микробы обладают полиморфизмом - индивидуальной изменчивостью, при которой наблюдается разнообразие их форм и размеров, проявляющиеся независимо от возраста и стадии развития. Они легко подвергаются изменениям под влиянием различных факторов: температуры, питательной среды, концентрации солей, кислотности, продуктов метаболизма, дезинфектантов, лекарственных препаратов и др. Особенно часто проявляется полиморфизм у бактерий при культивировании их на искусственных средах. Такие изменения морфологии бактерий связаны с нарушением либо синтеза оболочки, либо механизма регуляции клеточного деления. В зависимости от силы и глубины воздействия на микробную клетку изменения могут быть наследственными и ненаследственными.
4.3. Ультраструктура бактериальной клетки
Строение типичной бактериальной клетки показано на рис. 4.
Рис. 4. Схематическое изображение прокариотической клетки: 1 -цитоплазма, 2 - цитоплазматическая мембрана, 3 - клеточная стенка, 4 - капсула, 5 - жгутик, б - пили (ворсинки) 7 - нуклеоид, 8 - плазмиды, 9 - мезосома, 10 - рибосомы, 11 - включения
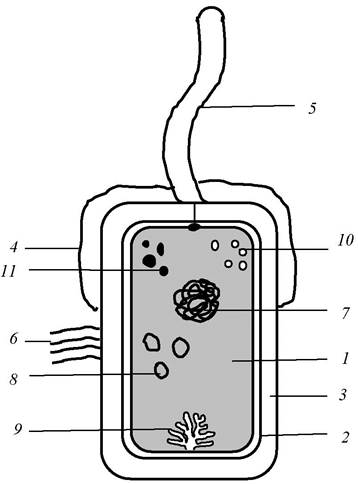
Основные структуры микробной клетки - оболочка, цитоплазма, ядерный аппарат (нуклеоид) и различные цитоплазматические включения.
Оболочка бактерий состоит из цитоплазматической мембраны, клеточной стенки и капсульного слоя, превращающегося у некоторых видов в истинную капсулу.
Под влиянием различных факторов среды некоторые микробы обладают способностью откладывать на поверхности мощный слизистый слой вокруг клеточной стенки, получивший название капсулы. В зависимости от плотности и толщины слизистого слоя различают макро- микрокапсулы, слизистый чехол (гликокаликс) и S-слои:
- макрокапсула (собственно капсула);
- слизистый чехол (гликокаликс) так же, как и капсула, образован высокополимерными мукополисахаридами и отличается от микрокапсулы тем, что не имеет постоянной прочной связи с клеточной стенкой;
- S-слои выстилают наружную поверхность клеточной стенки равномерно упакованными белковыми структурами правильной формы. Они имеют упаковку и расположены подобно паркетным дощечкам, плотно прикрывающим клетку.
Капсульное вещество бактерий состоит из полисахаридов, мукополисахаридов (гиалуроновой кислоты) или полипептидов. В зависимости от особенностей микроорганизмов и состава питательной среды химический состав капсул весьма вариабелен. Благодаря высокому содержанию воды (до 98 %) они слабо окрашиваются красителями. Капсулообразование считается приспособительной функцией микробов. Капсулы защищают клетку от действия токсических веществ, механических повреждений и высыхания. Патогенные капсульные микробы более устойчивы к фагоцитозу, действию защитных факторов организма. Капсулы не являются необходимой частью клетки. Несмотря на то, что капсулообразование является видовым признаком, оно может быть утрачено как в естественных, так и в искусственных условиях.
Жгутики. Многие бактерии подвижны, на основании чего их подразделяют на плавающие и скользящие. Подвижность плавающих бактерий (энтеробактерий, псевдомонад, спирилл) обусловлена наличием у них одного или нескольких жгутиков. Жгутики устроены в виде микротрубочек и состоят из одинаковых сферических субъединиц белка флагеллина, которые расположены по спирали и образуют полый цилиндр. Длина жгутиков 10-20 мкм, толщина 12-20 нм.
