Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Введение в биологию клетки
Макромолекулы: структура, форма и информационные функции
Нуклеиновые кислоты
3.2.1. Гены состоят из ДНК [9]
Еще в те времена, когда человек начал сеять культурные растения и разводить домашних животных, было очевидно, что каждое зернышко или оплодотворенная яйцеклетка должны содержать скрытый план или схему развития организма. Уже в наше время возникла генетика - наука, в основу которой легли представления о генах - невидимых, содержащих информацию элементах, равномерно распределяемых между двумя дочерними клетками при каждом клеточном делении. Чтобы передать дочерним клеткам полный набор генов, перед делением клетка должна сделать копию этих генов. Гены спермия и яйцеклетки передают наследственную информацию от поколения к поколению.
В наследовании биологических признаков используются совокупности атомов, подчиняющихся физическим и химическим законам. Другими словами, гены должны состоять из молекул. Вначале трудно было представить себе природу этих молекул. Что это за молекула, которая могла бы храниться в клетке, направлять процесс развития организма и быть в то же время способной к точной и практически неограниченной репликации?
К концу XIX столетия биологи обнаружили, что хромосомы (которые становятся различимыми в ядре в начале деления) являются носителями наследственной информации. Но данные о том, что веществом, из которого состоят гены, является дезоксирибонуклеиновая кислота (ДНК) хромосом, были получены значительно позже при изучении бактерий. В 1944 г. было установлено, что очищенная ДНК одного бактериального штамма способна передавать наследственные свойства этого штамма другому штамму, несколько отличному от первого. Это открытие оказалось слишком неожиданным и не получило широкого признания до начала 50-х годов, так как считалось, что лишь белки обладают достаточно сложной конформацией, чтобы быть носителями заключенной в генах информации. Сегодня представление о том, что именно ДНК является носителем генетической информации (хранящейся в ее длинных полинуклеотидных цепях), столь прочно вошло в биологическое мышление, что порой трудно осознать, какой огромный пробел в наших знаниях заполнило это представление.
3.2.2. Молекулы ДНК состоят из двух длинных комплементарных цепей, удерживаемых вместе благодаря спариванию оснований [10]
Учитывая простое химическое строение ДНК, легко понять, почему генетики с таким трудом признали в ней носителя наследственности. ДНК - это длинный неразветвленный полимер, состоящий всего из четырех субъединиц-дезоксирибонуклеотидов, азотистые основания которых представлены аденином (А), цитозином (С), гуанином (G) и тимином (Т). Нуклеотиды связаны между собой ковалентными фосфодиэфирными связями, соединяющими 5'-атом углерода одного остатка с 3'-атомом углерода следующего остатка (схема 2-6). Основания четырех типов «нанизаны» на сахарфосфатную цепь наподобие четырех разных типов бусинок, надетых на одну нитку.
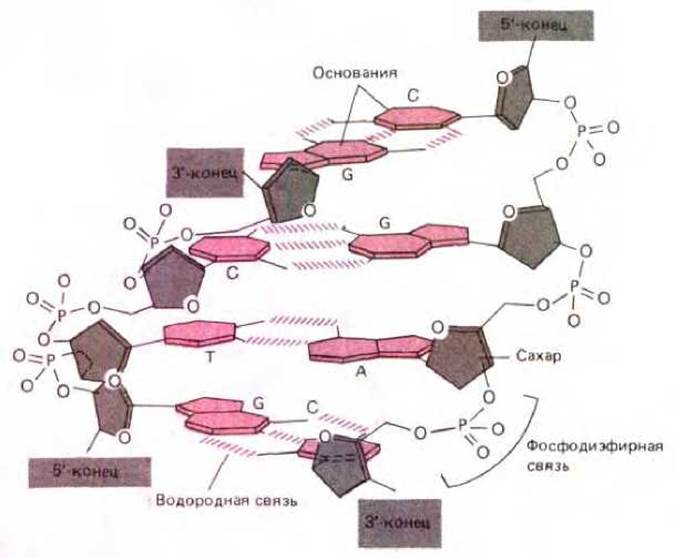
Рис. 3-8. Короткий участок двойной спирали ДНК. Показаны четыре пары комплементарных оснований (цветные) и дезоксирибоза (серая).
Следует обратить внимание, что две цепи ДНК закручены в противоположных направлениях и что каждая пара оснований удерживается вместе либо двумя, либо тремя водородными связями (см. также схему 3-2).
Как же может длинная полинуклеотидная цепь кодировать программу развития целого организма или даже одной клетки? И как эта программа передается от одного поколения клеток к другому? Ответ заключен в пространственной структуре молекулы ДНК.
Полученные в начале 50-х годов данные рентгеноструктурного анализа ДНК указывали на то, что молекула ДНК имеет форму спирали, состоящей из двух цепей. Спиральное строение ДНК не вызвало удивления, поскольку, как мы уже убедились, спирализация - частое явление для полимеров, состоящих из регулярно ориентированных субъединиц. Всех поразил тот факт, что ДНК состоит из двух цепей. В 1953 г. была предложена модель структуры ДНК, удовлетворяющая рентгеноструктурным данным и связывающая воедино структуру и функцию ДНК (рис. 3-8 и схема 3-2).
Согласно этой модели, разработанной Уотсоном и Криком, все основания ДНК расположены внутри двойной спирали, а сахарофосфатный остов - снаружи. Отсюда следует, что основания одной цепи должны быть очень сильно сближены с основаниями другой цепи. Это предположение требовало наличия специфического связывания между большим пуриновым основанием (А или G, каждое из которых имеет двойной гетероцикл) одной цепи и меньшим по размеру пиримидиновым основанием (Т или С с одинарным гетероциклом) другой цепи.
И данные ранних биохимических опытов, и выводы из построенной модели приводили к заключению, что между А и Т и между G и С (так называемые пары оснований Уотсона и Крика) происходит комплементарное спаривание. Биохимические анализы препаратов ДНК, выделенных из разных видов, показали, что, хотя нуклеотидный состав ДНК широко варьирует (например, содержание А у разных видов бактерий колеблется от 13 до 36%), наблюдается общая закономерность: количество G всегда равно количеству С и количество А-количеству Т. Построенная модель показала, что число эффективных водородных связей, которые могут образоваться между G и С или между А и Т будет в этом случае больше, чем при любой другой комбинации. Таким образом, двухспиральная модель ДНК изящно объяснила количественные биохимические результаты.
