БИОТЕХНОЛОГИЯ РАСТЕНИЙ И БИОБЕЗОПАСНОСТЬ - А. П. Ермишин - 2015
ГЛАВА 7. ПОЛУЧЕНИЕ ГАПЛОИДОВ И МАНИПУЛЯЦИИ С ПЛОИДНОСТЬЮ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ СЕЛЕКЦИИ РАСТЕНИЙ
Гаплоидами у покрытосеменных растений называют особи, имеющие гаметический набор хромосом по отношению к уровню плоидности родительской особи. Удвоение хромосом у гаплоидов диплоидных видов (получение удвоенных гаплоидов) позволяет получать гомозиготные линии, которые представляют значительный интерес для селекции.
Гаплоиды могут возникать естественным путем в результате нарушения процесса оплодотворения из неоплодотворенных яйцеклеток или других клеток зародышевого мешка, при элиминации хромосом одного из родителей в зиготе или замещении ядра яйцеклетки ядром спермия. Первым растением, которое было идентифицировано в качестве гаплоида и выявлено A. F. Blakeslee в 1921 г., был дурман Datura stramonium (Solanacea). Позже гаплоиды были обнаружены и у многих других растений, однако частота их появления в природе оказалась крайне незначительной (0,001-0,01 %) и не могла удовлетворить потребности генетики и селекции.
Разработка методов культуры клеток растений in vitro дала возможность получать в эксперименте гаплоиды и удвоенные гаплоиды с частотой, намного превышающей естественный уровень. Для многих видов растений разработаны эффективные технологии, позволяющие использовать гаплоиды и манипуляции с плоидностью для ускорения селекционного процесса, снижения его трудоемкости.
В настоящее время для редукции числа хромосом у растений применяют два подхода, в основе которых лежит фундаментальное свойство тотипотентности половых клеток. Первый из них предполагает получение гаплоидных растений-регенерантов путем андрогенеза (от греч. andros - мужчина и genesis - зарождение, происхождение) в культуре in vitro пыльников или микроспор, второй - путем гиногенеза (от греч. gyne - женщина и genesis - зарождение, происхождение) из неоплодотворенных яйцеклеток или других гаплоидных клеток зародышевого мешка в культуре in vitro, а также из имеющих гаплоидный зародыш семян, полученных при опылении материнского растения пыльцой другого вида или специального гаплоиндуктора. В последнем случае появление гаплоидного зародыша связано с индуцированным чужеродной пыльцой развитием неоплодотворенной яйцеклетки (партеногенеза, от греч. parthenos - девушка и genesis - зарождение, происхождение) либо с избирательной элиминацией в зиготе хромосом опылителя или материнского растения.
7.1. Культура in vitro пыльников и микроспор
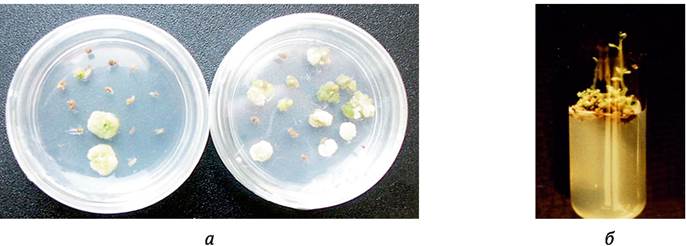
Высшие растения имеют жизненный цикл с чередующимися фазами гаплоидного гаметофита и диплоидного спорофита. Во время фазы гаметофита материнские клетки пыльцы, находящиеся в пыльнике, претерпевают мейотическое и митотическое деления, результатом которых является образование мужского гаметофита - пыльцевых зерен. Под воздействием определенных факторов в условиях культуры пыльников in vitro некоторые микроспоры способны менять программу развития с гаметофитной на спорофитную и давать в результате начало эмбриоидам (рис. 7.1) или образовывать каллюс (рис. 7.2). Из эмбриоидов или каллюса можно получить растения-регенеранты - гаплоиды или удвоенные гаплоиды. Это явление получило название андрогенеза или пыльцевого эмбриогенеза (pollen embryogenesis, microspore embryogenesis). В русскоязычной литературе для его обозначения также используется термин «андроклиния».
Рис. 7.1. Пыльцевой эмбриогенез в культуре пыльников перца Capsicum annuum L.: а, б - высвобождение эмбриоида из пыльника через 30 дней культуры: сначала появляются корешки (а), затем зародыш полностью (б); в - проросток пыльцевого происхождения с семядолями; 2 - проросток с листьями (по J. Barany и др., 2005)

Рис. 7.2. Культура пыльников картофеля in vitro: a - образование каллюса из пыльников; б - стеблевой органогенез из каллюса
Возможность получения гаплоидов путем культивирования in vitro пыльников растений впервые была продемонстрирована в 1964 г. сотрудниками Отдела ботаники университета г. Дели (Индия) S. Guha и S. C. Maheswari. Они обнаружили формирование эмбриоидов в культуре пыльников дурмана Datura innoxia, которую собирались использовать для изучения биохимии мейоза. Позднее ими было установлено, что эмбриоиды происходили из незрелых пыльцевых зерен (микроспор) и имели гаплоидное число хромосом (S. Guha, C. Maheswari, 1966).
В 1973 г. французские ученые С. Nitsch и В. Norreel впервые получили гаплоидные эмбриоиды путем культивирования in vitro микроспор Datura innoxia, изолированных из пыльников.
Первоначально для получения культуры микроспор пыльники вскрывали скальпелем и их содержимое высвобождалось в жидкую питательную среду. Однако эта процедура была весьма трудоемкой, а выход микроспор был невысоким. В дальнейшем этот метод был усовершенствован: для лучшего извлечения микроспор применили магнитную мешалку. Кардинальное улучшение эффективности выделения микроспор было достигнуто благодаря использованию механической гомогенизации бутонов, содержащих пыльники с микроспорами на определенной стадии, пропусканию гомогената через сито, центрифугированию в градиенте плотности. Например, эффективная технология выделения микроспор пшеницы для культуры in vitro включает в себя следующие этапы (M. Zheng, 2003):
✵ колоски, погруженные в питательную среду, измельчают с помощью блендера. Важно подобрать оптимальную скорость вращения ножа блендера и продолжительность гомогенизации;
✵ полученный гомогенат фильтруют последовательно через сетчатые фильтры с размером ячеек 100 цм и 38 цм, чтобы отделить микроспоры от грубых остатков тканей и клеток;
✵ микроспоры, собранные на фильтре с размерами ячеек 38 μм, смывают раствором 0,3 М маннитола, наслаивают на 0,58 М раствор мальтозы и центрифугируют при небольшой скорости (100-150 g). Жизнеспособные эмбриогенные микроспоры располагаются на границе этих двух растворов, в то время как другие клетки и примеси оказываются в осадке. Фракцию микроспор несложно собрать с помощью автоматической пипетки и перенести в жидкую питательную среду для культивирования.
Несмотря на то что культура пыльников растений представляется более простой технологией для использования в селекции по сравнению с культурой микроспор, последняя имеет ряд преимуществ. Применение культуры микроспор позволяет существенно сократить трудозатраты, связанные с получением андрогенетических гаплоидов, так как процедура извлечения пыльников из бутонов у многих видов растений весьма сложная. Исключается возможность получения растений-регенерантов не из микроспор, а из соматических клеток, устраняется негативное воздействие на развитие микроспор окружающих тканей пыльника (правда, во многих случаях эти ткани могут оказывать и положительное влияние на развитие эмбриоидов). Культура микроспор является удобной модельной системой для изучения механизмов андрогенеза, проведения исследований, связанных с использованием экспериментального мутагенеза и генно-инженерных модификаций.
7.2. Особенности морфогенеза в культуре пыльников и микроспор
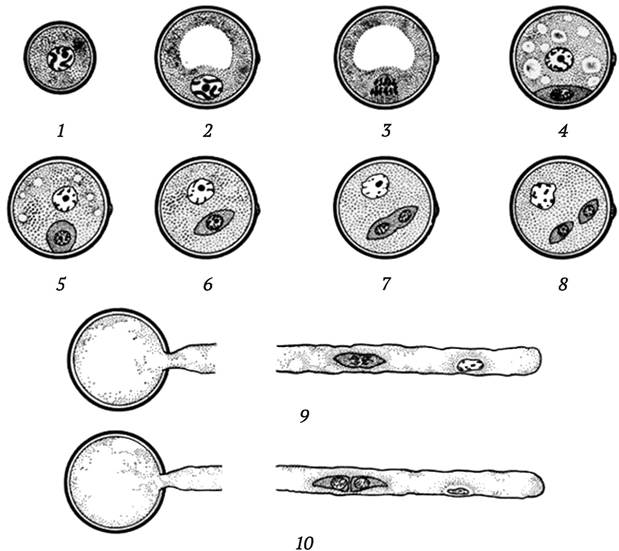
В ходе естественного формирования пыльцевых зерен (гаметофитный путь развития) у высших растений после мейотического деления материнской клетки пыльцы образуется четыре гаплоидные клетки (тетрада микроспор), которые высвобождаются из тетрады в полость пыльника после деградации межспоровых оболочек. Микроспоры первоначально имеют расположенное по центру ядро и большое количество мелких вакуолей в цитоплазме. Перед первым митотическим делением вакуоли сливаются в одну большую вакуоль, а ядро смещается к периферии клетки, т. е. происходит поляризация микроспор, в результате чего при митотическом делении образуются две клетки разного размера: большая вегетативная и маленькая генеративная (рис. 7.3, 1-4).
Рис. 7.3. Развитие мужского гаметофита у высших растений: 1 - микроспора после выхода из тетрады; 2 - образование вакуоли; 3 - деление ядра микроспоры; 4 - образование генеративной (внизу) и вегетативной клеток; 5 - генеративная клетка отходит от оболочки пыльцевого зерна; 6 - генеративная клетка расположена свободно в цитоплазме вегетативной клетки; 7 и 8 - деление генеративной клетки в пыльцевом зерне; 9 и 10 - деление генеративной клетки в пыльцевой трубке (по N. Maheswari, 1950)
Вегетативная клетка получает основную часть цитоплазмы, органелл, РНК и белков микроспоры. Ее ядро находится в деконденсированном состоянии, она содержит в цитоплазме большое количество рибосом и других органелл, наблюдается высокая транскрипционная активность. В ходе интенсивного роста вегетативной клетки вакуоль постепенно уменьшается и исчезает, транскрипционная активность падает, в пластидах накапливается большое количество крахмальных зерен, являющихся источником энергии для развития мужского гаметофита после прорастания пыльцевых зерен.
Генеративная клетка находится внутри цитоплазмы вегетативной клетки, ее ядро в конденсированном состоянии, метаболически неактивно. У нее мало цитоплазмы и всего несколько органелл. Она претерпевает одно симметричное митотическое деление, в результате которого образуется два гаплоидных ядра удлиненной формы - спермии. У некоторых видов растений деление генеративной клетки происходит в пыльцевой трубке после прорастания пыльцевого зерна на рыльце пестика (рис. 7.3, 5-10). Один из спермиев оплодотворяет яйцеклетку (образуется диплоидная зигота, в результате деления которой формируется зародыш), а второй - сливается с ядром центральной клетки зародышевого мешка, что дает начало триплоидному эндосперму.
Изменение описанной выше программы развития с гаметофитной на спорофитную в культуре in vitro происходит только в том случае, когда микроспоры находятся на определенной стадии развития и являются чувствительными (компетентными) к различным внешним воздействиям, способным вызвать такое изменение программы. Например, для большинства злаков оптимальная фаза для индукции пыльцевого эмбриогенеза - это стадия сильно вакуолизированной микроспоры (средняя и поздняя одноядерные микроспоры).
Имеется достаточно много экспериментальных данных, указывающих на связь между андрогенетической способностью генотипа и формированием микроспор определенного типа (эмбриогенных). Было показано, что число эмбриогенных микроспор детерминировано для конкретных растений-доноров пыльников и не увеличивается в ходе культивирования пыльников, а условия культивирования лишь способствуют или препятствуют реализации их развития. Теория, которая считает приоритетной роль эмбриогенных микроспор в процессах андрогенеза в культуре пыльников и микроспор, получила название теории детерминации (E. Heberle-Bors, 1985).