По расположению жгутиков подвижные микробы подразделяются на четыре группы: монотрихи - бактерии с одним жгутиком на конце (холерный вибрион, синегнойная палочка); лофотрихи - бактерии, имеющие по пучку жгутиков на одном конце; амфитрихи - бактерии с двумя полярно расположенными жгутиками или имеющие по пучку жгутиков на обоих концах; перитрихи - покрытые жгутиками по всей поверхности тела (кишечная палочка, сальмонеллы брюшного тифа) (рис. 5).
Рис. 5. Расположение жгутиков у бактерий: а - монотрихи: б - лофотрихи: в - амфитрихи: г - перитрихи
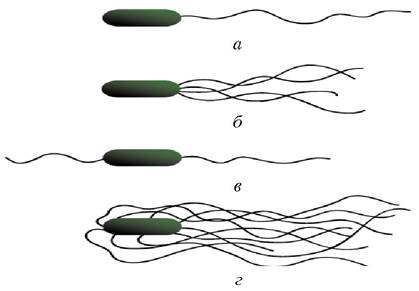
Движение клеток осуществляется путем вращения жгутика по или против часовой стрелки (у монотрихов), при этом сама клетка медленно вращается по часовой стрелке. При вращении жгутика по часовой стрелке клетка тянется за жгутиком вперед (жгутик впереди). При вращении жгутика против часовой стрелки клетка выталкивается жгутиком вперед (жгутик сзади). Некоторые монотрихи вращают жгутик только по часовой стрелке, и для смены направления им надо остановиться и переориентироваться. Перитрихи вращают жгутики (пучок жгутиков) против часовой стрелки (жгутики остаются сзади клетки по направлению движения). Наибольшей подвижностью обладают монотрихи (до 60 мкм/с).
Все подвижные бактерии передвигаются в направлении, определяемом внешними воздействиями. В зависимости от природы внешнего фактора, под воздействием которого осуществляется движение, различают хемотаксис (движение относительно того или иного химического соединения), аэротаксис (связан с разницей содержания в среде кислорода), магнетотаксис (способность двигаться по силовым линиям магнитного поля Земли), вискозитаксис (реакция бактерий на изменение вязкости раствора).
Еще одним способом движения бактерий является скольжение клеток по твердой поверхности в результате их волнообразных сокращений. Механизм этого типа движения изучен недостаточно. В некоторых случаях скольжение осуществляется за счет белков внешней мембраны и движение вперед по поверхности представляет собой вид отталкивания. Скользящие бактерии не имеют жгутиков и при движении по поверхности оставляют слизистый след. У некоторых бактерий описано «прыгающее» движение, природа которого не выяснена.
Пили. У различных видов микробов имеются тонкие выросты в виде прямых нитей - пили (ворсинки, или фимбрии). Они короче и тоньше жгутиков, покрывают все тело клетки и обнаружены как у подвижных, так и неподвижных организмов. Количество их от 50 до 400 штук на одной особи. Длина фимбрии составляет 1,5 мкм, диаметр - 7 нм. Пили способствуют прикреплению микробных клеток друг к другу или к какой-нибудь поверхности. Другой тип пилей, так называемые половые фимбрии, или F-пили, имеют внутри канал, через который передается генетический материал от одной клетки к другой при конъюгации бактерий. Пили состоят из белка пилина и не являются, как и жгутики, обязательными структурами клеток бактерий.
Клеточная стенка является чрезвычайно важным элементом структуры бактерий. Она придает клетке определенную форму и жесткость, защищает внутренне содержимое клетки, препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Ей принадлежит важная роль в регуляции роста и делении бактерий. Вода, другие малые молекулы, ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот. Толщина клеточной стенки колеблется от 20 до 100 нм и составляет около 20 % сухого вещества бактериальной клетки.
