Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Введение в биологию клетки
Малые молекулы, энергия и биосинтез
Координация катаболизма и биосинтеза
2.5.1. Метаболизм - организуемый и регулируемый процесс
Некоторое представление о том, насколько мудро сконструирована клетка, если рассматривать ее как химическую машину, можно получить из рис. 2-35, где приведена карта, содержащая лишь часть всех метаболических путей. Все эти многочисленные реакции происходят в клетке диаметр которой не превышает 0,1 мм, причем для каждой реакции требуются различные ферменты, которые сами являются продуктам целого ряда реакций переноса информации и синтеза белка. Возьмем любую малую молекулу, например аминокислоту серин; найдется с пол дюжины или более ферментов, способных химически видоизменять ее различными путями. Серин может связываться с AMP (аденилирование на стадии, предшествующей его включению в синтез белков, или расщепляться до глицина, или превращаться в пируват, прежде чем быть окисленным; он может быть ацетилирован с помощью ацетил-СоА или перенесен на жирную кислоту с образованием фосфатидилсерина. Все эти различные пути конкурируют за одну и ту же молекулу серина причем в это же время идет аналогичная конкурентная борьба за тысячи других малых молекул. Можно подумать, что система в целом нуждается в столь тонкой балансировке, что любое незначительное нарушение, например временное изменение рациона, приведет ее к гибели.
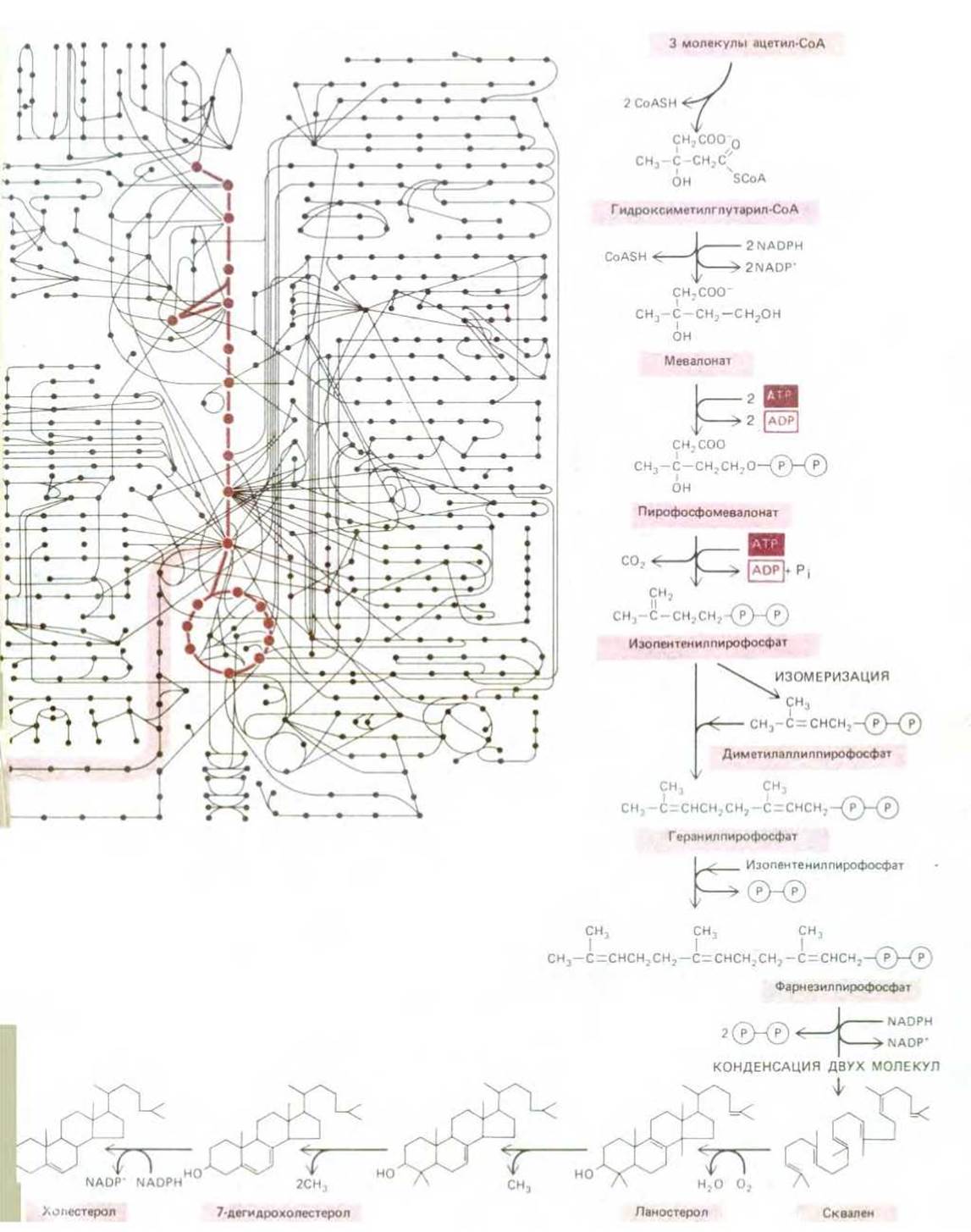
Рис. 2-35. Некоторые из химических реакций, протекающих в клетке. А. Около 500 общих метаболических реакций расходятся в разных направлениях от гликолитического пути и цикла лимонной кислоты (показаны более темной краской). Каждая химическая реакция представлена в виде сплошного кружка. Типичная клетка млекопитающего синтезирует более 10000 белков, большая часть которых - ферменты. На произвольно выбранном участке (выделен более светлой краской) этого сложного переплетения метаболических путей происходит синтез холестерола из ацетил-СоА. Справа и внизу от «лабиринта» этот участок показан более детально в увеличенном масштабе (Б). →
