Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Молекулярная организация клеток
Основные генетические механизмы
Вирусы, плазмиды и транспозоны
Знакомясь с основными генетическими механизмами, мы до сих пор сосредоточивали свое внимание на селективных преимуществах, которые они обеспечивают клетке. Мы убедились в том, что клетке для выживания совершенно необходимо сохранение генетической информации путем репарации ее ДНК, а для размножения клеток столь же необходимо быстрое и точное ее воспроизведение. Выживание вида в целом связано, как известно, с появлением новых, более приспособленных генетических вариантов, а оно в значительной мере облегчается перегруппировкой генов и случайным перераспределением последовательностей ДНК в результате генетической рекомбинации. Теперь нам предстоит познакомиться с группой генетических элементов, которые ведут себя как паразиты, т.е. используют генетические механизмы клетки для своих собственных нужд.
При некоторых условиях определенные нуклеотидные последовательности ДНК могут воспроизводиться независимо от остального генома. По степени независимости от клетки-хозяина эти последовательности весьма существенно различаются. Наиболее независимы хромосомы вирусов, потому что у вирусов есть белковые оболочки, благодаря которым они могут свободно переходить из клетки в клетку. В той или иной степени близки к вирусам (хотя и более зависимы от клетки) нуклеотидные последовательности, называемые плазмидами и транспозонами; у них нет белковой оболочки, и потому они могут реплицироваться только в одной-единственной клетке или в ее потомстве. Еще более примитивны некоторые последовательности ДНК, которые можно предположительно рассматривать как мобильные генетические элементы на том основании, что в хромосомах клеток они встречаются во многих копиях, однако эти последовательности перемещаются или размножаются столь редко, что трудно решить, представляют ли они собой вообще отдельные генетические элементы.
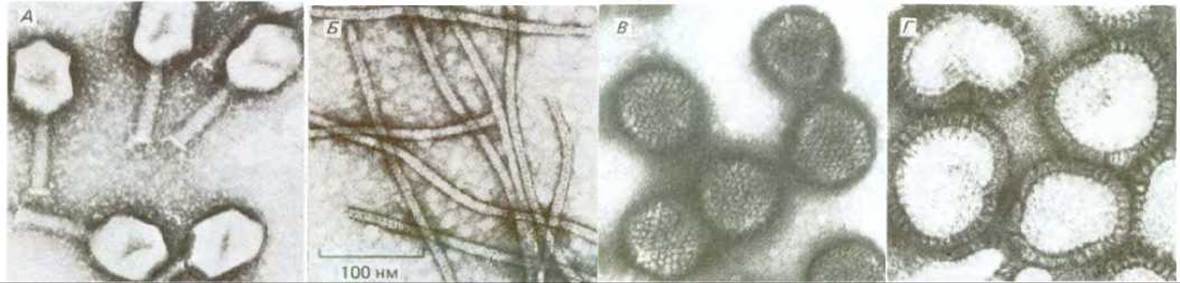
Рис. 5-68. Электронные микрофотографии вирусных частиц (метод негативного контрастирования). Все вирусы сфотографированы при одном и том же увеличении, масштаб указан на фото Б. А. Бактериофаг Т4-крупный ДНК-содержащий вирус, заражающий бактерию Е. coli. ДНК находится в головке бактериофага и впрыскивается в бактериальную клетку при помощи его цилиндрического хвоста. (С любезного разрешения James Paulson.) Б. Вирус X картофеля. Нитевидные частицы этого вируса растений содержат РНК-геном. (С любезного разрешения Graham Hills.) В. Аденовирус - ДНК-содержащий вирус, заражающий различные клетки человека. Снаружи вирусные частицы окружены белковой оболочкой - капсидом. (С любезного разрешения Mei Lie Wong.) Г. Вирус гриппа крупный РНК-содержащий вирус животных. Помимо белкового капсида у него имеется еще мембранная оболочка липидный бислой с выступающими из него включениями вирусного гликопротеина. (С любезного разрешения R. С. Williams, H. W. Fisher.)