По строению клеточной стенки бактерии делят на две группы. Одни окрашиваются по Граму, поэтому их называют грамположительными, а другие обесцвечиваются при отмывке красителя и их называют грамотрицательными. В клеточной стенке и тех и других есть особая жесткая решетка, состоящая из муреина, или пептидогликана. Молекула муреина представляет собой правильную сеть из параллельно расположенных полисахаридных цепей, сшитых друг с другом короткими цепями пептидов. У грамположительных бактерий пептидогликан составляет 40-60 % сухой массы клеточной стенки. У этих микроорганизмов в муреиновую сетку встроены дополнительные структуры, образованные полисахаридами, белками или гликопротеидами. У грамотрицательных бактерий клеточная стенка тоньше, но устроена она сложнее. Муреиновый слой у этих бактерий составляет около 10 % массы клеточной стенки, снаружи покрыт гладким слоем липидов, образующих так называемую наружную мембрану, состоящую из фосфолипидов, липополисахарида, липопротеида и белков. Липидный слой придает клетке устойчивость к ферментам и антибиотикам (пенициллину). Под наружной мембраной расположена периплазма, или перизматическое пространство. Периплазма содержит слой пептидогликана, специфические для нее белки и олигосахариды, а также неорганические соединения. Бактерии, полностью лишенные клеточной стенки, называются протопластами, они имеют шаровидную форму, обладают способностью к делению, дыханию, синтезу белков и спорообразованию. Однако протопласты очень чувствительны к изменениям осмотического давления, механическим воздействиям и аэрации, не обладают активной подвижностью. Если происходит частичное растворение клеточной стенки, то бактерии превращаются в сферические тела, называемые сферопластами.
Цитоплазматическая мембрана прилегает к внутренней поверхности стенки клетки, ее толщина составляет 7-8 нм. Цитоплазматическая мембрана (ЦПМ) служит главным барьером между цитоплазмой клетки и окружающей внешней средой. Ключевым компонентом биологических мембран, представляющих бислойные структуры, являются полярные липиды, в основном фосфолипиды, содержащие жирные кислоты. ЦПМ выполняет функцию разделительной перегородки, через нее с помощью ферментов непрерывно осуществляется активный транспорт различных веществ и ионов, необходимых для жизнедеятельности клетки. У некоторых бактерий плазматическая мембрана впячивается внутрь клетки и образует мезосомы.
Мезосомы - складчатые мембранные структуры, состоящие из трубочек, пузырьков и пластинок, на поверхности которых находятся ферменты, участвующие в процессе дыхания, т. е. выполняют функцию генерирования энергии. Наряду с дыхательными ферментами и механизмами регуляции проницаемости, мезосомы бактерий совместно с ЦПМ содержат специфические ферментные системы, участвующие в процессах азотфиксации и хемосинтеза. Кроме того, мезосомы играют определенную роль в образовании клеточных стенок, делении, выделении экзоферментов, спорообразовании и т. д.
Генетический аппарат прокариот (хромосома или нуклеоид), в котором содержится генетическая информация относительно всех признаков, присущих клетке, представлен гигантской двухспиральной молекулой ДНК длиной 1 мм, толщиной около 2 нм. Нуклеоид представлен замкнутой структурой, не отграничен от цитоплазмы какой-либо мембраной и не связан с белками бактериальной клетки. При быстром росте бактерий деление хромосомы обычно опережает деление самой клетки, поэтому молодые клетки могут содержать несколько идентичных копий одной хромосомы. Например, клетки кишечной палочки при быстром росте на полноценной питательной среде содержат до 11 нуклеоидов, в медленно растущих клетках обнаруживают 1-2 хромосомы.
Большинство бактерий несет все гены на одной хромосоме. Однако появляются свидетельства того, что важные гены могут располагаться и на разных хромосомах. У некоторых вибрионов, бруцелл, лептоспир и др. обнаружены две кольцевые хромосомы, у Burkholderia cepacia - от двух до четырех. Более того, число видов, имеющих более одной группы сцепления генов (несколько хромосом), может оказаться гораздо больше, чем мы представляем сегодня, поскольку, согласно последним данным молекулярной биологии, нам известно менее 1 % от всех существующих в мире микроорганизмов.