На самом же деле клетке присуща поразительная стабильность. При любых внешних воздействиях реакция клетки направлена на восстановление ее исходного состояния. Она может приспосабливаться и продолжать координированно функционировать во время голода или болезни. Мутации многих типов могут привести к элиминированию отдельных последовательностей реакций, и тем не менее клетка выживает при условии, что удовлетворяются определенные минимальные требования. Это возможно благодаря наличию сложной системы механизмов, регулирующих и координирующих химические реакции в клетках. Некоторые из более высоких уровней контроля рассматриваются в следующих главах. Здесь мы касаемся лишь простейших механизмов, которые регулируют поток малых молекул в различных метаболических путях клетки.
2.5.2. Метаболические пути регулируются изменениями ферментативной активности [20]
Концентрации различных малых молекул в клетке довольно устойчивы, что достигается регуляцией по принципу обратной связи. Регуляторные молекулы такого типа корректируют поток метаболитов по определенному метаболическому пути посредством временного увеличения или уменьшения активности ключевых ферментов. Например, первый фермент в той или иной последовательности реакций обычно ингибируется конечным продуктом этого метаболического пути по принципу отрицательной обратной связи; таким образом, если накапливается слишком много конечного продукта, дальнейшее поступление предшественников в данный метаболический путь автоматически ингибируется (рис. 2-36). В случае ветвления или пересечения метаболических путей, что происходит довольно часто, имеется, как правило, несколько точек, в которых осуществляется контроль различными конечными продуктами. Насколько важны такие процессы регуляции по принципу обратной связи, видно из рис. 2-37, где показана регуляция ферментативной активности в последовательностях реакций, ведущих к синтезу аминокислот.
Регуляция по принципу обратной связи может срабатывать почти мгновенно и является обратимой; кроме того, определенный конечный продукт может ингибировать ферменты, катализирующие его синтез, и при этом быть активатором ферментов другого метаболического пути Молекулярная основа такого типа контроля в клетках хорошо изучена, однако здесь мы данного вопроса касаться не будем, поскольку для этого необходимо иметь представление о структуре белка. Поэтому мы отложим этот вопрос до гл. 3.