Все такие псевдонезависимые элементы могут размножаться, лишь всемерно используя метаболизм клетки-хозяина, поэтому они служат удобным инструментом для исследования нормальных клеточных механизмов. Мы начнем свое обсуждение с вирусов, так как из всех мобильных генетических элементов они изучены наиболее полно. Затем мы рассмотрим свойства плазмид и транспозонов, которые иногда удивительно сходны с вирусами и, быть может, являются их предками.
5-39
5.5.1. Вирусы - это мобильные генетические элементы [43]
Вирусы впервые были описаны как болезнетворные агенты, которые размножаются только в клетках и имеют настолько малые размеры, что способны проходить через ультратонкие фильтры, задерживающие самые мелкие бактерии. До появления электронного микроскопа природа их оставалась неясной, хотя уже тогда высказывалось мнение, что это, возможно, просто гены, которые приобрели способность переходить из одной клетки в другую. В 1930-х годах использование ультрацентрифуги сделало возможным отделение вирусов от компонентов клетки-хозяина. В результате уже в начале 1940-х годов стало более или менее ясно, что все вирусы содержат нуклеиновые кислоты. Это укрепило исследователей в мысли, что вирусы и генетический материал выполняют сходные функции. Подтверждение такой точки зрения было получено при изучении вирусов бактерий (бактериофагов). В 1952 г. удалось показать, что в клетку бактерии-хозяина проникает одна только ДНК бактериофага (без его белка) и что именно она инициирует здесь процесс репликации, приводящий в конечном счете к появлению в инфицированной клетке нескольких сотен дочерних вирусных частиц. Таким образом, вирусы можно рассматривать как генетические элементы, одетые в защитную оболочку и способные переходить из одной клетки в другую. Размножение вирусов само по себе часто оказывается летальным для клетки, в которой оно происходит. Многие вирусы разрушают инфицированную клетку (вызывают ее лизис), что и дает возможность потомству вируса переходить в соседние клетки. Клинические симптомы вирусной инфекции во многих случаях отражают именно эту цитолитическую способность вируса. Высыпание при инфекции вирусом герпеса или оспенная сыпь отражают, например, гибель эпителиальных клеток на отдельных пораженных участках кожи. Строение оболочки вируса, тип его нуклеиновой кислоты, способ проникновения в клетку и механизм размножения в клетке - все эти свойства у разных вирусов сильно варьируют. Электронные микрофотографии на рис. 5-68 дают представление о некоторых структурных различиях между вирусами.
5.5.2. Вирус заключен в белковый капсид или в мембранную оболочку [44]
Первоначально предполагалось, что наружная оболочка вирусов построена из белковых молекул какого-нибудь одного типа. Считалось, что вирусные инфекции начинаются с разъединения внутри клетки-хозяина хромосомы (т. е. нуклеиновой кислоты) вируса и его белковой оболочки. Далее следует самовоспроизведение хромосомы с образованием большого числа ее копий и синтез вирусоспецифичных белков оболочки. Затем происходит образование дочерних вирусных частиц в результате спонтанной сборки белковой оболочки вокруг дочерних вирусных хромосом (рис. 5-69).
Теперь мы отдаем себе отчет в том, что эти первые описания, хотя и верные в общем виде, давали упрощенное представление о чрезвычайно разнообразных жизненных циклах вирусов. Во-первых, белковая оболочка (или капсид) почти у всех вирусов состоит из полипептидных цепей более чем одного типа, причем эти цепи нередко сгруппированы в несколько слоев. Во-вторых, у многих вирусов их белковый капсид окружен еще и мембраной, в которую помимо белка входят и липиды. У многих из этих вирусов сборка наружной оболочки происходит в плазматической мембране клетки-хозяина, и дочерние вирусные частицы выходят наружу, отпочковываясь от этой плазматической мембраны (рис. 5-70). Отпочковывание позволяет потомству вируса покидать клетку, не нарушая ее плазматической мембраны, т. е. не убивая клетку. Липидные компоненты мембраны вируса идентичны липидам плазматической мембраны клетки-хозяина, тогда как белки, присутствующие в липидном бислое, вирусоспецифичны. Сборку вирусной мембраны в плазматической мембране клетки-хозяина мы обсудим в гл. 8, что же касается сборки белкового капсида вируса, то ее иллюстрирует рис. 3-43.