3.2.3. Структура ДНК дает ключ к пониманию механизмов наследственности [11]
Биологическая информация записана в гене в такой форме, что она может точно копироваться и передаваться клеткам-потомкам. Огромное значение расшифровки структуры ДНК состоит в том, что предложенная Уотсоном и Криком модель позволила сформулировать общие принципы важнейшего процесса передачи генетической информации. Поскольку каждая цепь содержит последовательность нуклеотидов, в точности комплементарную последовательности цепи-партнера, то на деле обе цепи несут одну и ту же генетическую информацию. Если обозначить две цепи А и А', то цепь А служит шаблоном, или матрицей, для образования новой цепи А', а цепь А' может играть ту же роль в образовании новой цепи А. Таким образом, генетическая информация может копироваться при разделении цепей А и А', что позволяет каждой из них служить матрицей для образования нового комплементарного партнера.
Уже сам по себе механизм комплементарного копирования указывает, что наследственная информация ДНК записана в линейной последовательности нуклеотидов. Каждый нуклеотид - А, G, Т или С - можно рассматривать как букву в простом четырехбуквенном алфавите, который используется для написания биологических инструкций в виде линейной «телеграфной ленты». Животные разных видов отличаются друг от друга, потому что молекулы ДНК их клеток имеют различную последовательность нуклеотидов и, следовательно, различное информационное содержание.
Число различных последовательностей ДНК, которые могут быть составлены из n нуклеотидов, равно 4n. Поэтому ДНК даже умеренной длины способна обеспечить колоссальное биологическое разнообразие, а типичная клетка животного содержит ДНК длиной около метра (3-109 нуклеотидов). Записанный в виде линейной последовательности один необычно маленький ген человека занял бы четверть страницы текста (рис. 39), а записанная в таком виде генетическая информация клетки человека представляла собой книгу в 500000 страниц!
Хотя принцип репликации генов прост и элегантен, реальный клеточный аппарат копирования сложен и включает в себя много различных белков. Основная реакция показана на рис. 3-10. Фермент ДНК-полимераза катализирует присоединение дезоксирибонуклеотида к 3'-концу цепи ДНК. Каждый нуклеотид вступает в реакцию в форме дезоксирибонуклеозидтрифосфата; высвобождение из этой активированной формы пирофосфата и его последующий гидролиз обеспечивают энергией реакцию репликации ДНК и делают ее фактически необратимой (см. разд. 5.3).