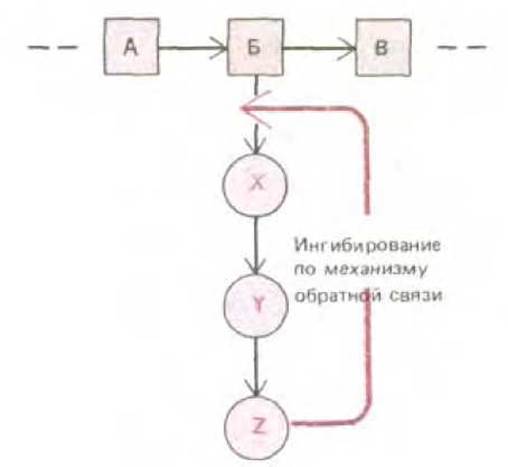
Рис. 2-36. Ингибирование по принципу обратной связи на одном биосинтетическом пути. Конечный продукт Z ингибирует первый фермент, необходимый для синтеза этого продукта, регулируя таким образом собственное содержание в клетке.
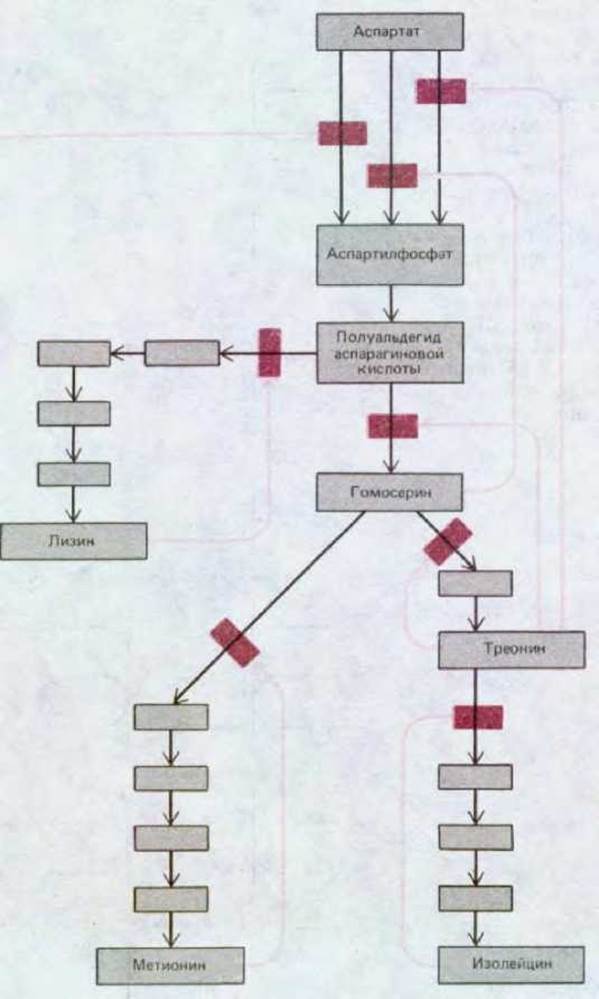
Рис. 2-37. Ингибирование по принципу обратной связи при синтезе аминокислот лизина, метионина, треонина и изолейцина у бактерий. Цветными стрелками показаны участки, в которых происходит ингибирование ферментов продуктами реакций. Отметим что начальную реакцию катализируют три различных фермента (называемые изоферментами), каждый из которых ингибируется своим конечным продуктом.
2.5.3. Катаболические реакции могут обращаться при поглощении энергии [21]
Крупномасштабные изменения, влияющие на метаболизм всей клетки, могут быть достигнуты регуляцией ключевых ферментов. Например, особая схема регуляции по принципу обратной связи позволяет клетке переключаться с расщепления глюкозы на ее биосинтез, или глюконеогенез. Потребность в таком обращении метаболического пути бывает особенно острой как в периоды напряженных тренировок, когда необходимая для мышечного сокращения глюкоза синтезируется в клетках печени, так и во время голодания, при котором глюкоза для выживания организма должна образовываться из жирных кислот и аминокислот. Обычный распад глюкозы до пирувата в процессе гликолиза катализируется несколькими различными последовательно действующими ферментами. Большинство реакций, катализируемых этими ферментами, легко обращается, однако три из них (стадии 1, 3 и 9 на рис. 2-20) фактически необратимы. На самом деле процесс расщепления глюкозы направляется обычно большим отрицательным изменением свободной энергии указанных реакций. Чтобы этот процесс протекал в противоположном направлении и происходило образование глюкозы из пирувата, вокруг каждой из этих реакций должен существовать обходной путь (шунт), причем реакции обходного пути должны идти «вверх» - процесс, происходящий с потреблением энергии (рис. 2-38). Таким образом, если при расщеплении одной молекулы глюкозы до двух молекул пирувата образуются две молекулы АТР, то для обращения реакции в процессе глюконеогенеза необходим гидролиз четырех молекул АТР и двух молекул GTP. Это эквивалентно гидролизу шести молекул АТР на каждую вновь синтезированную молекулу глюкозы.

