Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Основы молекулярной биотехнологии
ДНК, РНК и синтез белка
Структура ДНК
Вся информация о строении и функционировании любого живого организма содержится в закодированном виде в его генетическом материале, основу которого составляет дезоксирибонуклеиновая кислота (ДНК). ДНК большинства организмов — это длинная двухцепочечная полимерная молекула. Последовательность мономерных единиц (дезоксирибонуклеотидов) в одной ее цепи соответствует (комплементарна) последовательности дезоксирибонуклеотидов в другой. Принцип комплементарности обеспечивает идентичность новосинтезированных молекул ДНК, образующихся при их удвоении (репликации), исходным молекулам. Индивидуальными генетическими элементами со строго специфичной нуклеотидной последовательностью, кодирующими определенные продукты, являются гены. Одни из них кодируют белки, другие — только молекулы РНК. Информация, содержащаяся в генах, которые кодируют белки (структурных генах), расшифровывается в ходе двух последовательных процессов: синтеза РНК (транскрипции) и синтеза белка (трансляции). Сначала на определенном участке ДНК как на матрице синтезируется матричная РНК (мРНК). Затем в ходе согласованной работы многокомпонентной системы при участии транспортных РНК (тРНК), мРНК, ферментов и различных белковых факторов осуществляется синтез белковой молекулы. Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы однозначно задает ее структуру и функции.
Для получения ценных биотехнологических продуктов используют гены самых разнообразных организмов. Чтобы лучше понять, как работают биотехнологические системы, рассмотрим строение молекулы ДНК и процессы репликации, транскрипции и трансляции.
Первые данные о химических свойствах ДНК появились в 1868 г. К началу 40-х годов XX в. было установлено, что молекула ДНК — это линейный полимер. Его мономерными единицами являются нуклеотиды, состоящие из азотистого основания, пятиуглеродного сахара (пентозы) и фосфатной группы (рис. 3.1, А). Фосфатная группа присоединена к 5'-атому углерода моносахаридного остатка, а органическое основание — к 1'-атому. Основания в ДНК бывают двух типов: пуриновые [аденин (А) и гуанин (G)] и пиримидиновые [цитозин (С) и тимин (Т)] (рис. 3.1, Б). В ДНК моносахарид представлен 2'-дезоксирибозой, содержащей только одну гидроксильную группу (ОН), а в РНК — рибозой, имеющей две гидроксильные группы. Нуклеотиды соединены друг с другом фосфодиэфирными связями, при этом фосфатная группа 5'-углеродного атома одного нуклеотида связана с 3'-ОН-группой дезоксирибозы соседнего нуклеотида (рис. 3.2). На одном конце полинуклеотидной цепи находится 3'-ОН-группа (3'-конец), а на другом — 5'-фосфатная группа (5'-конец).
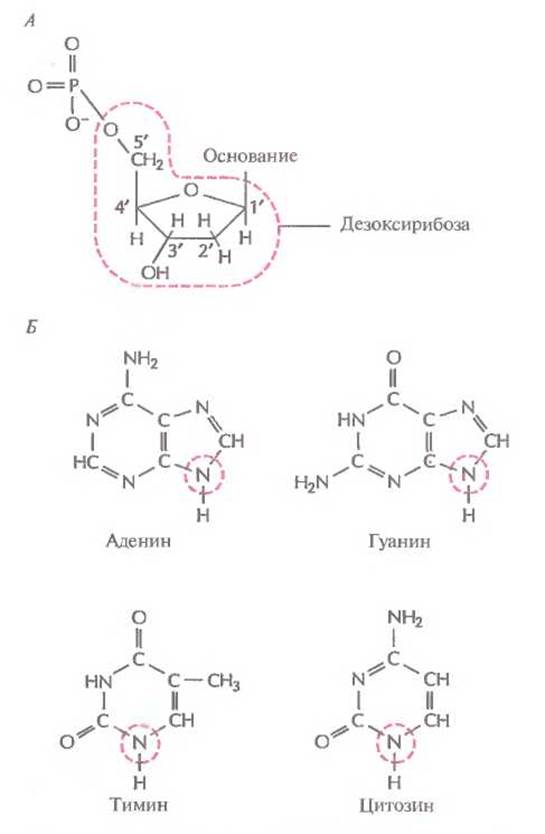
Рис. 3.1. Структурные формулы компонентов ДНК. А. Нуклеотид. Основанием может быть аденин, гуанин, цитозин или тимин. Цветной штриховой линией обведен сахарный остаток (дезоксирибоза): цифрами указаны его углеродные атомы. Б. Основания. Цветной штриховой линией обведен атом азота, по которому к основанию присоединяется дезоксирибоза.