Репликация ДНК начинается с локального разделения двух комплементарных цепей. Затем каждая цепь используется в качестве матрицы для образования новой молекулы ДНК путем последовательного присоединения дезоксирибонуклеотидов. Выбор каждого следующего нуклеотида происходит на основе его способности образовывать комплементарную пару с очередным нуклеотидом родительской матричной цепи (рис. 3-10). В результате генетическая информация полностью удваивается - в конце концов образуются две полные двойные спирали ДНК, каждая из которых идентична родительской молекуле ДНК по последовательности нуклеотидов. Поскольку две цепи родительской молекулы в конце концов оказываются в разных дочерних молекулах ДНК, механизм репликации называют «полуконсервативным» (рис. 3-11).
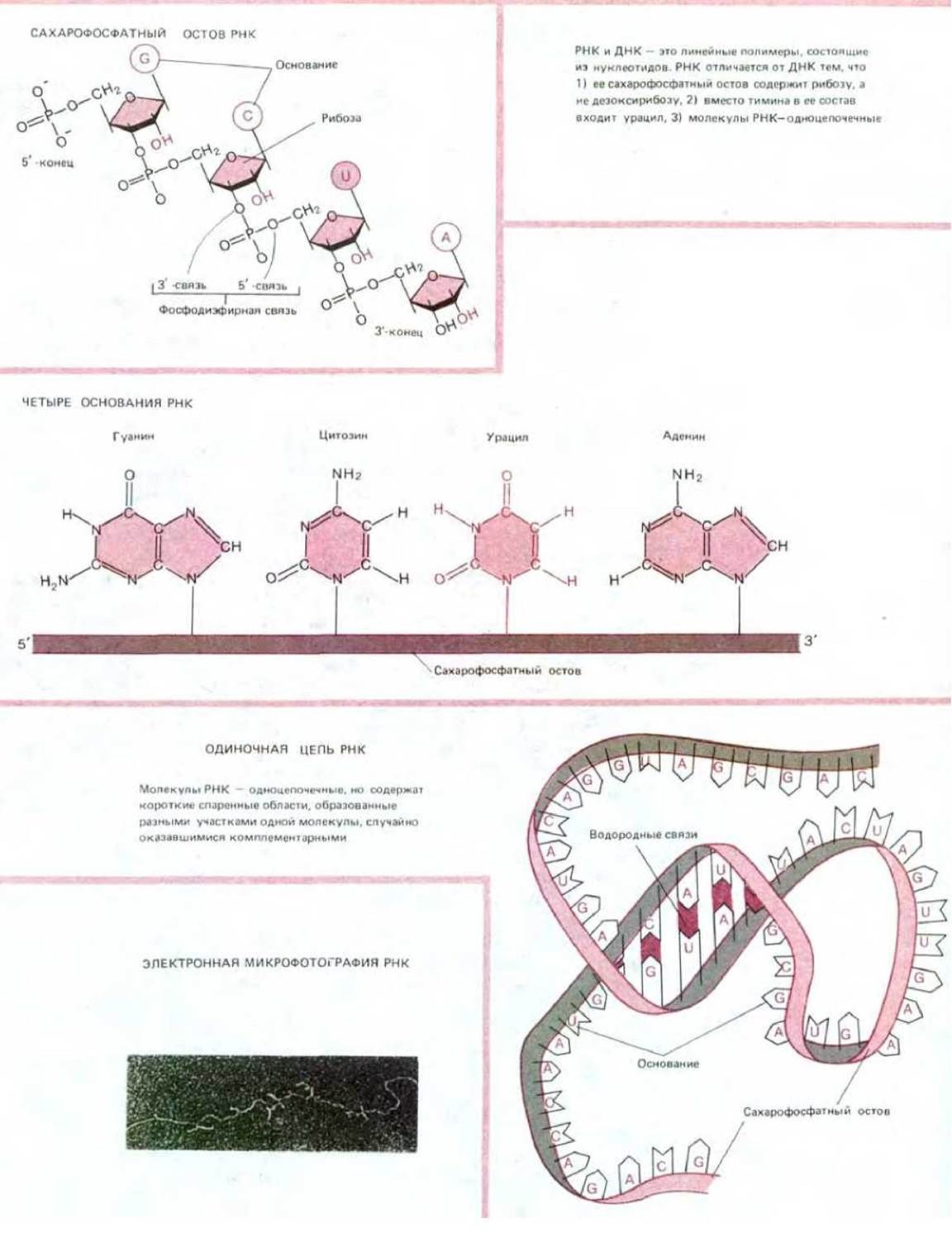
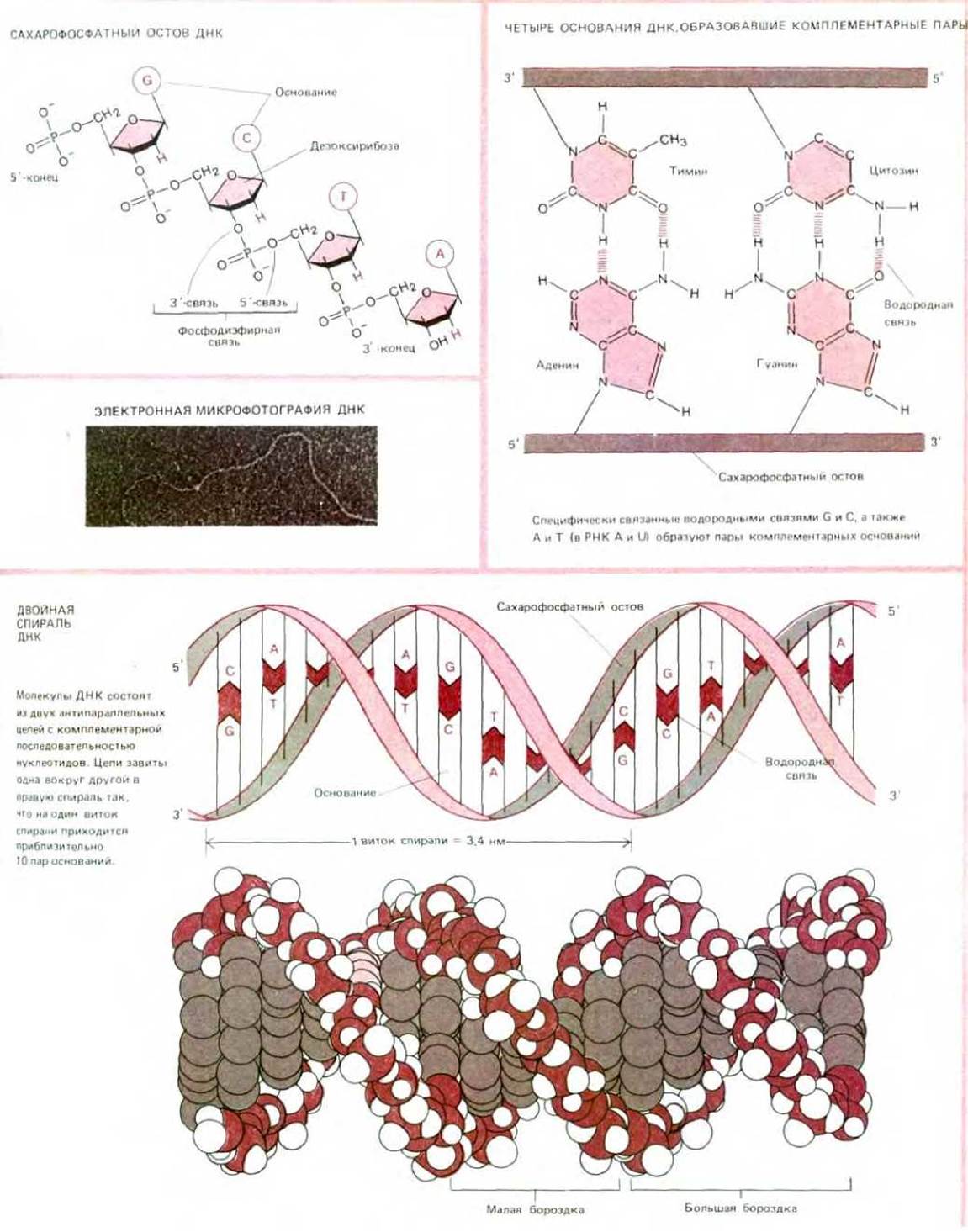

Рис. 3-9. Последовательность ДНК гена ß -глобулина человека (одной из двух субъединиц молекулы гемоглобина, переносящего кислород в крови взрослого человека). Показана только одна из двух цепей ДНК («кодирующая цепь»), вторая цепь имеет комплементарную последовательность. Последовательность следует читать слева направо, строка за строкой, как обычный текст.
3.2.4. Ошибки репликации ДНК приводят к мутациям [12]
Наиболее впечатляющая особенность репликации ДНК - ее высокая точность. Для устранения неправильно расположенных нуклеотидов используется несколько «корректорских» механизмов, в результате их работы последовательность нуклеотидов в молекуле ДНК копируется очень точно (одна ошибка на 109 присоединенных нуклеотидов).
Но иногда, хотя и очень редко, репликативная машина пропускает несколько оснований или вставляет несколько лишних, включает Т вместо С или А вместо G. Каждое такое изменение последовательности ДНК - генетическая ошибка, называемая мутацией. Такие ошибки будут воспроизводиться во всех последующих поколениях клеток, так как «плохие» последовательности ДНК копируются столь же успешно, как и «хорошие». Последствия такой ошибки могут быть существенными, поскольку даже один измененный нуклеотид способен оказать большое влияние на работу клетки, в зависимости от того, где эта мутация произошла.