Рис. 2-38. Сравнение реакций, ведущих к синтезу глюкозы в процессе глюконеогенеза, с реакциями расщепления глюкозы в ходе гликолиза. Реакции расщепления глюкозы (гликолитические реакции) энергетически выгодны (изменение свободной энергии меньше нуля), тогда как реакции синтеза протекают с затратой энергии. Для синтеза глюкозы необходимы различные ферменты обходного пути, шунтирующие реакции 1, 3 и 9 гликолиза. Общее направление протекания реакций между глюкозой и пируватом определяется контрольными механизмами, действующими по принципу обратной связи на указанных трех ключевых стадиях.
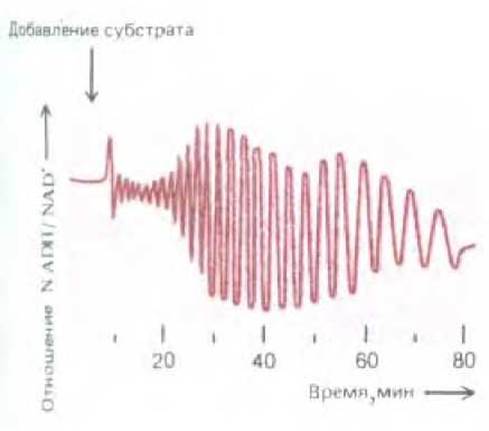
Рис. 2-39. Внезапное добавление глюкозы к экстракту, содержащему ферменты и коферменты гликолиза, может вызвать значительные флуктуации в концентрации определенных промежуточных продуктов, например NADH. Такие метаболические колебания могут быть, в частности, обусловлены регуляцией гликолитического фермента фосфофруктокиназы по принципу положительной обратной связи.
Реакции обходного пути (рис. 2-38) должны жестко контролироваться, так чтобы глюкоза быстро расщеплялась в случае «энергетического голода», а синтезировалась тогда, когда клетка насыщена питательными веществами. Если бы прямые и обратные реакции могли протекать без ограничений, они впустую гоняли бы множество метаболитов туда и обратно через холостые циклы, потребляющие огромные количества АТР и бесполезно выделяющие тепло.
Изящество подобных механизмов контроля можно проиллюстрировать на одном примере. Этап 3 гликолиза представляет собой одну из реакций, которая должна быть шунтирована при образовании глюкозы. Обычно на этом этапе происходит присоединение к фруктозо-6-фосфату второй фосфатной группы из АТР; реакция катализируется ферментом фосфофруктокиназой. Этот фермент активируется AMP, ADP и неорганическим фосфатом, а ингибируется АТР, цитратом и жирными кислотами. Иначе говоря, фермент активируется, когда запасы энергии малы и накапливаются продукты распада АТР; инактивируется же он в том случае, когда имеются обильные запасы энергии (в форме АТР) либо питательных веществ в виде жирных кислот или цитрата (извлекаемого из аминокислот). Ферментом, катализирующим обращенную (обходную) реакцию (образование глюкозы в результате гидролиза фруктозо-1,6-бисфосфата до фруктозо-6-фосфата), является фруктозобисфосфатаза. Активность этого фермента регулируется по принципу обратной связи таким же образом, что и действие фосфофруктокиназы, но с противоположным эффектом, так что фруктозо-бисфосфатаза активна, когда неактивна фосфофрукто киназа.