Эмбриогенные микроспоры имеют существенные ультраструктурные и функциональные отличия от нормальных микроспор. Для них характерно смещение вакуоли от центра к экзине на вакуолярной стадии развития микроспор и последующая реадсорбция вакуоли на двуядерной стадии с сохранением ее видимых фрагментов возле экзины. В результате формируется специфическая так называемая «звездчатая структура» (рис. 7.4), состоящая из цитоплазматических тяжей, соединяющих перинуклеарную и субкортикальную области цитоплазмы. Отмечается менее плотная цитоплазма, отсутствие накопления крахмальных зерен, характерное для нормальных созревающих генеративных клеток, происходит изменение клеточного цикла. В отличие от нормальных, у эмбриогенных микроспор хроматин вегетативного и генеративного ядер имеет одинаковый уровень конденсации, что говорит об одинаковой транскрипционной активности обоих ядер. Было обнаружено образование в эмбриогенных микроспорах в ходе культивирования in vitro специфической мРНК, не присутствующей в нормальных микроспорах, что говорит о транскрипции у них особых генов, связанных с развитием по спорофитному пути.
Рис. 7.4. «Звездчатая структура», появляющаяся у культивируемых in vitro микроспор пшеницы после воздействия стрессового фактора (по U. Bal и др., 2012)
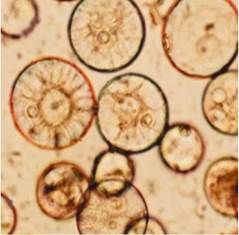
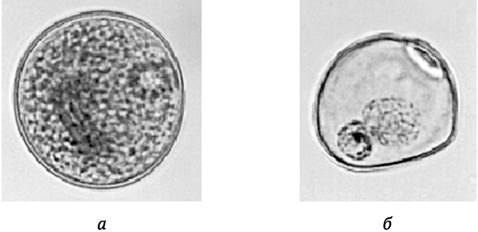
Пыльцевой диморфизм (присутствие в созревающей пыльце нормальных и эмбриогенных микроспор) проявляется и на уровне зрелой пыльцы (M. Horner, H. Street, 1978). Нормальные пыльцевые зерна содержат много крахмала, интенсивно окрашиваются ацетокармином (вегетативное и генеративное ядра не просматриваются). Пыльцевые зерна, происходящие от эмбриогенных микроспор, по размеру меньше нормальных, имеют слабое окрашивание цитоплазмы ацетокармином, у них отсутствуют крахмальные зерна; они содержат четко различимые вегетативное и генеративное ядра или одно ядро, или два одинаковых ядра (рис. 7.5). Такие зерна получили название р-зерна (от англ. pre-mitotic). Благодаря пыльцевому диморфизму имеется возможность проводить предварительную оценку андрогенетической способности генотипов на основе цитологических данных изучения препаратов зрелой пыльцы, оценивать эффективность воздействия на растения-доноры пыльников тех или иных средовых факторов до начала культивирования пыльников или микроспор.
Рис. 7.5. Пыльцевой диморфизм у пшеницы: а - нормальное пыльцевое зерно, окрашенное ацетокармином, с тремя ядрами и большим количеством крахмальных гранул; б - р-зерно, меньшее по размеру, слабо окрашенное ацетокармином, содержит вегетативное и генеративное ядра, крахмальные гранулы отсутствуют (по M. F. Grando, M. J. Moraes-Fernandes, 1997)
Изучение поведения микроспор в процессе культивирования in vitro показало, что большинство из них дегенерирует в течение нескольких дней после помещения пыльников на питательную среду. Лишь эмбриогенные микроспоры претерпевают последовательные митотические деления, давая начало новообразованиям (каллюсу или эмбриоидам). Выявлено несколько путей формирования эмбриоидов или каллюса из эмбриогенных микроспор (T. Aionesei и др., 2005).
Е-путь. Микроспора подвергается первому неравному митотическому делению, в дальнейшем происходят симметричные деления генеративной клетки (рис. 7.6, IV). В результате образуются гаплоидные эмбриоиды или каллюс. Впервые этот механизм был описан у культуры пыльников белены черной Hyasciamus niger (V. Raghavan, 1976), позднее и у других видов (ячменя, кукурузы, пшеницы, риса). Возможно также развитие многоклеточных структур, представляющих собой химеру из клеток, происходящих от деления как генеративной, так и вегетативной клеток.
Рис. 7.6. Пути формирования эмбриоидов или каллюса из эмбриогенных микроспор: I - одноядерная микроспора; II - микроспора, претерпевшая неравное митотическое деление (образовались генеративная и вегетативная клетки); III - микроспора, претерпевшая симметричное митотическое деление (образовались две одинаковые вегетативные клетки); IV - митотические деления только генеративной клетки (Е-путь); V - митотические деления только вегетативной клетки (А-путь); VI - митотические деления вегетативной и генеративной клеток (С-путь); VII - митотические деления вегетативных клеток, образованных после симметричного первого митотического деления микроспоры (В-путь)
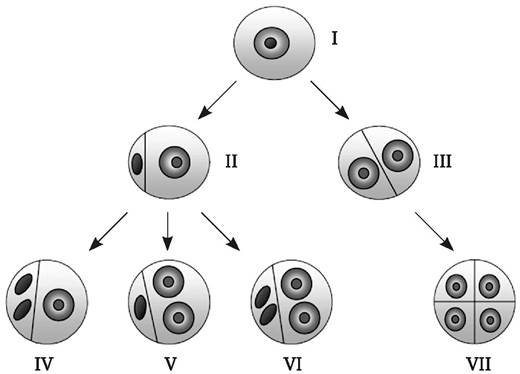
A-путь. Микроспора подвергается первому неравному митотическому делению. В дальнейшем происходят симметричные деления только вегетативной клетки, в результате чего образуются гаплоидные эмбриоиды. Генеративная клетка не делится и погибает (рис. 7.6, V). Этот механизм впервые описан у культуры пыльников табака (N. Sunderland, F. Wicks, 1971) и был преобладающим в случае культивирования пыльников, содержащих молодые двуядерные микроспоры. Образование эмбриоидов или каллюса из вегетативной клетки позднее было продемонстрировано и у ряда других видов (рапс, паслен, дурман, люфа и др.), при этом у некоторых из видов (ячменя, пшеницы, кукурузы, риса и ржи) отмечено формирование каллюса.
С-путь. Микроспора претерпевает нормальное митотическое деление, в результате которого формируются вегетативное и генеративное ядра. Затем ядра сливаются, последующие митотические деления приводят к образованию многоклеточных структур, которые в дальнейшем развиваются посредством эмбриоидогенеза или каллюсогенеза, давая начало преимущественно удвоенным гаплоидам (рис. 7.6, VI). При слиянии эндоредуплицированных ядер могут формироваться эмбриоиды, имеющие большую плоидность, однако они погибают на ранних стадиях развития.
В-путь. Первое митотическое деление, в отличие от нормального, является симметричным, что приводит к формированию двух одинаковых вегетативных клеток. Их последующее митотическое деление ведет к образованию многоклеточных структур (рис. 7.6, VII). Этот механизм преобладает при образовании гаплоидных эмбриоидов у дурмана Datura innoxia и рапса Brassica napus. Однако он обнаружен и у других видов, особенно в случае использования стрессовых воздействий на культивируемые микроспоры. Предполагается, что одна из причин симметричного деления - это отсутствие или формирование слаборазвитой вакуоли при первом делении микроспоры. Это может происходить у некоторых видов растений под действием температурного стресса (типичный случай - холодовое воздействие на одноядерные микроспоры дурмана Datura innoxia). При отсутствии вакуоли не происходит смещение ядра к периферии клетки, нарушается ее полярность. При В-пути развития микроспор клеточная оболочка после деления ядер, как правило, не формируется, ее закладка происходит после образования многоядерных структур. В связи с этим может происходить слияние равных ядер и увеличение плоидности (спонтанное удвоение хромосом). Развитие по В-пути обычно приводит к формированию гаплоидных или диплоидных эмбриоидов (удвоенных гаплоидов).
Таким образом, в результате развития эмбриогенных микроспор по одному из этих путей могут образовываться как гаплоидные, так и диплоидные эмбриоиды (удвоенные гаплоиды). Во многих случаях в культуре пыльников или микроспор образуются не эмбриоиды, а каллюс, из которого также могут быть регенерированы и гаплоидные, и диплоидные растения.
Считается, что получение удвоенных гаплоидов связано со спонтанным удвоением хромосом. Частота появления удвоенных гаплоидов может колебаться в значительных пределах в зависимости от вида растений, использованных генотипов, методов культивирования микроспор. Например, частота спонтанного удвоения хромосом варьировала от 0 до 21 % у различных генотипов кукурузы, от 10 до 40 % - у рапса Brassica napus. У некоторых сортов ячменя она достигала 87 %.
Механизм спонтанного удвоения хромосом в процессе пыльцевого эмбриогенеза в большинстве случаев неясен. В литературе описаны примеры эндоредупликации (удвоения ДНК без митоза), эндомитоза (митоза без образования веретен деления и разрушения оболочки ядра), слияния ядер. Тем не менее появление спонтанных удвоенных гаплоидов имеет важное практическое значение, поскольку предоставляется возможность получать гомозиготные линии, избежав трудозатрат, связанных с искусственным удвоением хромосом у гаплоидов.
7.3. Факторы, оказывающие влияние на эффективность андрогенеза в культуре пыльников и микроспор
Роль генотипа. Способность к андрогенезу в культуре пыльников и микроспор существенно различается у разных видов растений. У большинства двудольных растений получить гаплоиды с помощью этого метода проще, чем у однодольных. В свою очередь, среди двудольных наиболее высокую андрогенетическую способность имеют представители семейств пасленовых и крестоцветных. В пределах вида имеются генотипы, значительно различающиеся по этому показателю.
Показано на многих видах растений, что различные этапы андрогенеза (индукция эмбриогенеза и регенерация растений) находятся под контролем разных генов. Отдельным генетически детерминированным признаком является также выход зеленых растений-регенерантов. Этот признак имеет особое значение для злаковых культур, у которых преобладают альбиносные растения-регенеранты.
Как правило, гибриды превосходят по андрогенетической способности родительские формы (гетерозис), однако могут и уступать им. Характер наследования показателей пыльцевого эмбриогенеза различается в зависимости от вида растений и набора изучаемых генотипов. Во многих случаях наблюдали наследование по доминантному или сверхдоминантному типу в сторону высокого выхода эмбриоидов. В других случаях наследование этих признаков было аддитивным.
В исследованиях на пшенице было показано, что индукция пыльцевого эмбриогенеза находится под контролем генов, расположенных на хромосомах 7А и 1В, регенерацию растений из эмбриоидов контролируют хромосомы 3А и 5В, а выход зеленых растений-регенерантов зависит от генов хромосомы 2D (B. Barnabas и др., 2001).
Сравнение транскриптома с помощью микрочипа Barleyl Gene Chip линий ячменя, близких по происхождению, но c разной андрогенетической способностью, позволило идентифицировать у них различия в экспрессии 213 генов. Гены, связанные со способностью микроспор к делению и образованию эмбриоидов, относились к генам, вовлеченным в процессы изменения структуры и функции мембран, а также эффективного использования энергии. Образование зеленых регенерантов было связано с экспрессией генов ответа растения на стресс, регуляции транскрипции и трансляции, деградации специфичных пыльцевых протеинов, альбиносных растений - генов, вовлеченных в процесс развития пластид (M. Munos-Amatriain и др., 2009).
По мнению H. Uhrig и F. Salamini (1987), андрогенетическая способность у сортов и внутривидовых гибридов культурного картофеля Solanum tuberosum L. контролируется немногими или, вероятно, одним доминантным геном. Аналогичные результаты получены в экспериментах с диплоидными видами картофеля. По данным H. Veronneau и других (1992), полученным на диплоидном диком виде картофеля S. chacoense, формирование эмбриоидов в культуре пыльников находится под контролем двух генов, еще два гена контролируют процесс регенерации растений из эмбриоидов. T. E. Taylor, R. E. Veilleux (1992) установили, что процесс регенерации эмбриоидов в культуре пыльников диплоидного примитивного культурного вида картофеля S. phureja контролируется одним доминантным геном, но при этом способность к андрогенезу in vitro находится под контролем аддитивного эффекта других генов.