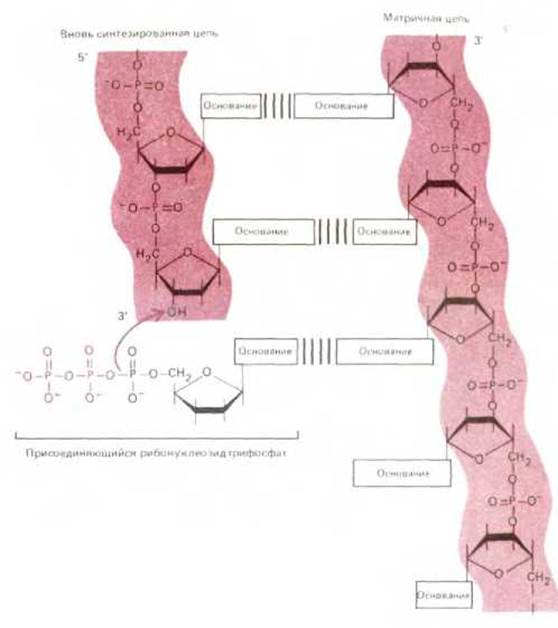
Рис. 3-10. Основная реакция при синтезе новых молекул ДНК -это добавление дезоксирибонуклеотида к 3'-концу растущей цепи. Показано, как спаривание оснований поступающих дезоксирибонуклеотидов и исходной (матричной) цепи ДНК направляет образование дочерней цепи ДНК с комплементарной последовательностью оснований.

Рис. 3-11. Полуконсервативная репликация ДНК. В каждом цикле репликации каждая из двух цепей ДНК используется в качестве матрицы для образования новой комплементарной цепи. Поэтому на протяжении многих клеточных поколений исходные цепи сохраняют свою целостность.
В начале 40-х годов генетики окончательно доказали, что единицы последовательности, называемые генами, определяют структуру индивидуальных белков. Поэтому мутация гена, вызванная изменением последовательности его ДНК, может инактивировать ключевой белок, и клетка тогда погибнет. В результате измененная последовательность ДНК потеряется. Мутация может произойти в несущественном участке и не будет иметь эффекта; такие мутации называют молчащими. Очень редко в результате мутации образуется ген с улучшенными или новыми полезными функциями. В этих случаях несущий мутацию организм будет иметь преимущества и мутантный ген может в конце концов путем естественного отбора заменить исходный ген в большей части популяции.
3.2.5. Последовательность нуклеотидов в гене определяет последовательность аминокислот в белке [13]
Химически ДНК относительно инертна. Содержащаяся в ней информация выражается опосредованно через другие молекулы: ДНК направляет синтез специфических РНК и белковых молекул, которые и определяют химические и физические свойства клеток.
Примерно в то же время, когда биофизики с помощью дифракции рентгеновских лучей исследовали пространственную структуру ДНК, биохимики интенсивно изучали химическое строение белков. Уже было известно, что белки - это цепи аминокислот, последовательно соединенных пептидными связями; но лишь в начале 50-х годов, когда была определена последовательность аминокислот маленького белка инсулина (рис. 3-12), было установлено, что каждый тип белка образует полипептидная цепь со строго определенной последовательностью аминокислот.
Подобно тому как для выяснения молекулярных основ генетики и наследственности решающую роль сыграла расшифровка структуры ДНК, определение последовательности инсулина имело основополагающее значение для выяснения структуры и функций белков. Если инсулин имеет определенную генетически детерминированную последовательность, то, видимо, то же относится и ко всем другим белкам. Более того, можно предположить, что свойства того или иного белка должны зависеть от той конкретной последовательности, в которой расположены в этом белке аминокислоты.
И ДНК, и белки образованы линейной последовательностью мономеров. В результате биохимического анализа белков - продуктов мутантных генов - в конце концов было показано, что последовательность двух этих полимеров колинеарна: последовательность нуклеотидов в участке ДНК, кодирующем белок, соответствует последовательности аминокислот в этом белке. Стало очевидным, что последовательность ДНК содержит закодированную информацию о белковых последовательностях. Центральной проблемой молекулярной биологии стал вопрос о том, как же клетка осуществляет такое биохимически сложное превращение, как перевод последовательности нуклеотидов ДНК в последовательность аминокислот белка.

Рис. 3-12. Последовательность аминокислот инсулина крупного рогатого скота. Инсулин - маленький белок, состоящий из двух полипептидных цепей, каждая из которых обладает уникальной, генетически детерминированной последовательностью аминокислот. Для обозначения аминокислот использованы трехбуквенные символы, приведенные на схеме 2-5. Показаны дисульфидные (-S-S-) связи между остатками цистеина. Изначально белок синтезируется как одна длинная полипептидная цепь (кодируемая одним геном), которая затем разделяется, давая две цепи (см. рис. 3-48).
3.2.6. С последовательностей ДНК снимаются РНК-копии для синтеза белка [14]
При синтезе белка определенные участки ДНК, называемые генами, копируются в виде другого полинуклеотида - рибонуклеиновой кислоты, или РНК, - отличающегося от ДНК как по химическому составу, так и по выполняемой функции. Подобно ДНК, РНК образована линейной последовательностью нуклеотидов, но имеет два небольших химических отличия: 1) вместо дезоксирибозы сахарофосфатный остов содержит сахар рибозу и 2) вместо основания тимина (Т) в РНК содержится близкородственное основание урацил (U) (см. рис. 3-6).
РНК сохраняет все информационное содержание той ДНК, копией которой она является, а также способность к спариванию комплементарных оснований. Синтез молекул РНК называется транскрипцией ДНК; во многих отношениях он аналогичен репликации ДНК. Одна из цепей ДНК служит матрицей, на которой испытывается способность очередных нуклеотидов к комплементарному спариванию. При хорошем соответствии с ДНК-матрицей рибонуклеотид включается в растущую цепь РНК как ковалентно связанная составная часть. Таким способом цепь РНК удлиняется последовательным добавлением одного нуклеотида.