Отметим, что фосфофруктокиназа активируется ADP, представляющим собой продукт катализируемой данным ферментом реакции (АТР + фруктозо-6-фосфат → ADP + фруктозе-1,6-бисфосфат), и ингибируется АТР - одним из субстратов этой реакции. В результате этот фермент может активировать сам себя, являясь объектом сложного контроля по принципу положительной обратной связи. При определенных условиях такой контроль по принципу обратной связи вызывает необычные колебания активности данного фермента, что приводит к соответствующим колебаниям концентраций различных промежуточных продуктов гликолиза (рис. 2-39). Физиологическое значение таких специфических колебаний неясно. Однако на их примере можно видеть, как благодаря нескольким ферментам может быть создан биологический осциллятор. Такие осцилляции в принципе могут служить своеобразными «внутренними часами», позволяющими клетке «отсчитывать время» и, к примеру, выполнять конкретные функции в определенные периоды.
2.5.4. Ферменты могут переходить в активное или неактивное состояние путем ковалентных модификаций [22]
Рассмотренные выше типы контроля по принципу обратной связи позволяют осуществлять непрерывную регуляцию скоростей протекания метаболических последовательностей в ответ на ежесекундные флуктуации метаболизма. Кроме того, клетки обладают специальными регуляторными механизмами для ситуаций, в которых требуется продолжительное (от нескольких минут до нескольких часов) изменение активности ферментов. Такие механизмы включают в себя обратимые ковалентные модификации ферментов, которые часто (хотя и не всегда) достигаются присоединением фосфатной группы к специфическому (серии, треонин или тирозин) аминокислотному остатку фермента. Фосфат поступает от АТР, а его перенос катализируется ферментами, называемыми протеинкиназами.
В следующей главе мы рассмотрим вопрос о том, каким образом изменение формы фермента при фосфорилировании усиливает или подавляет его активность. Последующее удаление фосфатной группы, сводящее к нулю эффект фосфорилирования, достигается при помощи другого фермента, называемого фосфопротеин-фосфатазой. Ковалентная модификация ферментов - это регуляция в новом измерении, поскольку она делает возможной регуляцию специфических последовательностей реакций такими сигналами (например, гормонами), которые не являются промежуточными продуктами метаболизма.
2.5.5. Реакции компартментализованы как на уровне клеток, так и на уровне всего организма [23]
Не все метаболические реакции клетки протекают в одних и тех же субклеточных компартментах (обособленных субклеточных структурах). Поскольку различные ферменты находятся в разных компартментах клетки, поток химических компонентов направляется не только химическим, но и физическим путем.
Простейшая форма такого пространственного разобщения наблюдается, когда два фермента, катализирующие две последовательные реакции, образуют единый ферментный комплекс, и, следовательно, продукту первой ферментативной реакции не нужно диффундировать через цитоплазму, чтобы встретиться со вторым ферментом. Как только заканчивается первая реакция, сразу же начинается вторая. Некоторые крупные агрегаты ферментов осуществляют всю последовательность реакций, оставаясь в контакте с субстратом. Например, превращение пирувата в ацетил-СоА происходит в три этапа, каждый из которых протекает на одном и том же ферментном комплексе (рис. 2-40), a при синтезе жирной кислоты даже еще более длинная последовательность реакций катализируется единым ферментным ансамблем. Неудивительно, что некоторые из наиболее крупных ферментных комплексов ответственны за синтез макромолекул такого типа, как белки и ДНК.
На следующем уровне пространственного разобщения в клетке происходит концентрирование функционально связанных ферментов в одной и той же мембране или в ограниченных мембранами водим компартментах органелл. Проиллюстрировать это можно на примере метаболизма глюкозы (рис. 2-41). Образовавшийся в результате гликолиза пируват активно захватывается из цитозоля во внутренне пространство митохондрий, где имеются все ферменты и метаболиты цикла лимонной кислоты. Более того, сама внутренняя митохондриальная мембрана содержит все ферменты, катализирующие последовательные реакции окислительного фосфорилирования, включая реакции переноса электронов от NADH к О2 и реакции синтеза АТР. Следовательно, всю митохондрию можно считать небольшим заводом, производящим АТР. Аналогичным образом другие клеточные органеллы, такие, например, как ядро, аппарат Гольджи и лизосомы, можно рассматривать как специализированные компартменты, в которые заключены функционально связанные ферменты для выполнения специальных задач. В определенном смысле живая клетка подобна современному городу со множеством специализированных служб, концентрирующихся в разных районах и связанных друг с другом при помощи обширной сети различных коммуникаций.