Помимо ядерных генов, на эффективность андрогенеза могут оказывать влияние и цитоплазматические факторы, что было установлено в опытах на аллоплазматических линиях пшеницы (получены методом беккросса путем замещения цитоплазмы одного вида пшеницы на цитоплазму другого вида) (П. Орлов, 2006). Таким образом, андрогенетическая способность генотипа - комплексный признак, в генетическом контроле которого могут участвовать разные гены и генетические системы.
Влияние физиологического состояния и условий выращивания растений-доноров пыльников на эффективность андрогенеза. Условия выращивания растений, их физиологическое состояние могут оказывать весьма существенное влияние на выход гаплоидов или удвоенных гаплоидов в культуре пыльников или микроспор. В связи с этим нередки случаи, когда в разные годы или при выращивании в разных условиях растений одних и тех же генотипов получают сильно различающиеся результаты. Считается, что более высокий выход гаплоидов получают в культуре пыльников от выращиваемых в поле растений по сравнению с тепличными. Однако во многих случаях из-за сильной контаминации культур, полученных от выращиваемых в поле растений, предпочтительным является использование теплиц. Кроме того, при выращивании в теплицах имеется возможность контролировать ряд важных для андрогенеза факторов среды: интенсивность освещения, его качество (спект-
ральный состав света) и продолжительность (фотопериод), температуру выращивания растений, их питание и полив. В любом случае растения-доноры пыльников должны быть в здоровом состоянии, получать полноценный уход.
E. Heberle-Bors и J. Reinert (1981) показали, что при выращивании растений табака Nicotiana tabacum в условиях короткого дня и пониженных температур происходит существенное увеличение концентрации в пыльце эмбриогенных р-зерен и соответственно увеличение частоты эмбриоидов в культуре микроспор. На основании полученных результатов было высказано предположение, что образование р-зерен - тип мужской стерильности, формирующейся под воздействием во время мейоза материнских клеток пыльцы факторов внешней среды, оказывающих на растения феминизирующий эффект (E. Heberle-Bors, 1982). Это предположение было подтверждено позднее в опытах по обработке разных видов растений физиологически активными веществами, сдвигающими пол в женскую сторону.
В лаборатории генетики картофеля Института генетики и цитологии НАН Беларуси было проведено сравнение эффективности воздействия различных феминизирующих агентов (цитокининов - 6-БАП, кинетина; ауксинов - НОУК, 2,4-Д, а также хлорхолинхлорида и алара, обладающих антигиббереллиновой активностью) на процесс андрогенеза в культуре пыльников сортов картофеля. Лучшие результаты были получены при обработке формирующихся бутонов раствором 6-БАП (20-30 мг/л): наблюдали достоверное увеличение в зрелой пыльце сорта доли р-зерен и увеличение интенсивности андрогенеза в 2-3 раза по сравнению с контролем (A. Yermishin и др., 1989).
В качестве эффективного приема повышения андрогенетической способности сортов картофеля было предложено использовать прививку на томаты (А. Ермишин, Е. Воронкова, 1996). Если для интактного растения картофеля характерна большая листовая поверхность (место синтеза гиббереллинов) при относительно слаборазвитой корневой системе (место синтеза цитокининов), то в результате прививки получается растение с мощной корневой системой подвоя (томатов) и небольшой листовой поверхностью привоя. Следовательно, можно предположить, что микроспорогенез у привоя происходит в гормональном режиме, существенно отличающемся от такового интактного растения картофеля (сдвиг полового баланса в женскую сторону из-за изменения соотношения между цитокининами и гиббереллинами). По сравнению с применением опрыскиваний растений фитогормонами, прививка картофеля на томаты имеет ряд преимуществ: исключается эффект неравномерного попадания физиологически активных веществ на разные части соцветия, обеспечивается постоянное воздействие фитогормонов на созревающие микроспоры.
Изучение влияния различных факторов внешней среды на андрогенетическую способность растений позволило идентифицировать для разных видов способы воздействий, позволяющие существенно усилить у них формирование эмбриогенных микроспор и увеличить выход гаплоидов в культуре пыльников и микроспор. Так, высокотемпературный стресс (воздействие в фазу бутонизации на растения в течение двух суток температурой 32 °С) был успешно использован для повышения андрогенетической способности растений рода Brassica, ряда пасленовых культур (S. Zoriniants и др., 2005), в том числе некоторых клонов S. phureja. Однако для сортов картофеля S. tuberosum этот прием оказался неэффективным. После такого стресса наблюдалось полное осыпание формирующихся бутонов. В отличие от высоких температур, воздействие холода (2 суток при температуре 4 °С) оказалось весьма благоприятным. При этом наблюдали не только значительное (до 30 %) увеличение концентрации эмбриогенных р-зерен, но и одновременное увеличение пыльцевой продуктивности растений. Эти факторы, очевидно, оказывали положительное влияние на андрогенетическую способность картофеля и позволили существенно повысить выход каллюса в культуре пыльников (А. Ермишин, Е. Воронкова, 1990).
Существенное влияние на эффективность андрогенеза in vitro у растений имеет характер освещения растений-доноров пыльников. В исследованиях лаборатории генетики картофеля Института генетики и цитологии НАН Беларуси было показано, что при выращивании растений картофеля в условиях теплицы с досветкой лампами ДРИ-2000-6 значительно повышалась андрогенетическая способность практически всех исследованных сортов картофеля (см. таблицу) (А. Ермишин, Е. Воронкова, 1989).
Показатели андрогенетической способности сортов картофеля в зависимости от источника освещения растений-доноров пыльников (естественное освещение и подсветка лампами ДРИ-2000-6)
Сорт |
Частота каллюсообразования (% от числа эксплантированных пыльников) |
Концентрация р-зерен в зрелой пыльце, % |
Пыльцевая продуктивность (масса пыльцы с 10 цветков), мг |
|||
Естественное освещение |
Подсветка лампами ДРИ-2000-6 |
Естественное освещение |
Подсветка лампами ДРИ-2000-6 |
Естественное освещение |
Подсветка лампами ДРИ-2000-6 |
|
Ирис |
16,88 |
66,06* |
0,38 |
1,31* |
3,10 |
17,60 |
Отрада |
0 |
59,58* |
0,94 |
3,40* |
20,45 |
41,70 |
Ласунак |
32,40 |
50,40* |
1,52 |
2,11* |
8,90 |
27,00 |
Ясень |
2,72 |
24,85* |
0,88 |
2,33* |
7,90 |
12,37 |
Зорька |
7,79 |
36,55* |
0,82 |
3,04* |
4,80 |
9,20 |
Орленок |
1,54 |
25,14* |
0,14 |
1,13* |
20,00 |
42,00 |
* Р ≤ 0,01.
Эффект воздействия света, излучаемого этими лампами, по-видимому, связан с преобладанием в его спектре коротковолнового (синего - 430-490 нм) излучения с тремя максимумами при 400, 500 и 600 нм и сильно пониженной интенсивностью излучения в инфракрасной области (700-760 нм). Предполагается, что воздействие коротковолнового света усиливало синтез в растениях цитокининов, что, как отмечалось выше, может приводить к сдвигу пола в женскую сторону и стимулировать образование эмбриогенных микроспор.
Таким образом, экспериментальные воздействия на растения-доноры пыльников на стадии мейоза позволяют существенно усилить образование эмбриогенных микроспор, и можно сделать вывод, что на данной стадии растения чувствительны к внешним воздействиям для изменения программы развития микроспор с гаметофитной на спорофитную.
Оптимальная фаза развития микроспор для индукции андрогенеза. Как отмечалось выше, изменение программы развития с гаметофитной на спорофитную в культуре in vitroвозможно, только когда микроспоры являются компетентными к условиям культивирования, способным вызвать такое изменение программы. Для большинства видов растений это стадия одноядерной микроспоры. Однако в некоторых случаях лучшие результаты были достигнуты при использовании микроспор, находящихся на более ранних (вовремя, или сразу после мейоза) или поздних (двуядерная микроспора) стадиях развития. В каждом конкретном случае оптимальная для индукции андрогенеза стадия развития микроспор определяется экспериментально с помощью цитологических методов. Поскольку делать это для каждого эксплантируемого пыльника невозможно, используют различные внешние критерии для отбора пыльников, большинство микроспор в которых находится на оптимальной для андрогенеза стадии. Например, берут бутоны определенного размера, или колосья, когда они только появляются из-под флагового листа.
Эффект стрессовых воздействий на растения, эксплантаты или микроспоры до начала культивирования пыльников или микроспор (предобработки). Впервые положительный эффект предобработки растений холодом на выход гаплоидных эмбриоидов в культуре пыльников и микроспор дурмана Datura innoxia обнаружили C. Nitsch и B. Norreel (1973). Как показали дальнейшие исследования, эффект холодового стресса на стадии одноядерной микроспоры выражался в увеличении частоты симметричных митозов микроспор и, как следствие, увеличении выхода эмбриоидов за счет развития микроспор по пути B (см. раздел 7.2). В настоящее время холодовая предобработка эксплантатов для культуры пыльников или микроспор - один из обязательных элементов технологии получения андрогенетических гаплоидов многих видов растений.
Существует несколько вариантов применения этого методического приема. Воздействию холодом подвергают целые растения (редко, из-за трудоемкости), срезанные побеги с бутонами или колосьями на стадии одноядерной микроспоры, помещенные в сосуды с водой или питательной средой (используют чаще всего), бутоны или колосья, помещенные в чашки Петри, эксплантированные пыльники. Обычная температура - около 4 °С (температура в бытовых холодильниках), продолжительность - от 1 до 30 сут.
Помимо холодовой обработки, нашли применение и другие стрессовые воздействия, которые используют в зависимости от вида растений: тепловой стресс (температура 32-38 °С, продолжительность от нескольких часов до нескольких дней), инкубирование пыльников или микроспор на питательной среде, содержащей неметаболизируемые источники углеводов, например, маннитол (углеводное голодание), с пониженным содержанием азота (азотное голодание), на среде с добавлением колхицина. Реже в качестве стрессовых воздействий используют центрифугирование эксплантатов, пониженное атмосферное давление, γ-облучение, инкубирование пыльников на питательной среде с высокой рН, с абсцизовой кислотой, полиэтиленгликолем, этанолом, в присутствии тяжелых металлов и др. Также применяют комбинации стрессовых воздействий, например, тепловой стресс плюс углеводное и азотное голодание.
Наблюдения, сделанные на разных растительных системах, показали, что вне зависимости от типа использованного стрессового воздействия образование эмбриогенных микроспор сопровождается: 1) увеличением размера микроспор; 2) репликацией ДНК или митозом, после которых происходит остановка клеточного цикла; 3) уменьшением объема цитоплазмы; 4) перемещением ядра микроспоры с периферии к центру; 5) фрагментацией вакуоли и образованием цитоплазматических тяжей, идущих от ядра к периферии клетки (формирование звездчатой структуры (см. рис. 7.4)); 6) образованием новой клеточной стенки с внутренней стороны существующей; 7) увеличением компактности хроматина; 8) изменением активности генов.
Механизмы, которые индуцируют описанные изменения микроспор, и каким образом эти изменения определяют перепрограммирование микроспор на спорофитный путь развития, пока неизвестны. Тем не менее имеющейся информации достаточно, чтобы сделать вывод, что стадия одноядерной микроспоры (или ранней двуядерной для некоторых видов) является, наряду со стадией мейоза, второй «чувствительной точкой» микроспорогенеза, когда такое перепрограммирование возможно в ответ на действие внешних воздействий, в качестве которых эффективными оказываются различные стрессы. Все это позволяет рассматривать предобработку растений или генеративных органов перед помещением пыльников или микроспор на питательную среду для культивирования в качестве одного из наиболее важных элементов технологии получения андрогенетических гаплоидов.
Выбор питательной среды и условий культивирования пыльников или микроспор. После того, как запущен механизм перепрограммирования микроспор с гаметофитного на спорофитный путь развития, важно создать благоприятные условия для индукции их делений и образования эмбриоидов или каллюса. Оптимальная температура культивирования пыльников или микроспор обычно не сильно отличается от той, при которой производят культивирование соматических клеток того же вида растений. Считается, что для индукции эмбриогенеза освещение культур не требуется, однако во многих случаях пыльники или микроспоры культивируют на свету (под лампами дневного света), что, как полагают, может стимулировать процесс эмбриогенеза. Сформировавшиеся эмбриоиды для получения растений-регенерантов требуют обязательного освещения, причем необходима оптимизация интенсивности освещения, его продолжительности, подбор источника света с оптимальными спектральными характеристиками света. Подсветка культур необходима также при получении растений-регенерантов из каллюса пыльцевого происхождения.