Транскрипция отличается от репликации ДНК рядом особенностей. Во-первых, РНК-продукт не остается комплементарно связанным с ДНК-матрицей. Как только синтез копии РНК завершен, исходная двойная спираль ДНК восстанавливается, а молекула РНК освобождается. Таким образом, молекулы РНК одноцепочечные. Более того, молекулы РНК короче ДНК, так как являются копиями участков ДНК ограниченной длины, достаточной для кодирования одного или нескольких белков (рис. 3-13). РНК-транскрипты, направляющие синтез белковых молекул, называются информационными (матричными) РНК (мРНК); другие РНК-транскрипты используются как транспортные РНК (тРНК) (см. разд. 3.2.10), образуют компоненты рибосом (рибосомные, рРНК, см. разд. 3.2.10) или более мелкие нуклеопротеиновые частицы. Количество молекул РНК, копируемых с определенного участка ДНК, контролируется регуляторними белками, которые связываются со специфическими участками ДНК, закрывая кодирующие последовательности гена (см. разд. 10.2.1). В любой клетке в любой момент времени некоторые гены используются для синтеза РНК в очень больших количествах, тогда как другие гены не транскрибируются совсем. Для некоторых активных генов в каждом клеточном поколении один и тот же участок ДНК может транскрибироваться тысячи раз. Поскольку каждая молекула РНК может транслироваться во многие тысячи копий, то информация, содержащаяся в маленьком участке ДНК, может направлять синтез миллионов копий специфического белка. Например, белок фиброин - основной компонент шелка: один ген фиброина в каждой клетке шелкоотделительной железы производит 104 копий мРНК, на каждой из которых синтезируется 105 молекул фиброина, что за 4 сут дает 109 молекул фиброина на клетку.
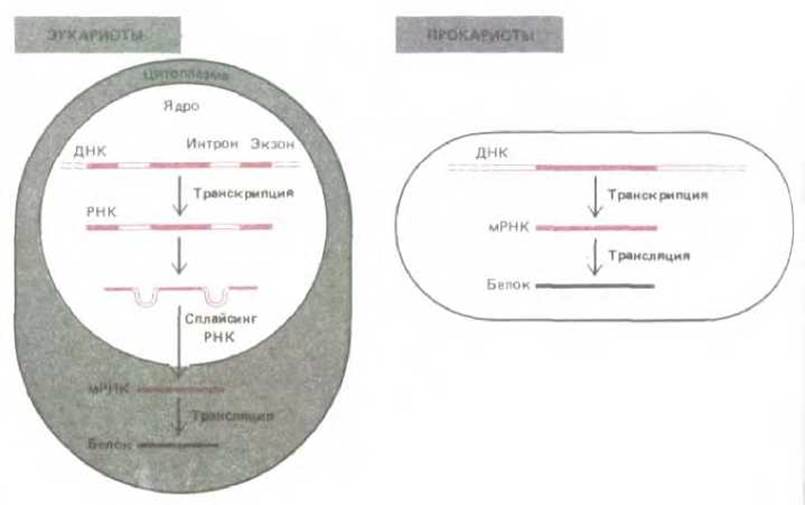
Рис. 3-13. Передача информации от ДНК к белку осуществляется с помощью РНК-посредника, называемого мРНК. У прокариотических клеток этот процесс проще, чем у эукариотических. У эукариот кодирующие участки ДНК-экзоны (выделены цветом) разделены некодирующими участками (интронами). Показано, что для образования мРНК интроны должны удаляться.
3.2.7. Молекулы РНК эукариотических клеток подвергаются сплайсингу, чтобы убрать интронные последовательности [15]
В бактериальных клетках большинство белков кодируется одной непрерывной последовательностью ДНК, которая копируется без изменения с образованием молекулы мРНК. В 1977 г. молекулярные биологи были изумлены, обнаружив, что у большинства эукариотических генов кодирующие последовательности (названные экзонами), чередуются с некодирующими последовательностями (названными нитронами). Для производства белка весь ген, включая и интроны, и экзоны, транскрибируется в очень длинную молекулу РНК (первичный транскрипт). Перед тем как эта молекула РНК покинет ядро, комплекс ферментов, осуществляющих процессинг РНК, удаляет у нее все последовательности интронов, делая молекулу РНК значительно короче. После завершения этой стадии процессинга РНК, которая носит название сплайсинга РНК, молекула РНК выходит в цитоплазму уже как мРНК и направляет синтез определенного белка (см. рис. 3-13).
Этот кажущийся расточительным способ передачи информации развился у эукариот, видимо, потому, что он делает синтез белка значительно более гибким. Например, первичные транскрипты РНК одного и того же гена могут подвергаться сплайсингу разными способами, давая разные мРНК в зависимости от клеточного типа или стадии развития. Это позволяет производить разные белки под контролем одного и того же гена. Более того, поскольку присутствие многочисленных нитронов облегчает генетическую рекомбинацию между экзонами, такой способ устройства гена, видимо, имел огромное значение в ранней эволюционной истории, ускоряя процесс, посредством которого организмы синтезировали новые белки из частей ранее существовавших, вместо того, чтобы вырабатывать целиком новые последовательности.

Рис. 3-14. Три рамки считывания, возможные при синтезе белка. Последовательность нуклеотидов РНК считывается по порядку от 5'- к 3'-концу по три нуклеотида и таким образом переводится в последовательность аминокислот. Поэтому одна и та же последовательность РНК может в принципе в зависимости от «рамки считывания» кодировать три совершенно различные последовательности аминокислот.
3.2.8. Последовательность мРНК «считывается» группами по три нуклеотида и переводится в последовательность аминокислот [16]
Правила перевода последовательности полинуклеотидов в аминокислотную последовательность белков - так называемый генетический код - были расшифрованы в начале 60-х годов. Оказалось, что последовательность нуклеотидов молекулы мРНК - посредника при передаче информации от ДНК к белку - считывается по порядку группами из трех нуклеотидов. Каждый триплет нуклеотидов, или кодон, определяет включение одной аминокислоты, и в принципе каждая молекула мРНК может быть прочитана в любой из трех рамок считывания в зависимости от того, с какого именно нуклеотида молекулы начался процесс декодирования (рис. 3-14). Почти всегда лишь одна из трех рамок считывания дает функциональный белок. Так как, за исключением начала и конца кодирующего участка, информация записана в РНК без знаков препинания, рамка считывания устанавливается при инициации трансляции и сохраняется на протяжении всего процесса.
Поскольку РНК является линейным полимером, состоящим из нуклеотидов четырех типов, то всего имеется 43 = 64 возможных триплета (напомним, что важное значение имеет последовательность нуклеотидов триплета). Учитывая, что в белках находят всего 20 различных аминокислот, можно сделать вывод, что большинство аминокислот должно кодироваться несколькими триплетами; другими словами генетический код вырожден. Генетический код, представленный на рис. 3-15, оказался чрезвычайно консервативным в эволюции: за небольшими исключениями он остается одинаковым у таких разных организмов, как бактерии, растения и человек.