5-45
5.5.3. Геномы вирусов представлены разнообразными формами, а генетическим материалом у них может быть как ДНК, так и РНК [45]
Когда удалось расшифровать структуру ДНК, естественно было сделать вывод, что вся генетическая информация хранится именно в этой двухспиральной форме, поскольку ее преимущества в поддержании стабильности ДНК и в смысле возможности репарации казались совершенно неоспоримыми. Случайное повреждение одной из полинуклеотидных цепей, действительно, всегда может быть исправлено при помощи комплементарной цепи. Однако это преимущество представляется несущественным, когда речь идет о крошечных вирусных хромосомах, насчитывающих всего несколько тысяч нуклеотидов, - вероятность их случайного повреждения очень невелика в сравнении с риском, которому подвергается клеточный геном, содержащий миллионы нуклеотидов.
По этой причине генетическая информация вирусов может храниться в разнообразных и необычных формах, в частности в форме РНК.
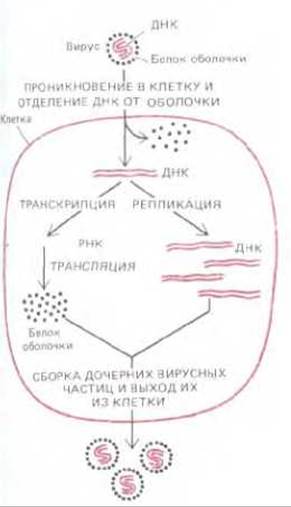
Рис. 5-69. Простейший жизненный цикл вируса. Представленный здесь гипотетический вирус содержит небольшую двухцепочечную молекулу ДНК, кодирующую единственный вирусный белок, из которого построен капсид вируса. Вирусов с таким простым строением мы не знаем.
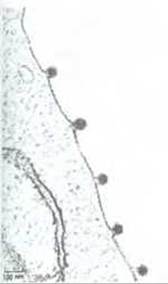
Рис. 5-70. Электронная микрофотография тонкого среза животной клетки, от которой отпочковывается несколько частиц вируса, заключенного в мембранную оболочку (вирус леса Семлики). Геном этого вируса представлен одноцепочечной РНК. (С любезного разрешения М. Olsen, G. Griffiths.)

Рис. 5-71. Схематическое изображение (масштабы не соблюдены) различных типов вирусных геномов. У самых мелких вирусов геном состоит всего из нескольких генов и генетическим материалом может быть у них как ДНК, так и РНК; у наиболее крупных вирусов геном всегда представлен двухцепочечной ДНК, включающей сотни генов. Такие особенности хромосом, как кольцевая форма молекул ДНК или особое строение концов цепей у линейных молекул, позволяют вирусам избежать трудностей, связанных с репликацией нескольких последних нуклеотидов на конце цепи ДНК.
Роль хромосомы вируса может играть одноцепочечная РНК (вирус табачной мозаики), двухцепочечная спираль РНК (реовирус), кольцевая одноцепочечная ДНК (бактериофаги М13 и ФХІ74) или линейная одноцепочечная ДНК (парвовирусы). Правда, первыми среди хромосом вирусов были изучены простые линейные двойные спирали ДНК, но потом выяснилось, что столь же часто встречаются кольцевые двойные спирали или более сложные формы линейных двойных спиралей ДНК. Есть вирусы, у которых к 5'-концам цепей ДНК ковалентно присоединен белок, а у очень крупных вирусов оспы две комплементарные цепи ДНК на одних концах соединены фосфодиэфирными связями (рис. 5-71). Для каждого типа вирусного генома характерна своя особая энзимология репликации, а потому он должен нести в себе информацию не только о белках, образующих оболочку вируса, но также и об одном или нескольких ферментах, необходимых для репликации вирусной нуклеиновой кислоты.