Культивирование пыльников удобно проводить в пластиковых чашках Петри диаметром 5 см, заклеенных парафильмом (см. рис. 7.2). На поверхность агаризованной среды высаживают 10-15 пыльников. В последнее время предпочтение отдается жидким средам, средам с половинным по сравнению с обычными средами содержанием агара или агарозы (полужидкие среды) или двухслойным средам с нижней агаризованной подложкой и верхним слоем жидкой среды, в которой пыльники свободно плавают. Было показано, что при культивировании пыльников на жидких средах вероятность прямого эмбриогенеза выше, чем на агаризованных. Микроспоры культивируют в тонком слое жидкой питательной среды. При получении культуры микроспор большое значение имеет исходная плотность клеток. Для культуры микроспор пшеницы эффективной является плотность от 5 х 103 до 2 х 105намл среды.
В качестве основной питательной среды для культуры пыльников или микроспор чаще всего используют среду Мурасиге - Скуга или ее модификации, а также среду Нич - Нич (J. P. Nitsch, C. Nitsch, 1969). Для злаковых культур рекомендуют среду № 6, разработанную китайскими исследователями (C. Chu, 1978). Большое распространение получила для злаковых также китайская картофельная среда (она содержит картофельный экстракт, который получают путем продолжительного отваривания молодого картофеля) (C. Chuang и др., 1978). Концентрацию сахарозы, как правило, повышают с обычных 2-3 % до 6-8 %. В последнее время принято вместо сахарозы использовать мальтозу, которая более стабильно поддерживает осмотические свойства питательной среды и дает возможность получать эмбриоиды с более высокой жизнеспособностью. Для культуры микроспор в питательную среду добавляют осмотики (например, для культуры микроспор пшеницы это 0,3М маннитол или 0,25М мальтоза).
Показано, что могут различаться требования разных генотипов к уровню отдельных макро- и микроэлементов, соотношению азотсодержащих солей, добавке в состав среды определенных аминокислот (например, глютамина, аспарагина, серина в качестве источников азота), адсорбирующих агентов (например, активированного угля, поливилипирролидона и др.). Индукционные питательные среды для культуры пыльников или микроспор двудольных растений в качестве обязательных компонентов содержат в разных соотношениях регуляторы роста (ауксины и цитокинины). У злаковых для индукции эмбриогенеза достаточно небольшого количества ауксина (0,1-0,2 мг/л 2,4-Д, более высокие концентрации приводят к образованию преимущественно каллюса).
Для культуры микроспор важным фактором индукционной питательной среды является кокультивирование микроспор с изолированными семяпочками или кондиционирование ее питательной средой, на которой проводили культивирование завязей (OVCM - ovary-conditioned medium). Этот прием способствует выходу эмбриоидов из экзины микроспор, повышает их жизнеспособность, благодаря чему удается получить андрогаплоиды даже у «неотзывчивых» генотипов.
Сформировавшиеся в культуре пыльников эмбриоиды, как правило, развиваются на свету в растения-регенеранты. В отдельных случаях для улучшения условий регенерации или для лучшего укоренения регенерантов их переносят на специальную регенерационную питательную среду. Перенос на регенерационную среду необходим и для получения растений-регенерантов из каллюса. Регенерационная среда содержит, в отличие от индукционной, обычное для культуры соматических клеток количество сахарозы (2-3 %), а также регуляторы роста, необходимые для индукции морфогенеза из каллюса. Эмбриоиды, сформировавшиеся в культуре микроспор, для регенерации растений переносят на полужидкую среду с пониженным содержанием сахаров и без регуляторов роста. Сформировавшиеся растения-регенеранты после процедуры адаптации (см. гл. 5) высаживают в грунт.
7.4. Определение плоидности растений-регенератов. Получение удвоенных гаплоидов, оценка их гомозиготности
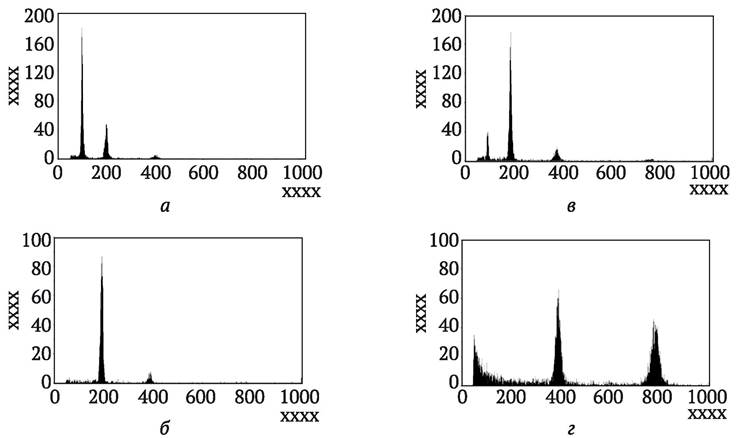
Полученные растения-регенеранты могут быть гаплоидами, удвоенными гаплоидами (в результате спонтанного удвоения хромосом, это оптимальный вариант) или иметь иную плоидность (анеуплоиды и полиплоиды). Наиболее простой метод отделить гаплоиды от диплоидов и полиплоидов по внешнему виду: сеянцы гаплоидов в большинстве случаев явно уступают в мощности и скорости развития. Гаплоиды являются стерильными и не завязывают семян при самоопылении. Еще один простой метод определения плоидности основан на подсчете числа хлоропластов в замыкающих клетках устьиц листа (N. Frandsen, 1968). Так, у моноплоидов картофеля (n = x = 12) это число равно 5-6, у дигаплоидов (2n = 2x = 24) - 10-13, у тетраплоидных сортов (2n = 4x = 48) - от 22 до 30. Точно определить плоидность позволяет подсчет числа хромосом в метафазных пластинках молодых корешков растений- регенерантов, или в мейотических материнских клетках пыльцы на стадии метафазы - анафазы II или пыльцевого митоза. Ускорить этот процесс можно путем анализа ядер клеток растений-регенерантов с помощью проточной цитометрии (англ. flow cytometry) (рис. 7.7).
Рис. 7.7. Определение плоидности растений-регенерантов рукколы Eruca sativa (L.) Mill. с помощью метода проточной цитометрии: а - гаплоид; б - диплоид; в - миксоплоид; 2 - тетраплоид (положение крайнего слева пика определяет плоидность образца, остальные пики связаны с ядрами на стадии G1, или G1 и G2 или эндоредуплицированными ядрами) (по J. Murovec, B. Bohanec, 2012)
В случае если растения-регенеранты - гаплоиды, у них проводят удвоение хромосом. Наиболее распространенная методика основана на применении
колхицина. Колхицин - алкалоид, выделенный из растений безвременника (Colchicum autumnale L.), обладающий свойством ингибировать полимеризацию микротрубочек, формирующихся при клеточном делении, что приводит к нерасхождению хромосом и в результате к увеличению плоидности клеток. Для удвоения хромосом у высаженных в грунт растений-регенерантов, например, ячменя, молодые стеблевые меристемы обрабатывают раствором, содержащим 0,05-0,1 % колхицина и 2 % диметилсульфоксида. Для этого на меристемы прикладывают вату, смоченную таким раствором, или наносят ланолиновую мазь на его основе. Обработки повторяют несколько раз в течение вегетации. Колосья, образовавшиеся из обработанных меристем, содержат диплоидные сегменты, на которых формируются фертильные цветки. Поскольку колхицин - сильно токсичное соединение, вместо него применяют другие вещества с аналогичным действием, например, кофеин, гербициды оризалин, амипрофосметил, трифлуралин, пронамид.
Для удвоения хромосом используют также закись азота. Гаплоидные растения на стадии формирования цветочных примордий помещают на 2 сут в специальную камеру, в которую закачивают закись азота и повышают давление до 600 кПа. Еще один метод удвоения хромосом основан на получении каллюсных культур гаплоидных растений с последующей индукцией из каллюса адвентивных побегов. Как известно, эндомитоз (удвоение хромосом без клеточного деления) - обычное явление для культуры клеток растений (см. разделы 3.3, 6.1). Этот подход показал высокую эффективность для картофеля, лука.
Диплоидные растения-регенеранты могут получиться не только благодаря спонтанному удвоению хромосом гаплоидных клеток, но и в результате регенерации из каллюса, происходящего из соматических клеток генеративных органов. Такие растения не будут удвоенными гаплоидами, а будут иметь генотип материнского растения. Вероятность их появления в целом невелика, особенно в случае развития микроспор с образованием эмбриоидов. Тем не менее иногда требуется подтверждение гомозиготности полученных предположительно удвоенных гаплоидов. Для этого, во-первых, необходимо убедиться, что генотип анализируемых растений отличается от генотипа растений-доноров пыльников (чаще всего это гибриды), во-вторых, их половое потомство имеет высокую однородность, характерную для удвоенных гаплоидов. Для этих целей используют морфологические, биохимические (изоферментные формы) и молекулярно-генетические (AFLP, RAPD, SSR и др.) маркерные признаки.
7.5. Проблема альбинизма растений-регенераторов, полученных в культуре пыльников и микроспор злаковых растений
Одним из основных факторов, лимитирующих получение и использование в практических целях андрогенетических удвоенных гаплоидов, является формирование альбиносных растений-регенерантов (рис. 7.8). Частота таких растений у многих генотипов злаковых культур достигает 100 %. В то же время появление альбиносных растений-регенерантов у двудольных - редкое явление.
Рис. 7.8. Образование альбиносных (а) и зеленых (б) растений-регенерантов в культуре пыльников тритикале

Биохимические и молекулярно-генетические исследования дифференциации пластид в процессе пыльцевого эмбриогенеза полученных в культуре пыльников и микроспор альбиносных растений показали различные нарушения, связанные с размером пластид, организацией у них тилакоидов и гран, аккумуляцией крахмала. Альбиносы теряют рибосомы пластид, у них наблюдаются серьезные нарушения процессов транскрипции и трансляции генов. Были обнаружены большие делеции в пластидном геноме, в том числе делеции районов расположения ключевых генов фотосинтеза, а также перестройки в пластидном геноме. Впрочем, описаны и альбиносные растения с интактными геномами пластид. Было показано, что у таких растений снижен уровень транскрипции пластидных генов фотосинтетического аппарата и рибосомальных РНК, в то время как остальные гены функционировали без изменений.
Следует заметить, что при естественном развитии микроспор по гаметофитному пути пластиды генеративной клетки у большинства покрытосеменных растений полностью исчезают до или вскоре после оплодотворения, а пластиды вегетативной клетки дифференцируются в амилопласты, их ДНК деградируют. Эти процессы лежат в основе наследования хлоропластного генома по материнской линии. Естественно предположить, что аналогичные механизмы деградации пластид действуют и при развитии микроспор по спорофитному пути. Однако у большинства двудольных растений, а также у отдельных генотипов однодольных после перепрограммирования части эмбриогенных микроспор на спорофитный путь происходит развитие нормальных пластид и формируются нормальные зеленые растения-регенеранты.
Было установлено, что образование зеленых растений-регенерантов в культуре пыльников и микроспор находится под генетическим контролем ядерных генов. В частности, у ячменя определены локусы количественных признаков (англ.: QTL - quantitative trait loci) на хромосомах 3Н и 5Н, на которые приходится до 65 % вариации образования зеленых растений-регенерантов (M. Munos-Amatriain и др., 2008).
Соотношение зеленых и альбиносных растений-регенерантов зависит также от применяемых предобработок и условий культивирования пыльников или микроспор. Как показано выше, различные стрессовые воздействия на стадии одноядерной микроспоры могут существенно повысить андрогенетическую способность генотипа за счет увеличения частоты эмбриогенных микроспор. Такие воздействия могут способствовать также увеличению среди растений-регенерантов доли зеленых растений. Однако неправильно подобранные предобработки (по виду, интенсивности и продолжительности стрессового воздействия) могут приводить к увеличению частоты образования альбиносных регенерантов. Согласно одной из теорий, под воздействием стресса в генеративных клетках могут образовываться антибиотикоподобные соединения или протеины, инактивирующие пластидные рибосомы, что, в свою очередь, может приводить к образованию альбиносных регенерантов.