3.2.9. Соответствие между аминокислотами и триплетами нуклеотидов устанавливают молекулы тРНК [17]
Кодоны мРНК узнают соответствующие аминокислоты не прямым путем - не так, как фермент узнает субстрат. При трансляции используются «адапторы» - молекулы, узнающие и аминокислоту, и триплет нуклеотидных оснований. Роль адапторов выполняет набор маленьких (длиной всего около 80 нуклеотидов каждая) молекул РНК, называемых транспортными РНК (или тРНК).

Рис. 3-15. Генетический код. При синтезе белка триплеты нуклеотидов РНК (кодоны) транслируются в соответствующие им аминокислоты. Например, кодоны GUG и GAG направляют в белок соответственно валин и глутаминовую кислоту. Обратите внимание, что кодоны с U или С во втором положении обычно кодируют более гидрофобные аминокислоты (схема 2-5).
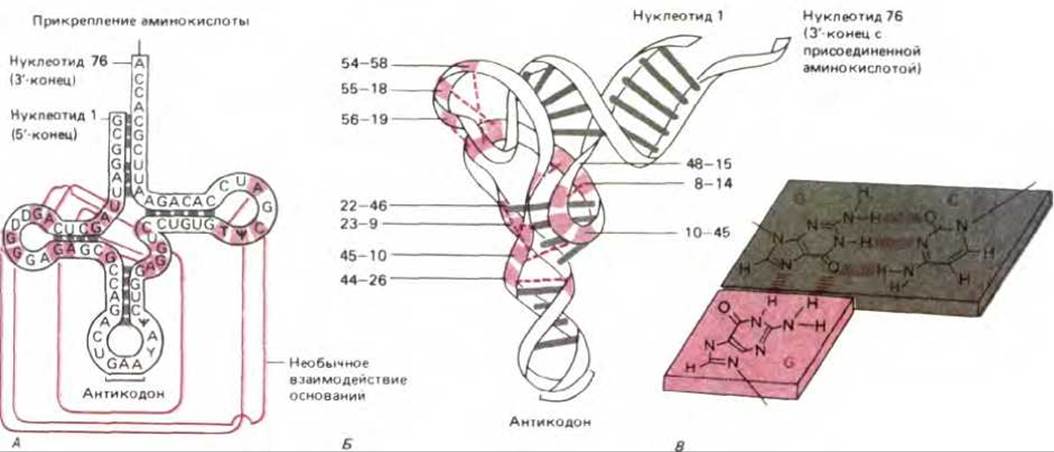
Рис. 3-16. Фенилаланиновая тРНК дрожжей. А. Молекула изображена в форме «кленового листа», чтобы показать комплементарное спаривание (выделено серым) внутри спиральных участков молекулы. Б. Схематическое изображение реальной формы молекулы, основанное на данных рентгеноструктурного анализа. Комплементарное спаривание обозначено серыми линиями. Нуклеотиды, участвующие в некомплементарном спаривании оснований, удерживающем вместе сегменты цепи, выделены цветом, а соответствующие пары оснований пронумерованы и связаны цветными пунктирными линиями, которые соответствуют цветным линиям в А. В. Необычное взаимодействие между парами оснований. Одно основание образует водородные связи с двумя другими; несколько таких «троек оснований» помогают свертывать эту молекулу тРНК.
Каждая молекула тРНК имеет характерную пространственную структуру, поддерживаемую теми же нековалентными взаимодействиями, которые удерживают вместе две цепи в двойной спирали ДНК. Однако в одноцепочечной молекуле тРНК комплементарное спаривание между нуклеотидными основаниями происходит в пределах одной цепи. Это приводит к тому, что молекула тРНК принимает определенную конформацию, существенную для выполнения функций адаптора. Четыре коротких сегмента молекулы образуют двухспиральную структуру, придающую молекуле вид двумерного «кленового листа». Этот кленовый лист в свою очередь упаковывается в многоскладчатую L-образную фигуру, которая скрепляется более сложными взаимодействиями на основе водородных связей (рис. 3-16). Два набора неспаренных нуклеотидных остатков по обоим концам «L» играют особенно важную роль для функционирования молекулы тРНК в биосинтезе белка: один из них образует антикодон, способный спариваться с комплементарным триплетом молекулы мРНК (кодоном); другой, имеющий последовательность ССА на 3'-конце молекулы, ковалентно связывается со специфической аминокислотой (рис. 3-16, А).
3.2.10. Считывание мРНК от одного конца до другого осуществляют рибосомы [18]
Перенос информации от мРНК к белку основан на том же принципе спаривания комплементарных оснований, что и перенос генетической информации от ДНК к ДНК и от ДНК к РНК (рис. 3-17). Однако процесс правильного расположения молекул тРНК на мРНК сложен и осуществляется рибосомами, комплексами, образованными более чем 50 различными белками, связанными с несколькими молекулами РНК (рРНК), выполняющими структурную роль. Каждая рибосома работает как большая биохимическая машина, на которой молекулы тРНК выстроены так, чтобы считывать закодированные в мРНК генетические инструкции. Сначала рибосома связывается со специальным участком молекулы мРНК и таким образом определяет рамку считывания и аминоконцевую аминокислоту белка. Затем рибосома по мере передвижения по молекуле мРНК транслирует кодон за кодовом, используя молекулы тРНК для последовательного присоединения аминокислот к растущему концу полипептидной цепи (рис. 3-18). Достигнув конца кодирующей части матрицы, рибосома и новосинтезированный карбоксильный конец белка отсоединяются от 3'-конца мРНК и переходят в цитоплазму клетки. Рибосомы работают очень эффективно: в 1 с одна бактериальная рибосома присоединяет к растущей полипептидной цепи 20 аминокислот. Подробнее структура рибосом и механизм синтеза белка описаны в гл. 5.