Рис. 2-40. Строение пируват-дегидрогеназы - крупного мультиферментного комплекса, в котором промежуточные продукты реакции переходят непосредственно от одного фермента к другому. Этот ферментный комплекс катализирует превращение пирувата в ацетил-СоА.

Рис. 2-41. Пространственное разобщение трех стадий расщепления глюкозы в эукариотической клетке. Гликолиз осуществляется в цитозоле, тогда как реакции цикла лимонной кислоты и окислительного фосфорилирования - только в митохондриях.
В многоклеточных организмах пространственная организация выходит далеко за пределы отдельной клетки. Различные ткани тела обладают разнообразным набором ферментов и по-разному способствуют выживанию организма. Кроме различий в специализированных продуктах, таких, как гормоны или антитела, между разными типами клеток одного и того же организма имеются еще и существенные различия в общих для всех клеток метаболических путях. Хотя фактически во всех клетках имеются ферменты гликолиза, цикла лимонной кислоты, синтеза и распада липидов и метаболизма аминокислот, уровни всех этих процессов по-разному регулируются в различных тканях. Нервные клетки, возможно, наиболее «привередливые» клетки организма, содержат крайне малые запасы гликогена или жирных кислот, целиком «полагаясь» на глюкозу, поставляемую с кровью. Клетки печени снабжают глюкозой клетки активно работающих мышц. Кроме того, они используют молочную кислоту, образованную в мышцах, для синтеза глюкозы (рис. 2-42). Клетки каждого типа обладают специфическими для них особенностями метаболизма и широко сотрудничают как в нормальном состоянии, так и при тренировках, стрессе или голодании.
Заключение
Тысячи и тысячи различных биохимических реакций, одновременно осуществляемых клеткой, тесно скоординированы между собой. Разнообразные механизмы контроля регулируют активность клеточных ферментов при изменении существующих в клетке условий. Наиболее общая форма регуляции - это легко обратимое ингибирование по принципу обратной связи, когда первый фермент метаболического пути ингибируется конечным продуктом этого пути. Более длительная форма регуляции включает в себя химическую модификацию одного фермента под действием другого, что часто происходит в результате фосфорилирования. Комбинации регуляторных механизмов могут вызывать сильные и длительные изменения в метаболизме клетки. Не все клеточные реакции происходят в одних и тех же внутриклеточных компартментах, и пространственное разграничение клетки внутренними мембранами позволяет органеллам осуществлять специализацию своих биохимических функций.
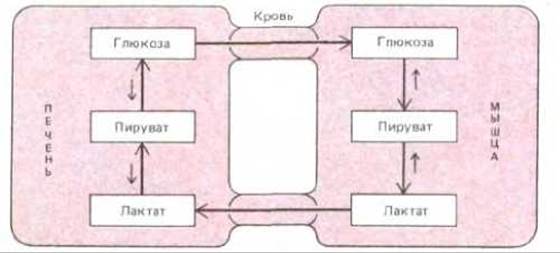
Рис. 2-42. Схематическое изображение метаболического взаимодействия между клетками печени и мышц. Основным «топливом» для клеток активно работающих мышц служит глюкоза, значительная часть которой поставляется клетками печени. Молочная кислота - конечный продукт анаэробного распада глюкозы в мышцах в процессе гликолиза - вновь превращается в глюкозу в ходе глюконеогенеза в печени.
Литература
Основная
Herriott J.; Jacobson G; Marmur J.; Parsom W. Papers in Biochemistry. Reading, MA: Addison-Wesley, 1984.
*Lehninger A.L. Principles of Biochemistry. New York: Worth, 1982.
Stryer L. Biochemistry, 3rd ed. New York: W.H. Freeman, 1988.
Wood W.B.; Wilson J.H.; Benbow R.M.; Hood I.E. Biochemistry: A Problems Approach, 2nd ed. Menlo Park. CA: Benjamin-Cummings, 1981.
Цитируемая
1. Henderson L.J. The Fitness of the Environment. Boston: Beacon, 1927; reprinted 1958. (Классический анализ в доступном изложении.)