У многих бактерий, помимо основной хромосомы, обнаружены дополнительные внеядерные (внехромосомные) кольцевые и линейные двухцепочечные молекулы ДНК, которые могут существовать и реплицироваться как независимо от хромосомы, так и быть интегрированными в нее. К ним относятся плазмиды, транспозоны и инсерционные (IS)последовательности. Плазмиды бактерий представляют собой двунитевые молекулы ДНК размером от 106 до 108 Д, несущие от 40 до 50 генов. Они выполняют регуляторные и кодирующие функции. Количество плазмидной ДНК составляет обычно не более нескольких процентов от хромосомной ДНК, а число плазмид от 1 до 200. Плазмиды содержат от 1500 до 40000 пар нуклеотидов. Плазмиды, как и другие внехромосомные элементы, не являются жизненно необходимыми, так как не кодируют информацию о синтезе ферментов, участвующих в метаболизме бактериальной клетки. Однако они могут изменять питательные потребности бактерий, придавать клетке такие свойства, как устойчивость к антибиотикам, солям тяжелых металлов, способность к продукции факторов патогенности, метаболических ферментов и др. Важная роль внехромосомных генетических элементов заключается в том, что они обеспечивают обмен генетическим материалом по горизонтали (от одной клетки популяции к другой) и играют определенную роль в эволюции прокариот. Таким образом, внехромосомные элементы делают возможным существование бактерий в более широком диапазоне условий окружающей среды, то есть действуют как фактор адаптации.
Цитоплазма бактерий представляет собой содержимое клетки, окруженное ЦПМ. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых веществ - белков, РНК, продуктов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена различными структурными элементами - генетическим аппаратом, рибосомами и различающимися по химической природе и функциям включениями. Цитоплазма характеризуется высоким показателем вязкости (соответствует вязкости глицерина или густого сиропа). Строение цитоплазмы зависит от возраста, питательной среды, вида бактерий. Цитоплазма старых бактерий становится более вязкой, напоминая по своей консистенции гель.
Цитоплазма содержит мелкие зерна диаметром 15-20 нм, состоящие из белка и нуклеиновых кислот (РНК), получившие название рибосом. Они являются обязательными и весьма важными структурами для жизнедеятельности бактерий. Количество рибосом в клетке колеблется от 5000 до 90000. Общая масса рибосом может составлять до четверти клеточной массы. Они являются местом белкового синтеза.
Рибосомы прокариот имеют константу седиментации 70S. Они построены из двух субъединиц: 30Sи 50S, 30S частица содержит одну молекулу 16S-рРНК и в большинстве случаев по одной молекуле белка более 20 видов. 50S-субъединица состоит из двух молекул рРНК (23S и 5S). В ее состав входит более 30 различных белков, также представленных одной копией. Большая часть рибосомальных белков выполняет структурную функцию.
Включения. В ходе жизнедеятельности бактериальной клетки в ее цитоплазме можно наблюдать морфологически дифференцированные образования, различающиеся по форме, размерам, химической природе и неодинаковы у разных бактерий. Их называют включениями. К бактериальным включениям относятся крупинки гликогена, крахмала, капли жира, серы, кристаллы щавелевой кислоты, зерна волютина, карбоната кальция, газовые вакуоли (аэросомы) и так далее. В некоторых случаях включение является скоплением продуктов обмена, в других - запасом питательных веществ. Разнообразные включения не являются необходимыми структурами для жизнедеятельности клетки и могут присутствовать либо отсутствовать у различных бактерий.
4.4. Споры и спорообразования
Микроорганизмы родов Bacillus, Clostridium, Desulfotomaculum, Sporolactobacillus, Sporosarcina и некоторых других (всего около 20 родов) способны образовывать споры(эндоспоры), представляющие собой тельца круглой или овальной формы. Спорообразование присуще как патогенным (возбудители сибирской язвы, ботулизма столбняка и др.), так и сапрофитным видам, обитающим в почве, воде, организме животных. Чаще всего споры образуют палочковидные микроорганизмы, сравнительно редко наблюдается образование спор у кокков и извитых форм.
Споры располагаются в клетках по-разному. Диаметр споры может быть приблизительно равен или несколько превышать диаметр клетки, из которой образовалась спора. У одних бактерий спора формируется на конце клетки (терминальное расположение), последняя при этом несколько расширяется, приобретая вид барабанной палочки; у других образуется в центре клетки, и последняя либо не меняет форму (род Bacillus), либо расширяется в центре, принимая вид веретена (род Clostridium) (рис. 6).