Рис. 3-17. Поток информации при синтезе белка. С участка в одной из цепей ДНК снимается комплементарная копия - матричная РНК. Затем нуклеотиды матричной РНК последовательно триплет за триплетом связывают комплементарные нуклеотиды антикодоновой петли определенных молекул тРНК. К противоположному концу каждой молекулы транспортной РНК (тРНК) прикреплена специфическая аминокислота и после спаривания эта аминокислота присоединяется к концу растущей белковой цепи. Таким образом, перевод последовательности нуклеотидов мРНК в последовательность аминокислот белка основан на комплементарном спаривании кодонов мРНК с антикодонами соответствующих молекул тРНК. Молекулярные основы переноса информации при трансляции оказываются аналогичными таковым при репликации и транскрипции ДНК. Заметим, что и синтез, и трансляция мРНК начинаются с 5'-конца.
3.2.11. Некоторые молекулы РНК функционируют как катализаторы [19]
Когда-то молекулы РНК рассматривались как цепочка нуклеотидов с относительно неинтересными химическими свойствами. В 1981 г. эта точка зрения была поколеблена открытием каталитической молекулы РЫК с такими изощренными химическими свойствами, которые биохимики раньше связывали только с белками. Рибосомные молекулы РНК ресничного простейшего Tetrahymena вначале были синтезированы как большая группа предшественников. Было показано, что одна из рРНК получается путем реакции сплайсинга РНК. Удивительным в этом открытии было то, что сплайсинг можно осуществить in vitro в отсутствие белка. Позже было показано, что сама интронная последовательность обладает ферментоподобной активностью и может катализировать двухступенчатую реакцию, показанную на рис. 3-19.
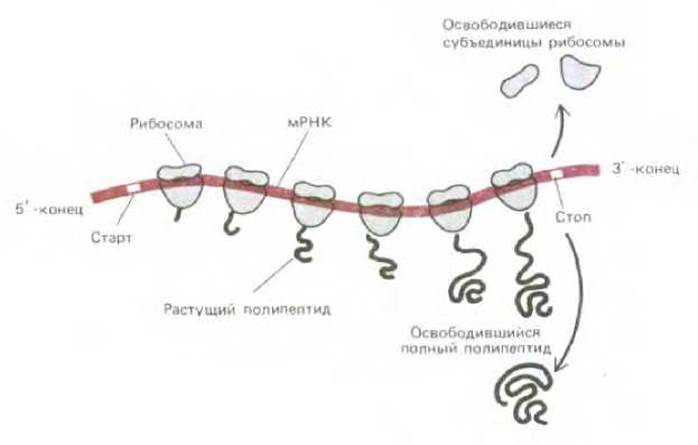
Рис. 3-18. Схема синтеза белка на рибосомах. Рибосомы присоединяются к стартовому сигналу вблизи 5'-конца молекулы мРНК и передвигаются к З'-концу, синтезируя по пути белок. Часто по одной молекуле мРНК движутся одновременно несколько рибосом, синтезируя несколько идентичных полипептидных цепей; такая структура в целом называется полирибосомой.
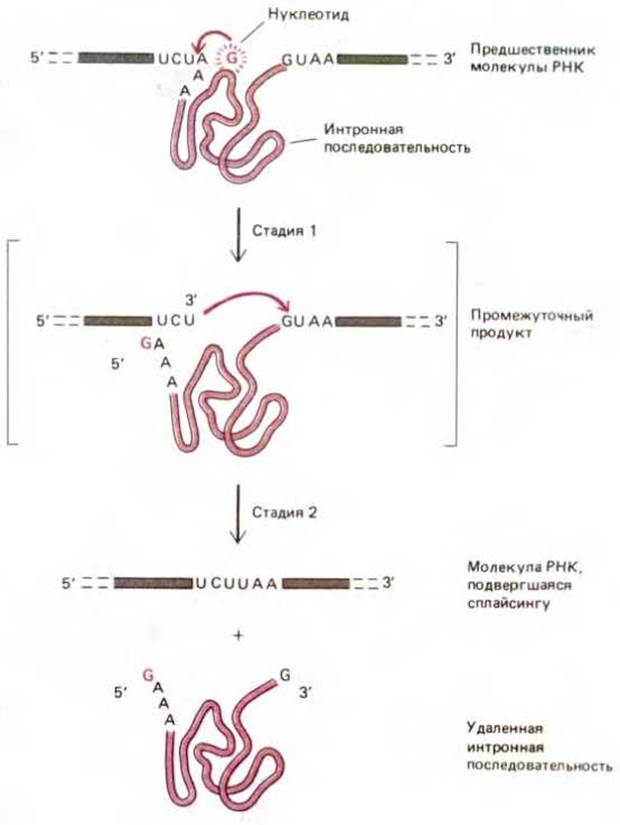
Рис. 3-19. Схема реакции самосплайсинга, при которой последовательность интрона катализирует собственное вырезание из молекулы рибосомной РНК у Tetrahymena. Реакция начинается с присоединения нуклеотида G к интронной последовательности, в результате чего происходит разделение цепи РНК. Затем вновь образованный 3'-конец цепи РНК подходит к другому концу и отделяет его, завершая реакцию.
В ходе дальнейших исследований было установлено, что синтезированная в пробирке интронная последовательность длиной в 400 нуклеотидов сворачивается с образованием структуры, способной функционировать как фермент в реакциях с другими молекулами РНК. Например, эта молекула способна связывать два специфических субстрата: нуклеотид гуанин и цепь РНК, и затем катализировать их ковалентное связывание, так что цепь РНК разрезается в специфической точке (рис. 3-20). РНК, способные к самосплайсингу и имеющие подобную структуру, были обнаружены в митохондриях грибов и в бактериальном вирусе (бактериофаг Т4).