Увеличить частоту появления зеленых растений-регенерантов в культуре пыльников или микроспор можно и путем оптимизации питательных сред. Известно, что снижение уровня азота в питательной среде, особенно его аммонийной формы, замена сахарозы на мальтозу и некоторые другие модификации способствуют увеличению выхода зеленых регенерантов. Эффективной в этом отношении оказалась так называемая китайская картофельная среда, которая содержит, помимо регуляторов роста и сахарозы, лишь шесть основных солей в пониженных концентрациях, тиамин и 10 % картофельный экстракт (C. Chuang и др., 1978).
Несмотря на определенные достижения, полностью решить проблему появления альбиносных растений-регенерантов в культуре пыльников и микроспор злаковых культур пока не удалось. До настоящего времени наиболее эффективным подходом получения гаплоидов у злаковых культур является отбор и использование генотипов, способных с относительно высокой частотой давать преимущественно зеленые растения-регенеранты.
7.6. Получение гаплоидов с помощью культуры неоплодотворенных семяпочек и завязей
Получение растений-регенерантов путем индукции эмбриогенеза или каллюсообразования в культуре неоплодотворенных семяпочек и завязей - альтернативный метод получения гаплоидов или удвоенных гаплоидов у видов растений, для которых метод экспериментального андрогенеза или другие методы получения гаплоидов оказались неэффективными. К таким растениям относятся, прежде всего, сахарная свекла и лук (рис. 7.9). Гиногенетические (т. е. происходящие от женских половых клеток) гаплоиды были получены также у огурцов, кабачков, подсолнечника, пшеницы, ячменя, шелковицы, герберы и других видов.
Рис. 7.9. Получение гаплоидов лука Allium cepa L. путем индукции гиногенеза in vitro: а - культура in vitro неоплодотворенных семяпочек; б - появление гаплоидного эмбриоида через 60-180 дней культивирования семяпочек; в - гаплоидный проросток (по J. Murovec, B. Bohanec, 2012)

В качестве эксплантатов для культивирования in vitro обычно используют молодые неоплодотворенные семяпочки или завязи. Так, для сахарной свеклы оптимальный вариант предполагает использование в качестве эксплантатов семяпочек из закрытых бутонов со средней части соцветия, которые содержат недифференцированные 8-ядерные зародышевые мешки. После помещения на питательную среду развитие женского гаметофита, в отличие от мужского, продолжается и в результате образуется зрелый зародышевый мешок. Он содержит несколько гаплоидных клеток, которые теоретически могут дать начало гаплоидному эмбриоиду или каллюсу: яйцеклетку, синергиды, антиподы. Однако по спорофитному пути, как правило, развивается при определенных условиях культивирования яйцеклетка.
Эффективность получения гиногенетических гаплоидов зависит от вида растений, генотипа внутри вида, условий выращивания растений, физиологического состояния эксплантата (положения соцветий, бутонов в соцветии), условий культивирования (прежде всего, от типа и концентрации углеводов и регуляторов роста в питательной среде). Как и в случае с андрогенезом in vitro, положительное влияние на выход гаплоидов оказывает холодовая предобработка эксплантатов.
У сахарной свеклы выход гиногенетических гаплоидов составляет в среднем от долей процента до 10-12 %, а у лука - до 20 % (от отдельных растений - до 50 %). Во многих случаях происходит спонтанное удвоение хромосом как на индукционных средах, так и в процессе культивирования полученных гаплоидов in vitro. Для экспериментального удвоения хромосом у гаплоидов применяют методы, описанные выше (см. раздел 7.4).
7.7. Получение гаплоидов с помощью гаплопродюсеров
Развитие гаплоидов из женских половых клеток можно индуцировать in situ путем опыления пыльцой другого вида, того же вида или облученной пыльцой. Возможны два основных механизма возникновения гаплоидов в результате таких опылений: 1) после оплодотворения яйцеклетки образуется гибридный зародыш, в котором на ранних стадиях эмбриогенеза происходит элиминация хромосом одного из родителей (чаще всего опылителя); 2) оплодотворение яйцеклетки не происходит, но стимулируется к дифференцировке.
В качестве опылителей, как правило, используют генотипы, способные обеспечить относительно высокий выход гаплоидов, - гаплопродюсеры. Их создают с помощью специальной селекции или выделяют в природных популяциях (рис. 7.10).
Рис. 7.10. Ботанические виды растений, используемые в качестве гаплопродюсеров: а - дикий ячмень Hordeum bulbosum L.; б - просо декоративное Pennisetum glaucum L., в - сорго Sorghum bicolor Moench; 2 - теосинте Zea mays ssp. Mexicana Mill.; д - примитивный культурный вид картофеля Solanum phureja Juz. & Bukasov (клон IvP35)

Получение гаплоидов путем опыления пыльцой другого вида. Классический пример - так называемый «бульбозный» метод, который широко распространен при получении гаплоидов ячменя (K. Kasha, K. Kao, 1970). Материнские растения культурного ячменя Hordeum vulgare L. (2n = 2x = 14) опыляют пыльцой дикого ячменя H. bulbosum L. (2n = 2x = 14). В результате элиминации хромосом дикого вида образуется гаплоидный зародыш, эндосперм получается недоразвитым. Поэтому зародыш через 12-14 дней после опыления вводят в культуру in vitro на среде B-5 Гамборга с 20 г/л сахарозы без регуляторов роста. Полученные гаплоидные растения-регенеранты требуют удвоения хромосом.
Для получения гаплоидов пшеницы ее опыляют пыльцой кукурузы (M. Zenkteller, W. Nitzsche, 1984). После оплодотворения хромосомы кукурузы элиминируют, в результате формируется гаплоидный зародыш материнского растения. Пыльца кукурузы обладает совместимостью со многими видами злаковых культур. Благодаря этому опыление кукурузой нашло применение для получения гаплоидов таких видов, как тритикале, рожь, овес, ячмень и др. Однако имеются проблемы, связанные с нарушением развития эндосперма. Помимо обязательного использования в таких случаях культуры зародышей in vitro (embryo rescue), применяют также обработку колосьев ауксинами, например, 2,4-Д. Это делают, чтобы продлить развитие зародыша на растении, благодаря чему повышается эффективность его введения в культуру. Аналогичным способом можно получать гаплоиды пшеницы, используя в качестве опылителя сорго Sorghum bicolor, теосинте Zea mays ssp Mexicana, декоративное просо Pennisetum glaucum, дикий ячмень H. bulbosum.
Культурный картофель Solanum tuberosum L. (2n = 4x = 48) является автотетраплоидом. Гаплоиды, полученные на основе тетраплоидного картофеля, имеют двойной набор гомологичных хромосом (2n = 2х = 24), поэтому в генетике картофеля их принято называть дигаплоидами. Соответственно гаплоиды, полученные на основе диплоидов картофеля (2n= 2х = 24), называют моноплоидами или моногаплоидами (2n = x= 12).
У картофеля дигаплоид впервые удалось выделить в 1939 г. Е. В. Ивановской (1939) в потомстве от опыления тетраплоидного сорта «Аврора» пыльцой S. tybinii. По современной классификации этот вид не является самостоятельным, а относится к примитивному культурному виду S. phureja (2n = 2x = 24). Хотя в литературе описаны случаи использования для индукции дигаплоидов S. tuberosum и других диплоидных видов картофеля, использование для этой цели S. phureja оказалось наиболее эффективным. Возможность массового получения дигаплоидов картофеля с помощью S. phureja была продемонстрирована R. W. Hougas и сотрудниками в 1958 г.
В основе гаплопродуцирущей способности S. phureja лежит явление псевдогамии (партеногенеза), при котором оба ядра пыльцевых зерен диплоидного вида (n = x = 12) сливаются с центральным ядром зародышевого мешка S. tuberosum (2n = 4x = 48), что ведет к формированию гексаплоидного эндосперма. Ядро яйцеклетки (n = 2x = 24) остается неоплодотворенным, но стимулируется к дифференцировке. Такая структура, сочетающая зародышевый мешок с гексаплоидным эндоспермом и диплоидный зародыш, является вполне жизнеспособной и может развиться в нормальное семя.
Существенный прорыв в создании высокопродуктивных клонов-гапло- продюсеров S. phureja произошел, когда были созданы образцы, сочетающие высокую гаплопродуцирующую способность с гомозиготным состоянием гена (BdBd), контролирующего распределение антоциановой окраски к основанию многих органов растения, в частности семядолей зародыша (J. Hermsen, J. Verdenius, 1973). В связи с тем, что аллель Bd редко встречается в европейских сортах картофеля, их скрещивание с такими гаплопродюсерами дает в потомстве несколько аллельных вариантов. Один из этих вариантов, а именно, отсутствие окраски у основания семядолей, так называемого эмбрионального пятна, свидетельствует о партеногенетическом происхождении семян и дает основание отбирать их в качестве дигаплоидов. Семена с эмбриональным пятном являются настоящими гибридами. Эмбриональное пятно легко идентифицируется при рассмотрении семян под бинокулярной лупой. С введением в практику подобных суперопылителей S. phureja (наиболее известные - голландские формы IvP35, IvP48, IvP101) появилась возможность с минимальными трудозатратами проводить первичный скрининг на наличие дигаплоидов на уровне семян, и затем по отсутствию антоциановой окраски у основания стебля - на уровне сеянцев. Далее проводится анализ значительно сократившейся популяции отобранных сеянцев с применением цитологического анализа. С помощью суперопылителей S. phureja можно получать не только дигаплоиды, но и моноплоиды картофеля.
Получение гаплоидов путем опыления пыльцой гаплопродюсера того же вида, что и материнское растение. Американский селекционер Е. Н. Coe в 1959 г. выделил инбредную линию кукурузы Stok 6, которая в скрещиваниях с другими линиями кукурузы в качестве опылителя была способна давать в потомстве до 2,3 % гаплоидных растений. Позднее на основе этой линии в результате специальной селекции были получены более эффективные гаплопродюсеры, способные давать около 10 % гаплоидов (K. Sarkar и др., 1994; F. Roberи др., 2005). Как и в случае с S. phureja, семена с гаплоидными зародышами при использовании таких гаплопродюсеров являются жизнеспособными. Поэтому применение культуры зародышей не требуется. Более того, современные гаплопродюсеры кукурузы имеют доминантные маркеры анто- циановой окраски зерновок и стебля, что позволяет проводить отбор гаплоидов на уровне семян и среди сеянцев. Созданы также линии-гаплопродюсеры кукурузы, которые используют в качестве материнских форм в целях получения гаплоидов от растений, пыльцой которых их опыляют (J. Kermicle, 1969).
Получение гаплоидов путем опыления облученной пыльцой. Если подобрать дозу облучения пыльцы, можно добиться того, что она сохраняет способность к прорастанию, но не может оплодотворить яйцеклетку. При этом она может индуцировать развитие яйцеклетки по спорофитному пути. Во многих случаях при использовании облученной пыльцы для получения гаплоидов необходимо применение культуры зародышей. Однако у некоторых видов (киви, лука, мандарина, табака) развивались выполненные семена с гаплоидным зародышем. С помощью этого метода получены гаплоиды арбуза, кабачков, дыни, огурца, подсолнечника, сливы, черешни, яблони, груши, ежевики, розы, гвоздики и других видов. Гаплоиды клементина Cytrus Clementina (гибрид между мандарином и апельсином) были получены при использовании для опыления пыльцы триплоидных растений.
7.8. Гаметоклональная изменчивость и ее использование в селекции
Термином «гаметоклональная изменчивость» обозначают изменчивость растений, полученных с помощью различных методов из генеративных клеток (гамет). Основным источником гаметоклональной изменчивости является генетическое разнообразие гамет, которое возникает в процессе рекомбинации генов во время мейоза. Эта изменчивость максимальная в случае получения гаплоидов или удвоенных гаплоидов высокогетерозиготных гибридов и практически незначительная, если получают гаплоиды или удвоенные гаплоиды относительно гомозиготных форм растений, например, сортов самоопыляемых культур, инбредных линий перекрестников. Частота рекомбинации между отдельными локусами может различаться в зависимости от способа получения гаплоидов, их происхождения (от мужских или женских половых клеток).
Поскольку процедура получения гаплоидов во многих случаях связана с культивированием клеток in vitro, то это также может стать источником гаметоклональной изменчивости, которая по своей природе не отличается от сомаклональной изменчивости (см. гл. 6). Эта изменчивость может быть усилена с помощью мутагенов. Следует ожидать, что степень ее выраженности у гаплоидных клеток будет выше, чем у диплоидных и полиплоидных.