2. Ingroham J. L.; Maaloe О.; Neidhardt F. C. Growth of the Bacterial Cell. Sunderland, MA: Sinauer, 1983.
3. Roehrig K. L. Carbohydrate Biochemistry and Metabolism. Westport, CT: AUI Publishing, 1984.
Sharon N. Carbohydrates. Sci. Am., 243(5): 90-116, 1980.
4. Robertson R. N. The Lively Membranes. Cambridge, U. K.; Cambridge University Press, 1983.
5. Saenger W. Principles of Nucleic Acid Structure. New York: Springer, 1984.
6. Hess В.; Markus M. Order and chaos in biochemistry. Trends Biochem. Sci. 12,: 45-48, 1987.
Lehninger A.L. Bioenergetics: The Molecular Basis of Biological Energy Transformations. 2nd ed. Menlo Park, CA: Benjamin-Commings, 1971.
**Schrodinger E. What is Life? Mind and Matter. Cambridge, U. K.: Cambridge University Press, 1969.
7. Dickerson R.E. Molecular Thermodynamics. Menlo Park. Ca.; Benjamin, 1969.
Klotz I. M. Energy Changes in Biochemical Reactions. New York: Academic Press, 1967.
8. Raven P. H.; Evert R. H.; Eihhorn S. E. Biology of Plants, 4th ed. New York: Worth, 1981.
9. Racker E. From Pasteur to Mitchell: a hundred years of bioenergetics. Fed. Proc. 39: 210-215, 1980.
10. Fothergill-Gilmore L.A. The evolution of the glicolitic pathway. Trends Biochem. Sci. 11: 47-51, 1986.
Shulman R.G. NMR Spectroscopy of living cells. Sci. Am. 248(1): 86-93, 1983.
11. Lipmann F. Wanderings of a Biochemist. New York: Wiley, 1971.
12. Schlenk F. The ancestry, birth and adolescence of ATP. Trends Biochem. Sci. 12: 367-368, 1987.
13. McGilvery R. W. Biochemistry: A Functional Approach, 3rd ed. Philadelphia: Saunders, 1983.
Racker E. A. New Look at Mechanisms in Bioenergetics. New York: Academic Press, 1976.
14. Kornberg H.L. Tricarboxylic acid cycles. Bioessays 7: 236-238, 1987.
15. Krebs H. A. The history of the tricarboxylic acid cycle. Perspect. Biol. Med. 14: 154-170, 1970.
Krebs H. A.; Martin A. Reminiscences and Reflections. Oxford, U. K.: Clarendon Press; New York: Oxford University Press, 1981.
16. Pullmann M. E.; at al. Partial resolution of the ensymes catalyzing oxidative phosphorylation. J. Biol. Chem. 235: 3322-3329, 1960.
17. Prigogine I.; Stengers I. Order out Chaos: Man's New Dialogue with Nature. New York: Bantam Books, 1984.
18. EisenbergD.; CrothersD. Physical Chemistry with Applications to the Life Sciences. Menlo Park, CA: Benjamin-Cummings, 1979.
Reich J. G.; Selkov E. E. Energy Metabolism of the Cell - A Theoretical Treatise. New York: Academic Press, 1981.
19. MartinB.R. Metabolic Regulation: A Molecular Approach. Oxford, U.K.; Blackwell Scientific, 1987.
***NewsholmeE.A.; Start C. Regulation in Metabolism. New York: Wiley, 1973.
20. Pardee A. B. Molecular basis of biological regulation: origins of feedback inhibition and allostery. Bioessays 2: 37-40, 1985.
21. HessB. Oscillating reactions. Trends Biochem. Sci. 2: 193-195, 1977.
22. Cohen P. Control of Enzyme Activity. 2nd ed. London: Chapman and Hall, 1983.
В русском переводе имеются следующие издания
*Ленинджер А. Основы биохимии. В 3-х томах. -М.: Мир. 1985.
**Шредингер Э. Что такое жизнь с точки зрения физика. Изд. 2-е. М., Атомиздат, 1972.
***Ньюсхолм Э., Старт К. Регуляция метаболизма. М.: Мир, 1977.