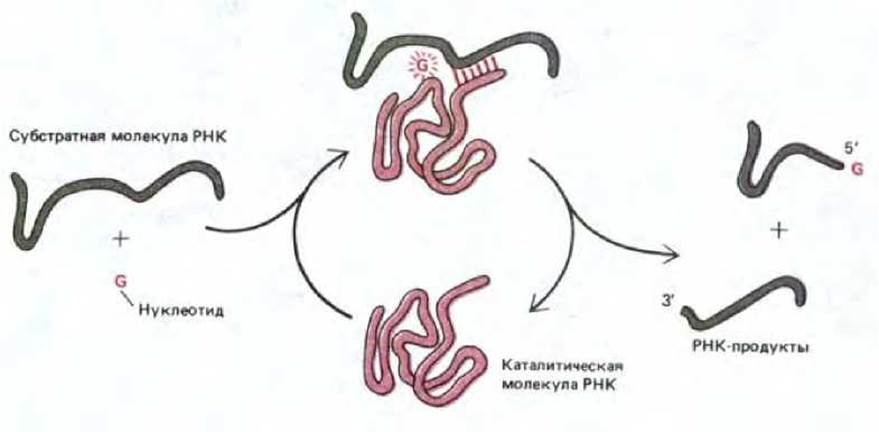
Рис. 3-20. Ферментоподобная реакция, катализируемая очищенной интронной последовательностью РНК у Tetrahymena. В этой реакции, которая соответствует первой стадии реакции, приведенной на рис. 3-19, и специфическая субстратная молекула РНК, и нуклеотид G тесно связываются с поверхностью каталитической молекулы РНК. Затем нуклеотид ковалентно связывается с субстратной молекулой РНК, разрезая ее в специфическом центре. Освобождение в результате этого двух цепей молекулы РНК дает возможность интронным последовательностям участвовать в дальнейших циклах реакции.
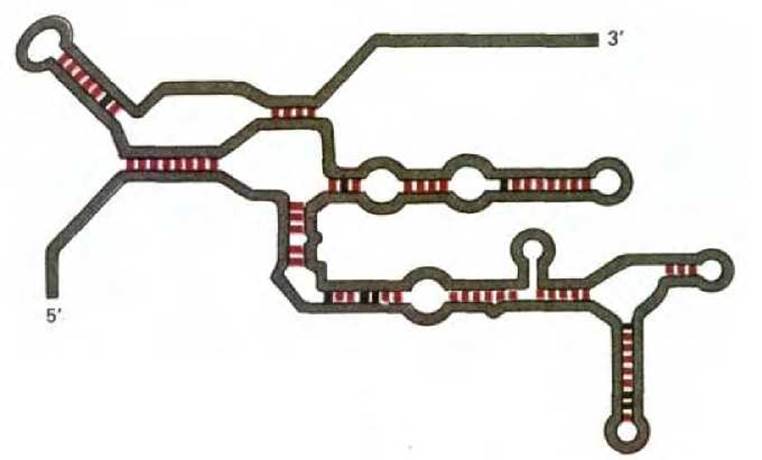
Рис. 3-21. Двумерное изображение каталитического остова интронной последовательности РНК, представленной на рис. 3-19 и рис. 3-20. Нормальные комплементарные пары оснований выделены цветом, а более слабые взаимодействия пар оснований показаны черным. Эта молекула содержит около 240 нуклеотидов; в нормальных условиях она свернута в плотную трехмерную структуру, но ее точная конформация неизвестна.
В этой модельной реакции, которая соответствует первому шагу реакции на рис. 3-19, та же интронная последовательность действует многократно, разрезая многие цепи РНК. Хотя обычно сплайсинг РНК проходит без автокатализа, самосплайсинг РНК, установленный у Tetrahymena, был открыт и в других типах клеток, включая грибы и бактерии. Это позволяет предположить, что такие последовательности РНК могли возникнуть до расхождения родословных эукариот и прокариот около 1,5 млрд. лет назад.
В последнее время были открыты некоторые другие семейства каталитических РНК. Например, большинство тРНК изначально синтезировались как предшественники РНК, затем было показано, что одна молекула РНК играет основную каталитическую роль в РНК-белковом комплексе, распознавая эти предшественники и разрезая их в специфических точках. Катализирующая последовательность РНК играет также важную роль в жизненном цикле многих растительных вирусов, подобная последовательность обнаружена в РНК лягушки, хотя ее роль в данном случае не доказана. Более примечательно то, что обнаружение катализа на основе РНК дает теперь основания подозревать, что рибосомы обладают более широкими функциями, чем предполагалось. Весьма вероятно, что рибосомные белки играют второстепенную роль по сравнению с рибосомными РНК, которые составляют более половины массы рибосомы.
Каким образом молекулы РНК могут действовать наподобие ферментов? Пример тРНК показывает, что молекулы РНК могут складываться высокоспецифичным образом. Предложенная двумерная структура остова интронной последовательности Tetrahymena, способной к самосплайсингу, представлена на рис. 3-21. Взаимодействия между разными участками этой молекулы РНК (аналогичные необычным водородным связям в молекулах тРНК - см. рис. 3-16) ответственны за ее дальнейшее сворачивание с образованием сложной трехмерной поверхности с каталитическими свойствами. Необычное взаимное расположение атомов может деформировать ковалентные связи и, следовательно, придавать отдельным атомам в свернутой цепи РНК необычную реакционноспособность.
Как указывалось в гл. 1, открытие каталитических молекул РНК в корне изменило наши представления о происхождении первых живых клеток (см. разд. 1.З.4.).
Заключение
Генетическая информация записана в линейной последовательности нуклеотидов ДНК. Каждая молекула ДНК состоит из двух комплементарных полинуклеотидных цепей, удерживаемых вместе водородными связями, образующими GC- и АТ-пары оснований. Репликация ДНК, обеспечивающая удвоение генетической информации, происходит путем образования новой комплементарной цепи на каждой из исходных цепей.
Экспрессия генетической информации, заключенной в ДНК, осуществляется путем трансляции линейной последовательности нуклеотидов в колинеарную последовательность аминокислот белка. Сначала ограниченный участок ДНК копируется на комплементарную цепь РНК. Этот первичный транскрипт РНК подвергается сплайсингу для удаления интронных последовательностей и превращается в молекулу мРНК. В конце концов мРНК транслируется с образованием белка путем сложного набора реакций, происходящих в рибосоме. Вначале аминокислоты, используемые для синтеза белка, прикрепляются к семейству молекул тРНК, каждая из которых путем комплементарного спаривания оснований узнает набор из трех нуклеотидов мРНК. Последовательность нуклеотидов мРНК считывается с одного конца к другому триплетами нуклеотидов в соответствии с универсальным генетическим кодом.
Другие молекулы РНК в клетках используются как ферментоподобные катализаторы. Эти молекулы РНК сворачиваются с образованием такой структуры, в которой некоторые нуклеотиды поверхности могут стать необычно активными.