В процессе получения гаплоидов могут образовываться диплоидные растения (удвоенные гаплоиды), а также растения более высокого уровня плоидности. Могут встречаться и анеуплоидные формы. Для восстановления фертильности гаплоидов у них проводят удвоение хромосом с помощью методов и веществ (например, колхицина), которые могут вызывать мутации. Изменчивость может возникать как при спонтанном, так и экспериментальном удвоении хромосом у гаплоидов.
Для селекции представляют интерес все названные источники гаметоклональной изменчивости. В случае, когда получают удвоенные гаплоиды относительно гомозиготных форм растений, основную роль играет изменчивость, вызванная культивированием клеток, и изменчивость, возникающая при удвоении хромосом у гаплоидов. При получении удвоенных гаплоидов гибридов селекционную ценность представляет, в основном, изменчивость, обусловленная рекомбинацией генов в процессе мейоза при образовании гамет.
7.9. Изменчивость растений-регенераторов, полученных из нередуцированных гамет
В литературе описано достаточно много примеров выделения перспективных генотипов, превосходящих исходные формы, благодаря отбору среди растений-регенерантов гаметного происхождения, полученных в культуре генеративных органов или изолированных микроспор. Этот метод, как отмечалось выше, в основном используют для улучшения сортов самоопылителей. В Институте генетики и цитологии НАН Беларуси впервые было предложено использовать гаметоклональную вариацию, связанную с формированием нередуцированных гамет, для улучшения сортов картофеля - высоко гетерозиготной полиплоидной культуры, размножаемой вегетативно (Е. Воронкова, А. Ермишин, 2003).
Было установлено, что при культивировании пыльников сортов картофеля наряду с диплоидными растениями-регенерантами в большом количестве образуются тетраплоидные регенеранты, которые могут представлять интерес для селекции. Возможны два основных источника их происхождения: из нормальных диплоидных микроспор путем спонтанного удвоения числа хромосом или из нередуцированных микроспор. Последние образуются с определенной частотой у разных генотипов картофеля в результате наличия у них определенных мейотических мутаций. С точки зрения селекции наибольший интерес представляют мейотические мутации, приводящие к образованию нередуцированных гамет FDR-типа (FDR-first division restitution: реституция (восстановление плоидности) по типу первого деления мейоза) или SDR (second division restitution: реституция (восстановление плоидности) по типу второго деления мейоза). При образовании таких гамет рекомбинация генов не сопровождается существенным снижением уровня гетерозиготности.
FDR связывают с появлением слившихся fs) веретен деления в метафазе II. В результате формируются только два полюса с двумя группами хромосом (см. рис. 7.6). Мейоз заканчивается формированием только одной клеточной стенки (эквационного деления). Образуется диада 2n-микроспор. В основе SDR лежит эндомитоз интерфазных ядер между первым и вторым делением мейоза и преждевременное образование клеточной пластинки редукционного деления. В этом случае метафаза, анафаза и телофаза второго деления отсутствуют. Этот тип SDR связан с мутациями рс-1 и рс-2 (premature cytokinesis - преждевременный цитокинез). Доминирующий механизм образования 2п-яйцеклеток - утрата второго деления мейоза. Он контролируется мутациями os или fc.
Основное различие между FDR и SDR заключается в том, что при FDR две хроматиды каждой хромосомы попадают в разные 2п-споры, что ведет к сохранению родительской гетерозиготности в сформировавшихся гаметах. При SDR хроматиды каждой хромосомы попадают в одну из 2п-спор, что приводит к увеличению гомозиготности. Подсчитано, что средний процент родительской гетерозиготности, представленной в FDR- и SDR-гаметах для четырех типов хромосом с известными сайтами центромер и кроссоверов, составляет 80,2 и 39,6 % соответственно.
Тетраплоидные растения-регенеранты, полученные в культуре пыльников из нормальных микроспор в результате митотического удвоения хромосом (удвоенные дигаплоиды), существенно уступают по продуктивности исходному сорту из-за снижения уровня гетерозиготности. У них наблюдается инбредная депрессия в силу перехода в гомозиготное состояние неблагоприятных аллелей. У тетраплоидных растений-регенерантов, полученных в культуре пыльников из нередуцированных гамет, снижение уровня гетерозиготности не столь значительное, как у удвоенных дигаплоидов. Поэтому инбредная депрессия у них незначительная и по продуктивности они могут не уступать исходному сорту или превосходить его. В то же время вследствие мейотической рекомбинации генов количество аллелей, детерминирующих отдельные селекционные признаки, у них может быть выше или ниже, чем у исходного сорта. Благодаря этому степень выраженности соответствующих признаков у них будет различной, что является весьма важным с точки зрения селекционного улучшения сортов картофеля.
Среди сортов картофеля довольно часто встречаются те, что способны образовывать нередуцированные гаметы. В частности, при изучении коллекции из 32 сортов было выявлено шесть (около 19 %), формирующих более 10 % 2п-пыльцы, семь сортов (22 %) имели от 2 до 10 % диад и триад в пуле микроспор. Основным механизмом формирования диад у сортов картофеля с высоким и средним уровнем проявления признака являлась мутация fs, т. е. большая часть изученных сортов картофеля формировала 2п-гаметы FDR-типа. Мутация pc являлась причиной формирования диад у одного сорта (нередуцированные гаметы формировались по SDR-типу) (V. Zharich и др., 2008).
Экспериментально было установлено, что наибольший выход растений- регенерантов, которые не уступают по продуктивности исходному сорту, однако превосходят его по другим селекционным показателям, был у сортов, формирующих нередуцированные гаметы FDR- или SDR-типа с частотой 10-45 %. У сортов картофеля, не формирующих 2п-гаметы, или формирующих их с невысокой частотой (менее 10 %), среди полученных у них в культуре пыльников растений-регенерантов преобладают удвоенные дигаплоиды, которые уступают по продуктивности исходному сорту. Сорта картофеля с очень высоким уровнем формирования диад (более 45 %) имеют пониженную андрогенетическую способность. У таких форм образуется пыльца с низкой жизнеспособностью с большим содержанием стерильных пыльцевых зерен, нередко наблюдается преждевременное осыпание бутонов (V. Zharich и др., 2008).
Отобранные в результате испытаний лучшие гаметоклоны в основном сохраняли фенотип исходного сорта, не уступая ему по продуктивности и превосходили по устойчивости к фитофторозу, парше обыкновенной, по крахмалистости клубней. Метод имеет для картофеля как вегетативно размножаемой культуры особое значение, поскольку обеспечивает получение молодых клонов с улучшенными характеристиками для ряда выдающихся сортов, хорошо зарекомендовавших себя в производстве.
7.10. Использование технологии получения гаплоидов и удвоенных гаплоидов в селекции
Селекция культур-самоопылителей, как правило, включает получение гибридов между выбранными по определенным показателям родительскими формами и отбор лучших по комплексу признаков растений в последовательных поколениях самоопыления до получения константных (относительно гомозиготных) линий, которые могут рассматриваться в качестве кандидатов в новые сорта. Считается, что у диплоидных видов такие линии могут быть отобраны в лучшем случае в поколении F6.
Использование удвоенных гаплоидов может существенно ускорить этот процесс, поскольку позволяет получать гомозиготные линии в течение одного сезона. Гаплоиды, происходящие от гибридов F1, по сути, представляют собой гаметоклональные варианты, сформировавшиеся в результате рекомбинации генетического материала, полученного от родительских генотипов. Удвоение хромосом у них фиксирует сложившееся сочетание генов, переведя их в гомозиготное состояние. По результатам испытания большого количества удвоенных гаплоидов можно отобрать трансгрессивные формы (т. е. превосходящие родителей по каким-либо количественным признакам, например, продуктивности) с новым сочетанием селекционно-ценных генов, например, генов устойчивости к болезням.
Первоначально были опасения, что селекционная ценность удвоенных гаплоидов может быть ниже, чем линий, созданных традиционной селекцией, так как в первом случае имеет место лишь один цикл рекомбинации, а во втором - несколько. Исследования, посвященные изучению этой проблемы, показали, что у линий, отобранных в поздних поколениях, самоопыление может быть выше, чем у удвоенных гаплоидов частота рекомбинации между близко расположенными локусами. Однако это, как правило, существенно не сказывается на их селекционной ценности. В большинстве случаев лучшие селекционные линии на основе удвоенных гаплоидов не уступали лучшим селекционным линиям, полученным традиционным методом. Представляется интересной история выведения сорта пшеницы Ets Desprez во Франции. Он был отобран среди всего 64 удвоенных гаплоидов, полученных от гибридов F1 (J. de Buyser и др., 1987). Тем не менее некоторые селекционеры рекомендуют использовать для получения удвоенных гаплоидов не гибриды F1, а F2 или F3.
Использование удвоенных гаплоидов представляет значительный интерес и для селекции перекрестно-опыляемых культур, так как позволяет со-
кратить сроки создания родительских гомозиготных линий для получения гетерозисных гибридов. Удвоенные гаплоиды нашли применение и для повышения эффективности рекуррентного отбора: хорошие результаты были получены в схемах, в которых лучшие по комплексу признаков линии (удвоенные гаплоиды) одного цикла селекции использовали в качестве родительских форм для гибридизации в целях получения гибридной популяции следующего цикла, также предполагающего получение удвоенных гаплоидов.
Как известно, в генетическом пуле перекрестников присутствует большое количество неблагоприятных рецессивных аллелей, которые вызывают инбредную депрессию при самоопылении, затрудняя процесс создания гомозиготных линий методами традиционной селекции. Использование технологии получения удвоенных гаплоидов обеспечивает раннюю элиминацию в популяции этих аллелей. В результате геном удвоенных гаплоидов представляет собой более жизнеспособную комбинацию генов, чем у самоопыленных линий. Получение удвоенных гаплоидов у некоторых перекрестно опыляемых культур, например, сахарной свеклы, позволяет решить проблему, связанную с наличием самонесовместимости, которая затрудняет применение самоопыления для получения инбредных линий. Поддержание таких линий возможно с помощью методов клонального размножения in vitro.
Применение гаплоидной технологии представляет также интерес для повышения эффективности использования в селекции мутагенеза. Перевод в гомозиготное состояние мутаций обеспечивает их проявление и фиксацию уже в первом поколении после мутагенной обработки, благодаря чему не требуется самоопыление для выявления рецессивных мутаций. Мутагенная обработка гаплоидных клеток может применяться в сочетании с клеточной селекцией, что позволяет отбирать ценные мутанты с устойчивостью к различным стрессовым воздействиям, токсинам (например, гербицидам).
Для размножения двудомной культуры спаржи Asparagus officinalis женские формы (XX) скрещивают с мужскими (XY). В результате получается популяция, в которой 50 % растений являются женскими и 50 % мужскими. Однако коммерческую ценность представляют лишь мужские растения, имеющие низкое содержание волокон. Получение удвоенных гаплоидов на основе мужских растений (XY) сделало возможным создание супер-опылителей (YY), использование которых позволяет получать популяции спаржи, содержащие 100 % мужских растений (L. Corriolis и др., 1990). Для папайи (Carica papaya), напротив, коммерческую ценность представляют женские растения. Получение удвоенных гаплоидов с помощью культуры пыльников облегчает получение женских чистых линий (F. Rimberia и др., 2006).
Сорт рапса (Brassica napus) Maris Haplona был первым сортом культурных растений, выведенным с помощью технологии удвоенных гаплоидов. Он поступил в производство в 1972 г. (K. Thompson, 1972). С тех пор в совершенствовании этой технологии достигнут значительный прогресс, и она стала рутинной в селекции многих сельскохозяйственных, декоративных и древесных культур. Использование удвоенных гаплоидов в селекции, особенно полученных с помощью культуры пыльников и микроспор, нашло наиболее широкое применение в Китае, где было создано большое количество новых ценных сортов пшеницы, риса, кукурузы, перца и других культур, многие из которых занимают большие площади. Большой резонанс в научном мире вызвало в свое время появление сорта озимой пшеницы Jinghua No. 1, на выведение которого потребовалось лишь шесть лет, т. е. на 4-5 лет меньше по сравнению с традиционной селекцией. За это время были проведены скрещивания, с помощью культуры пыльников на основе гибридов F1 получены удвоенные гаплоиды, проведены их полевые испытания, по результатам которых среди них был отобран сорт. Уже в 1984 г. он занимал 170 тыс. гектаров. К его положительным качествам относится высокая урожайность (4,5-6 т/га), устойчивость к ржавчине, короткостебельность и пригодность к выращиванию в широком диапазоне условий окружающей среды (см. H. Hu, B. Huang, 1987).