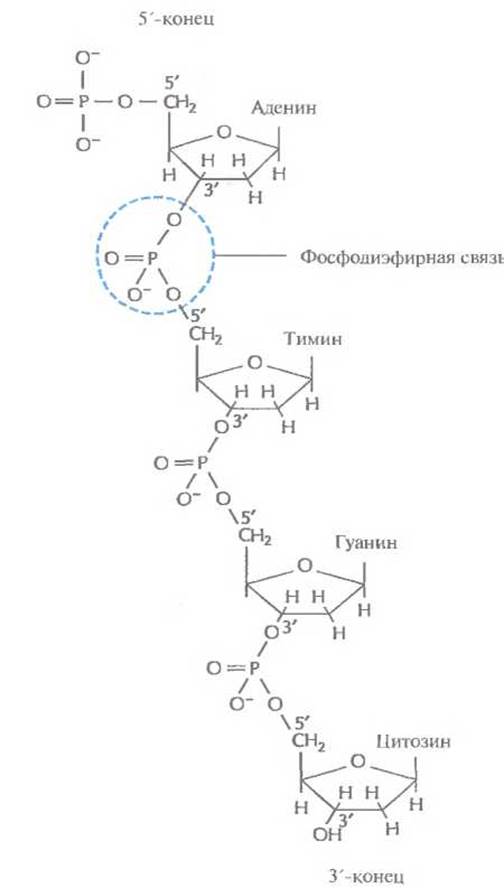
Рис. 3.2. Одна из цепей молекулы ДНК.
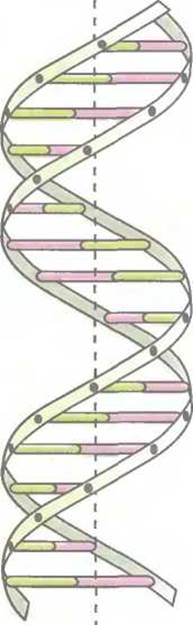
Рис. 3.3. Модель двойной спирали ДНК. Поперечные перекладины — комплементарные пары оснований, «боковины» — сахарофосфатный остов.
В 1953 г. Джеймс Уотсон и Фрэнсис Крик, основываясь на данных рентгеноструктурного анализа кристаллов ДНК, пришли к выводу, что нативная ДНК состоит из двух полимерных цепей, образующих двойную спираль (рис. 3.3). Навитые одна на другую полинуклеотидные цепи удерживаются вместе водородными связями, образующимися между комплементарными основаниями противоположных цепей (рис. 3.4). При этом аденин образует пару только с тимином, а гуанин — с цитозином. Пара оснований А—Т стабилизируется двумя водородными связями, а пара G—С — тремя. Длина двухцепочечной ДНК обычно измеряется числом пар комплементарных нуклеотидов (п.н.). Для молекул ДНК, состоящих из тысяч или миллионов пар нуклеотидов, приняты единицы т.п.н. и м.п.н. соответственно. Например, ДНК хромосомы 1 человека представляет собой одну двойную спираль длиной 263 м.п.н.

Рис. 3.4. Фрагмент двухцепочечной ДНК.
Сахарофосфатный остов молекулы, который состоит из фосфатных групп и дезоксирибозных остатков, соединенных 5'—3'-фосфодиэфирными связями, образует как бы боковины винтовой лестницы, а пары оснований А—Т и G—С — ее ступеньки (рис. 3.4). Цепи молекулы ДНК антипараллельны: одна из них имеет направление 3'→5', другая 5'→3'. В соответствии с принципом комплементарности, если в одной из цепей имеется нуклеотидная последовательность 5'-TAGGCAT-3то в комплементарной цепи в этом месте должна находиться последовательность 3'-ATCCGTA-5'. В этом случае двухцепочечная форма будет выглядеть следующим образом:
![]()
В такой записи 5'-конец верхней цепи всегда располагают слева, а 3'-конец — справа.
Носитель генетической информации должен удовлетворять двум основным требованиям: воспроизводиться (реплицироваться) с высокой точностью и детерминировать (кодировать) синтез белковых молекул. Модель ДНК Уотсона-Крика полностью отвечает этим требованиям. Во-первых, согласно принципу комплементарности, каждая цепь ДНК может служить матрицей для образования новой комплементарной цепи. Следовательно, после одного раунда репликации образуются две дочерние молекулы, каждая из которых имеет такую же нуклеотидную последовательность, как исходная молекула ДНК. Во-вторых, нуклеотидная последовательность структурного гена однозначно задает аминокислотную последовательность кодируемого ею белка.