Рис. 6. Расположение эндоспор в бактериальной клетке: 1, 2 - центральное; 3, 4, 5 - терминальное; 6 - латеральное
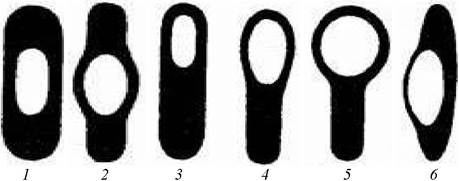
Споры характеризуются большой преломляемостью света и при микроскопическом исследовании в неокрашенном состоянии имеют вид блестящих зерен; они с трудом прокрашиваются красителями. Вследствие наличия плотной многослойной оболочки, имеющей пластинчатое строение, минимального количества свободной воды и высокого содержания кальция и липидов споры обладают повышенной устойчивостью к действию факторов внешней среды и могут длительно (десятки лет) сохраняться в неблагоприятных условиях. Споры некоторых бацилл выдерживают кипячение и действие высоких концентраций дезинфицирующих веществ. Они погибают в автоклаве от воздействия, насыщенного пара при температуре 115-125 °С (1,5-2 атм) спустя 20 минут, а также под влиянием температуры 150-170 °С в сухожаровом шкафу (1 час).
Продукция спор - это одна из стадий цикла развития определенных микроорганизмов, выработанная в процессе эволюции в борьбе за сохранение вида. Споры представляют стадию покоя. При попадании в неблагоприятные условия окружающей среды цитоплазма с генетическим материалом (одна или несколько хромосом) обособляется от остального клеточного материала за счет впячивания цитоплазматической мембраны внутрь клетки. Мембрана нарастает от периферии к центру, где срастается, что приводит к образованию споровой поперечной перегородки. Затем происходит образование проспоры - структуры, расположенной внутри материнской клетки и полностью отделенной от нее наружной и внутренней мембранами. Поверх наружной мембраны проспоры синтезируются споровые покровы. После сформирования споры происходит лизис (разрушение) материнской клеточной стенки и спора выходит в окружающую среду. Процесс образования споры длится 18-20 часов. Обычно в клетке закладывается одна спора, но известны случаи формирования до 5 спор в одной клетке. Спорообразование нельзя считать способом размножения бактерий, поскольку при формировании спор увеличения числа микроорганизмов не происходит.
Попадая в благоприятные условия, споры прорастают и превращаются снова в вегетативные формы. При этом они набухают, увеличиваются в размерах, содержание воды в них возрастает, процессы обмена усиливаются. На первом этапе происходит активация ферментов, идет активный синтез белка и РНК, но репликация ДНК начинается лишь через 1-2 часа после начала прорастания споры. В последующем происходит разрушение толстого слоя молекул пептидогликана, расположенного между двумя мембранами проспоры, разрыв споровых покровов и выход сформировавшейся к этому времени структуры, называемой ростовой трубочкой. В дальнейшем наблюдается ее удлинение, достраивание клееночной стенки и деление удлиненной клетки. Обычно прорастание идет значительно быстрее, чем образование споры (в течение 4-5 часов).
Способность к спорообразованию используют в систематике микробов, однако она может быть утрачена в результате частых пересевов на питательных средах или культивирования при высокой температуре.
Помимо эндоспор, у некоторых других бактерий существуют иные покоящиеся формы - экзоспоры и цисты. Экзоспоры возникают путем почкования материнской клетки, они сходны по своим свойствам с эндоспорами бацилл. Некоторые бактерии образуют шарообразные толстостенные клетки, называемые цистами, которые характерны для азотобактера, спирохет, миксобактерий, риккетсий, метилотрофных и некоторых других бактерий. При истощении пищевых ресурсов в цисту превращается вся палочковидная вегетативная клетка, а не часть ее, как при образовании эндоспоры. Цисты устойчивы к высушиванию, механическим нагрузкам и облучению, но не к действию высоких температур.