По приблизительной оценке, в настоящее время выращивается более трехсот выведенных с помощью гаплоидной технологии сортов и гибридов пшеницы, ячменя, кукурузы, риса, различных капустных (Brassica ssp), баклажанов, перца, спаржи, табака и других культур (M. Germana, 2011).
7.11. Манипуляция с плоидностью исходного материала для повышения эффективности селекции полиплоидов (на примере картоффеля)
Культурный картофель - вегетативно размножаемая культура. В связи с этим его селекция относительно простая: любой генотип, имеющий уникальную комбинацию генов, можно сохранить, размножить и длительное время использовать в хозяйственных и иных целях. Однако селекция картофеля весьма сложная, поскольку картофель S. tuberosum является автотетраплоидом. Новые сорта получают путем гибридизации тетраплоидных гетерозигот, поэтому воспроизвести их половым путем практически невозможно. Для картофеля характерен так называемый тетрасомный тип расщепления, который определяется наличием четырех гомологичных хромосом, случайной их конъюгацией в мейозе и случайным распределением по гаметам. В зависимости от количества доминантных аллелей в определенном локусе хромосомы возможны следующие типы зигот: квадруплекс (АААА), триплекс (АААа), дуплекс (ААаа), симплекс (Аааа) и нуллиплекс (аааа). По сравнению с диплоидами, у которых имеется только два варианта моногибридных скрещиваний, дающих расщепление по фенотипу (Аа х Аа и Аа х аа), у картофеля возможны пять вариантов: ААаа х ААаа (расщепление 35: 1), ААаа х Аааа (11: 1), ААаа х аааа (5: 1), Аааа х Аааа (3: 1), Аааа х аааа (1: 1).
Описанные генетические особенности культурного картофеля в значительной степени затрудняют селекцию этой важной сельскохозяйственной культуры. Для повышения ее эффективности предлагается использовать схему селекции, которая предполагает снижение уровня плоидности селекционного материала до диплоидного, проведение отбора на диплоидном уровне с возвратом на тетраплоидный уровень. В общем эта схема имеет следующий вид:
✵ получение дигаплоидов сортов картофеля;
✵ гибридизация дигаплоидов с дикими диплоидными видами;
✵ селекция на диплоидном уровне: комбинация и концентрация желаемых генов и элиминация нежелательных;
✵ мейотическое удвоение хромосом у отобранных генотипов (гибридизация сортов картофеля с дигаплоидами, формирующими нередуцированные гаметы) (S. Chase, 1963; S. Y. Peloquin и др., 1989).
Первый этап этой схемы, как правило, не вызывает проблем: картофель - одна из немногих сельскохозяйственных культур, для которой разработан действительно эффективный метод снижения уровня плоидности, основанный на использовании индуцированного партеногенеза (см. раздел 7.7). Не является, в принципе, сложным и восстановление тетраплоидного уровня. Для многих генотипов картофеля характерно формирование нередуцированных гамет, что позволяет использовать в селекции мейотическое удвоение хромосом (см. раздел 7.9).
В тех случаях, когда применение мейотического удвоения хромосом невозможно (из-за стерильности пыльцы и/или недостаточно высокого уровня 2п-гамет), для перевода селекционного материала на тетраплоидный уровень может быть использовано митотическое удвоение хромосом. Хотя жизнеспособность и продуктивность полученных тетраплоидов понижена, они могут иметь двойную дозу, по сравнению с исходными дигаплоидами, доминантных аллелей ценных для селекции генов. Благодаря этому они представляют интерес для использования в скрещиваниях в качестве мультиплексных линий (см. ниже) в целях передачи потомству определенных генов, например, устойчивости к болезням и вредителям.
Дополнительные возможности комбинации ценных генов дает использование в селекции соматической гибридизации дигаплоидов картофеля путем слияния протопластов (более подробно в гл. 8). У полученных с помощью этой технологии тетраплоидных гибридов представлены ценные гены обоих родителей. При этом возможно сохранение высокой гетерозиготности соматических гибридов при условии слияния протопластов неродственных родительских дигаплоидов. Современная селекция растений предполагает широкое использование генофонда диких сородичей для расширения аллельного разнообразия культурных форм, интрогрессии в селекционный материал ценных генов устойчивости к болезням, вредителям, неблагоприятным факторам среды. Описано около 220 диких видов картофеля. Большинство из них являются диплоидами (около 200). За небольшим исключением, эти виды практически не скрещиваются с культурным картофелем. Для гибридизации с ними необходимо использовать специальные приемы. Наиболее перспективным является снижение уровня плоидности культурного картофеля до диплоидного (получение дигаплоидов картофеля). Дигаплоиды S. tuberosum (2n = 2x = 24) относительно легко скрещиваются с большинством диплоидных диких видов, формируя жизнеспособные семена и фертильное потомство. Используя селекцию на диплоидном уровне, можно наиболее эффективным способом осуществить интрогрессию ценных генов в селекционный материал.
Гибридизация между дигаплоидами S. tuberosum и дикими диплоидными видами картофеля не является обязательным этапом селекции на диплоидном уровне. Скрещивания с дикими видами позволяют существенно повысить мужскую фертильность дигаплоидов. В принципе возможно проведение селекции на диплоидном уровне с использованием исходного материала, полученного на основе исключительно культурного картофеля. С учетом более простого наследования признаков на диплоидном уровне по сравнению с тетраплоидным, основная задача такой селекции заключается в получении генотипов с новой комбинацией признаков, например, с комплексной устойчивостью к болезням и вредителям.
В современной селекции картофеля при отборе принимается во внимание около 50 показателей. Целый ряд таких признаков, как устойчивость к X-, Y-, S- и M-вирусам, нематодам, раку картофеля и др., контролируется главными генами. Получить нерасщепляющееся потомство по ним на тетраплоидном уровне очень сложно. Так, если мы скрещиваем симплекс по гену устойчивости к нематоде H1 с неустойчивой формой (нуллиплекс), то в потомстве будет только половина устойчивых форм. В случае использования в таких скрещиваниях дуплекса доля устойчивых форм будет 5/6 (расщепление 5:1). И только в потомстве триплекса расщепления по устойчивости к патогену не будет (см. раздел 7.7). В связи с этим в современной селекции картофеля весьма перспективным считается создание родительских линий с комплексом генов устойчивости к болезням и вредителям, у которых названные гены представлены несколькими доминантными аллелями (мультиплексов).
Однако создание мультиплексных линий на тетраплоидном уровне с учетом названных выше генетических особенностей культурного картофеля - весьма сложный процесс. На диплоидном же уровне отбор соответствующих гомозигот вполне реален. Более того, при скрещивании дигаплоидов, формирующих нередуцированные гаметы, с тетраплоидными формами картофеля имеется возможность получать нерасщепляющееся по показателям устойчивости к болезням и вредителям потомство даже в случае использования диплоидных гетерозигот (см. раздел 7.7).
Поскольку FDR 2n-пыльца дигаплоидов более гетерозиготна и более гомогенна, чем n-пыльца тетраплоидных сортов, это позволяет получать более продуктивные, экологически стабильные и гомогенные гибридные популяции. По данным R. Ortiz и др. (1991), варианса ошибки в 4х х 2х семьях (полученных при опылении тетраплоидных клонов пыльцой диплоидных клонов, формирующих нередуцированные гаметы) была в три раза меньше, чем в соответствующих 4х х 4х семьях (полученных путем гибридизации тетраплоидных клонов). Следовательно, для достижения определенного селекционного результата размер гибридной популяции в первом случае может быть в три раза меньше, чем во втором, что позволяет сэкономить значительные материальные и трудовые ресурсы.
Таким образом, селекция картофеля с использованием отбора на диплоидном уровне и манипуляций с плоидностью имеет следующие преимущества по сравнению с традиционной селекцией:
✵ благодаря более простому дисомному наследованию требуется меньший объем популяции для выделения сложных рекомбинантов, отвечающих запросам селекции. Аккумуляция желаемых и элиминация нежелательных генов происходит на диплоидном уровне быстрее, чем на тетраплоидном;
✵ существует возможность широкого использования генетического разнообразия диких диплоидных видов, интрогрессии селекционно-ценных генов диких видов в селекционный материал, расширения аллельного разнообразия S. tuberosum, достижения более высокого уровня гетерозиготности селекционного материала;
✵ благодаря мейотическому удвоению хромосом можно получать более выровненные, продуктивные, экологически стабильные тетраплоидные гибридные популяции, не расщепляющиеся по комплексу признаков, отселектированных на диплоидном уровне, что позволяет существенно повысить вероятность отбора в них селекционно-ценных генотипов.
В ходе развития рассматриваемого направления селекции картофеля его широкие возможности подтверждены на практике. Получены гибриды, превосходящие по продуктивности сорта картофеля, выведенные традиционными методами. Продемонстрирована возможность отбирать их в небольших по объему популяциях. Установлено, что такие формы имеют повышенную экологическую стабильность, устойчивость к стрессам. Успешно осуществлен перенос генов, контролирующих селекционно-ценные признаки, от диких видов культурным сортам, показана возможность быстрого получения форм с комбинацией нужных признаков.
Заключение
Гаплоидами у покрытосеменных растений называют особи, имеющие гаметический набор хромосом по отношению к уровню плоидности родительской особи. Частота их появления в природе крайне незначительна (0,001-0,01 %) и не может удовлетворить потребности генетики и селекции. Разработка методов культуры клеток растений in vitro дала возможность получать в эксперименте гаплоиды и удвоенные гаплоиды с частотой, намного превышающей естественный уровень.
Для редукции числа хромосом у растений применяют два подхода, в основе которых лежит фундаментальное свойство тотипотентности половых клеток. Первый из них предполагает получение гаплоидных растений-регенерантов путем андрогенеза в культуре in vitro пыльников или микроспор, второй - путем гиногенеза из неоплодотворенных яйцеклеток или других гаплоидных клеток зародышевого мешка в культуре in vitro, а также из имеющих гаплоидный зародыш семян, полученных при опылении материнского растения пыльцой другого вида или специального гаплоиндуктора. В последнем случае появление гаплоидного зародыша связано с индуцированным чужеродной пыльцой развитием неоплодворенной яйцеклетки (партеногенеза) либо с избирательной элиминацией в зиготе хромосом опылителя или материнского растения.
Под влиянием определенных воздействий в условиях культуры пыльников in vitro некоторые микроспоры способны менять программу развития с гаметофитной на спорофитную и давать в результате начало эмбриоидам или образовывать каллюс. Из эмбриоидов или каллюса можно получить растения-регенеранты - гаплоиды или удвоенные гаплоиды. Это явление получило название андрогенеза. Изменение программы развития с гаметофитной на спорофитную в культуре in vitro происходит только в том случае, когда микроспоры находятся на определенной стадии развития и чувствительны
(компетентны) к различным внешним воздействиям, способным вызвать такое изменение программы. Имеется достаточно много экспериментальных данных, указывающих на связь между андрогенетической способностью генотипа и формированием микроспор определенного типа (эмбриогенных).
Эмбриогенные микроспоры имеют существенные ультраструктурные и функциональные отличия от нормальных микроспор.
Выявлено несколько путей формирования эмбриоидов или каллюса из эмбриогенных микроспор.
А-путь. Микроспора подвергается первому неравному митотическому делению. В дальнейшем происходят симметричные деления только вегетативной клетки, в результате чего образуются гаплоидные эмбриоиды.
Е-путь. Микроспора подвергается первому неравному митотическому делению, в дальнейшем происходят симметричные деления генеративной клетки. В результате образуются гаплоидные эмбриоиды или каллюс.
В-путь. Первое митотическое деление, в отличие от нормального, является симметричным, что приводит к формированию двух одинаковых вегетативных клеток. Их последующее митотическое деление ведет к образованию многоклеточных структур.
С-путь. Микроспора претерпевает нормальное митотическое деление, в результате которого формируются вегетативное и генеративное ядра. Затем ядра сливаются, последующие митотические деления приводят к образованию многоклеточных структур, которые в дальнейшем развиваются посредством эмбриоидогенеза или каллюсогенеза, давая начало преимущественно удвоенным гаплоидам.
Способность к андрогенезу в культуре пыльников и микроспор существенно различается у разных видов растений. У большинства двудольных растений получить гаплоиды с помощью этого метода проще, чем у однодольных. В свою очередь, среди двудольных наиболее высокую андрогенетическую способность имеют представители семейств пасленовых и крестоцветных. В пределах вида имеются генотипы, значительно различающиеся по этому показателю. Показано на многих видах растений, что различные этапы андрогенеза (индукция эмбриогенеза и регенерация растений) находятся под контролем разных генов. Отдельным генетически детерминированным признаком является также выход зеленых растений-регенерантов.
Экспериментальные воздействия на растения-доноры пыльников на стадии мейоза (холодовой или тепловой стресс, обработка физиологически активными веществами) позволяют существенно усилить образование эмбриогенных микроспор. Повысить эффективность андрогенеза можно также с помощью стрессовых воздействий на эксплантаты в фазу одно-двуядерной пыльцы. Нашли применение холодовой или тепловой стресс, инкубирование пыльников или микроспор на питательной среде, содержащей неметаболизируемые источники углеводов, например, маннитол (углеводное голодание), с пониженным содержанием азота (азотное голодание), на среде с добавлением колхицина. Реже в качестве стрессовых воздействий используют центрифугирование эксплантатов, пониженное атмосферное давление, γ-облучение, инкубирование пыльников на питательной среде с высокой рН, с абсцизовой кислотой, полиэтиленгликолем, этанолом, в присутствии тяже-
лых металлов и др. Также применяют комбинации стрессовых воздействий, например, тепловой стресс плюс углеводное и азотное голодание. Таким образом, стадия одноядерной микроспоры (или ранней двуядерной для некоторых видов) является, наряду со стадией мейоза, второй «чувствительной точкой» микроспорогенеза, когда возможно перепрограммирование развития микроспор в ответ на влияние внешних факторов.
Одним из основных факторов, лимитирующих получение и использование в практических целях андрогенетических удвоенных гаплоидов злаковых, является формирование альбиносных растений-регенерантов. Биохимические и молекулярно-генетические исследования дифференциации пластид в процессе пыльцевого эмбриогенеза, полученных в культуре пыльников и микроспор альбиносных растений, показали различные нарушения, связанные с размером пластид, организацией у них тилакоидов и гран, аккумуляцией крахмала. Альбиносы теряют рибосомы пластид, у них наблюдаются серьезные нарушения процессов транскрипции и трансляции генов. Были обнаружены большие делеции в пластидном геноме, в том числе делеции районов расположения ключевых генов фотосинтеза, а также перестройки в пластидном геноме. Было установлено, что образование зеленых растений- регенерантов в культуре пыльников и микроспор находится под генетическим контролем ядерных генов. Соотношение зеленых и альбиносных растений-регенерантов зависит также от применяемых предобработок и условий культивирования пыльников или микроспор, состава питательной среды.
Получение растений-регенерантов путем индукции эмбриогенеза или каллюсообразования в культуре неоплодотворенных семяпочек и завязей - альтернативный метод получения гаплоидов или удвоенных гаплоидов у видов растений, для которых метод экспериментального андрогенеза или другие методы получения гаплоидов оказались неэффективными. К таким растениям относятся, прежде всего, сахарная свекла и лук. Гиногенетические (т. е. происходящие от женских половых клеток) гаплоиды были получены также у огурцов, кабачков, подсолнечника, пшеницы, ячменя, шелковицы, герберы и других видов. В качестве эксплантатов для культивирования in vitro обычно используют молодые неоплодотворенные семяпочки или завязи. После помещения на питательную среду развитие женского гаметофита, в отличие от мужского, продолжается и в результате образуется зрелый зародышевый мешок. По спорофитному пути, как правило, развивается, при определенных условиях культивирования, яйцеклетка. Эффективность получения гиногенетических гаплоидов зависит от вида растений, генотипа внутри вида, условий выращивания растений, физиологического состояния эксплантата (положения соцветий, бутонов в соцветии), условий культивирования (прежде всего, от типа и концентрации углеводов и регуляторов роста в питательной среде). Как и в случае с андрогенезом in vitro,положительное влияние на выход гаплоидов оказывает холодовая предобработка эксплантатов.
Развитие гаплоидов из женских половых клеток можно индуцировать in situ путем опыления пыльцой другого вида, того же вида или облученной пыльцой. Возможны два основных механизма возникновения гаплоидов в результате таких опылений: 1) после оплодотворения яйцеклетки образуется гибридный зародыш, в котором на ранних стадиях эмбриогенеза происходит элиминация хромосом одного из родителей (чаще всего опылителя); 2) оплодотворение яйцеклетки не происходит, но стимулируется к дифференцировке. В качестве опылителей, как правило, используют генотипы, способные обеспечить относительно высокий выход гаплоидов - гаплопродюсеры. Их создают с помощью специальной селекции, или выделяют в природных популяциях.
Для получения гаплоидов ячменя материнские растения Hordeum vulgare (2n = 2x = 14) опыляют пыльцой дикого ячменя H. bulbosum (2n = 2x = 14). В результате элиминации хромосом дикого вида образуется гаплоидный зародыш, эндосперм получается недоразвитым. Поэтому зародыш через 12-14 дней после опыления вводят в культуру in vitro.Полученные гаплоидные растения-регенеранты требуют удвоения хромосом. Для получения гаплоидов пшеницы и некоторых других злаковых культур применяют опыление пыльцой кукурузы, сорго, теосинте, декоративного проса, дикого ячменя H. bulbosum.
Дигаплоиды культурного картофеля (2n = 2x = 24) получают путем использования в качестве гаплопродюсера примитивного культурного вида картофеля S. phureja (2n = 2x = 24). В основе метода лежит явление псевдогамии (партеногенеза), при котором оба ядра пыльцевых зерен диплоидного вида (n = x = 12) сливаются с центральным ядром зародышевого мешка S. tuberosum (2n = 4x = 48), что ведет к формированию гексаплоидного эндосперма. Ядро яйцеклетки (n = 2x = 24) остается неоплодотворенным, но стимулируется к дифференцировке. Такая структура, сочетающая зародышевый мешок с гексаплоидным эндоспермом и диплоидный зародыш, вполне жизнеспособна и может развиться в нормальное семя. В 1959 г. была выделена инбредная линия кукурузы Stok 6, которая в скрещиваниях с другими линиями кукурузы в качестве опылителя была способна давать в потомстве до 2,3 % гаплоидных растений. Позднее на основе этой линии в результате специальной селекции были получены более эффективные гаплопродюсеры, способные давать около 10 % гаплоидов. Как и в случае с S. phureja, семена с гаплоидными зародышами при использовании таких гаплопродюсеров являются жизнеспособными. Поэтому применение культуры зародышей не требуется. Гаплопродюсеры картофеля и кукурузы имеют доминантные маркеры анто- циановой окраски семян и стебля, что позволяет проводить отбор гаплоидов на уровне семян и среди сеянцев.
Термином гаметоклональная изменчивость обозначают изменчивость растений, полученных с помощью различных методов из генеративных клеток (гамет). Основной источник гаметоклональной изменчивости - генетическое разнообразие гамет, которое возникает в процессе рекомбинации генов во время мейоза. Эта изменчивость максимальная в случае получения гаплоидов или удвоенных гаплоидов высокогетерозиготных гибридов и практически незначительная, если получают гаплоиды или удвоенные гаплоиды относительно гомозиготных форм растений, например, сортов самоопыляемых культур, инбредных линий перекрестников. Поскольку процедура получения гаплоидов во многих случаях связана с культивированием клеток in vitro, этот фактор также может стать источником гаметоклональной изменчивости. Она может быть усилена с помощью мутагенов. Изменчивость возникает и при спонтанном, и при экспериментальном удвоении хромосом у гаплоидов.
Для селекции представляют интерес все названные источники гаметоклональной изменчивости. В случае, когда получают удвоенные гаплоиды относительно гомозиготных форм растений, основную роль играет изменчивость, вызванная культивированием клеток, и изменчивость, возникающая при удвоении хромосом у гаплоидов. При получении удвоенных гаплоидов гибридов селекционную ценность представляет в основном изменчивость, обусловленная рекомбинацией генов в процессе мейоза при образовании гамет.
В литературе описано много примеров выделения перспективных генотипов самоопылителей, превосходящих исходные формы, благодаря отбору среди растений-регенерантов гаметного происхождения, полученных в культуре генеративных органов или изолированных микроспор. Для улучшения высоко гетерозиготной полиплоидной культуры картофеля предложено использовать гаметоклональную вариацию, связанную с формированием нередуцированных гамет. Нередуцированные гаметы образуются с определенной частотой у разных генотипов картофеля в результате наличия у них определенных мейотических мутаций. С точки зрения селекции наибольший интерес представляют мейотические мутации, приводящие к образованию нередуцированных гамет FDR- или SDR-типа. У тетраплоидных растений-регенерантов, полученных в культуре пыльников из нередуцированных гамет, снижение уровня гетерозиготности не столь значительное, как у удвоенных дигаплоидов. Поэтому инбредная депрессия у них незначительная и по продуктивности они могут не уступать исходному сорту или превосходить его. В то же время вследствие мейотической рекомбинации генов количество аллелей, детерминирующих отдельные селекционные признаки, у них может быть выше или ниже, чем у исходного сорта.
Использование удвоенных гаплоидов существенно ускоряет селекцию самоопылителей, поскольку позволяет получать гомозиготные линии в течение одного сезона. Гаплоиды, происходящие от гибридов F1, по сути, представляют собой гаметоклональные варианты, сформировавшиеся в результате рекомбинации генетического материала, полученного от родительских генотипов. Удвоение хромосом у них фиксирует сложившееся сочетание генов, переведя их в гомозиготное состояние. По результатам испытания большого количества удвоенных гаплоидов можно отобрать трансгрессивные формы (т. е. превосходящие родителей по каким-либо количественным признакам, например, продуктивности) с новым сочетанием селекционно ценных генов, например, генов устойчивости к болезням.
Использование удвоенных гаплоидов представляет значительный интерес также для селекции перекрестно-опыляемых культур, поскольку сокращает сроки создания родительских гомозиготных линий для получения гетерозисных гибридов. Удвоенные гаплоиды нашли применение и для повышения эффективности рекуррентного отбора: хорошие результаты были получены в схемах, в которых лучшие по комплексу признаков линии (удвоенные гаплоиды) одного цикла селекции использовали в качестве родительских форм для гибридизации в целях получения гибридной популяции следующего цикла, также предполагающего получение удвоенных гаплоидов.
Для повышения эффективности селекции такой полиплоидной культуры, как картофель (2n = 4x = 48), предлагается использовать схему селекции,
которая предполагает снижение уровня плоидности селекционного материала до диплоидного, проведение отбора на диплоидном уровне с последующим возвратом на тетраплоидный уровень с помощью мейотического или митотического удвоения хромосом, соматической гибридизации дигаплоидов путем слияния протопластов.
Контрольные вопросы
1. Какие подходы используют для экспериментального получения гаплоидов растений?
2. Назовите пути формирования эмбриоидов или каллюса из эмбриогенных микроспор в культуре in vitro.
3. Какие стрессовые воздействия используют для повышения андрогенетической способности растений? На каких этапах онтогенеза донорных растений их применение наиболее эффективно?
4. Опишите методику получения гаплоидов пшеницы с помощью культуры микроспор.
5. Назовите методы отбора гаплоидных растений-регенерантов в культуре пыльников или микроспор. Какие методы используют для получения удвоенных гаплоидов, оценки их гомозиготности?
6. С чем связано образование альбиносных растений-регенерантов в культуре пыльников и микроспор злаковых?
7. Что такое гаплопродюсеры? Приведите примеры гаплопродюсеров с описанием механизма их действия.
8. Что такое гаметоклональная изменчивость? Что лежит в основе ее возникновения?
9. Какие особенности имеет изменчивость растений-регенерантов, полученных в культуре пыльников картофеля из нередуцированных гамет?
10. Для каких целей используют удвоенные гаплоиды в селекции самоопылителей, перекрестников?
11. Каким образом манипуляции с плоидностью могут повысить эффективность селекции полиплоидов, например, картофеля?