5-40
5.5.4. Хромосома вируса содержит информацию для синтеза ферментов, участвующих в репликации вирусной нуклеиновой кислоты [46]
Количество информации, которую вирус привносит в инфицируемую клетку, чтобы обеспечить себе воспроизведение, различается у разных вирусов весьма заметно. Так, в ДНК сравнительно крупного бактериофага Т4 закодировано не менее 30 различных ферментов, обеспечивающих избирательную и быструю репликацию хромосомы бактериофага Т4 в ущерб репликации ДНК клетки-хозяина, т. е. Е. coli (рис. 572). Эти белки участвуют в непрерывных циклах репликации Т4-ДНК и осуществляют избирательное включение 5-гидроксиметилцитозина, который в Т4-ДНК замещает цитозин. В геноме бактериофага Т4 закодированы также и нуклеазы, избирательно разрушающие ДНК Е. coli (геном самого бактериофага из-за необычного состава оснований не подвержен действию этих нуклеаз). Кроме того, в нем закодированы белки. изменяющие молекулы бактериальной РНК-полимеразы таким образом, что они на разных стадиях инфекции транскрибируют различные группы генов бактериофага.
Более мелкие ДНК-содержащие вирусы, например обезьяний вирус SV40 или мельчайший бактериофаг ФХІ74, несут в себе гораздо меньше генетической информации и гораздо больше зависят от ферментов клетки-хозяина как в синтезе своих белков, так и в синтезе своей ДНК. Они подчиняют себе и используют для своих нужд клеточные ферменты, участвующие в репликации ДНК, в том числе и ДНК-полимеразу.
Однако даже в геноме самых мелких ДНК-вирусов закодированы ферменты, избирательно инициирующие синтез их собственной ДНК, для чего они узнают на хромосоме вируса особую нуклеотидную последовательность - точку начала репликации. Это существенно, потому что вирус может успешно размножаться лишь при том условии, если ему удастся игнорировать регуляторные сигналы клетки, которые в противном случае не дадут вирусной ДНК удваиваться более чем один раз в каждом клеточном цикле. Мы до сих пор не знаем, как эукариотические клетки регулируют синтез своей ДНК, и можно надеяться, что знакомство с механизмами, при помощи которых вирусы избавляются от этой регуляции (а их изучать, разумеется, гораздо легче), даст нам ключ к пониманию регуляторных механизмов клетки-хозяина.
К репликации РНК-вирусов предъявляются особые требования, поскольку для воспроизведения своего генома они должны копировать молекулы РНК, что означает полимеризацию нуклеозидтрифосфатов на РНК-матрице. Клетки, как правило, не располагают ферментами для осуществления этой реакции, поэтому даже в мельчайших вирусах должны быть закодированы их собственные РНК-зависимые полимеразы нуклеиновых кислот.
Рассмотрим теперь более подробно механизмы репликации разных типов вирусов.
5-42
5.5.5. РНК-вирусы и ДНК-вирусы реплицируются путем образования комплементарных цепей [47]
Репликация геномов РНК-вирусов точно так же, как и репликация ДНК, связана с образованием комплементарных полинуклеотидных цепей. У большинства РНК-вирусов этот процесс катализируют РНК-зависимые РНК-полимеразы (репликазы), кодируемые РНК-хромосомой вируса. Эти ферменты часто включаются в дочерние вирусные частицы, и тогда при вирусной инфекции они уже сразу имеются в наличии, т.е. могут немедленно начинать репликацию вирусной РНК. У так называемых вирусов с негативным геномом, к которым принадлежат, в частности, вирусы гриппа и везикулярного стоматита, репликазы всегда включаются в капсид. Вирусы этой группы называются так потому, что у них инфицирующая цепь не кодирует никаких белков; только комплементарная ей цепь несет необходимые для этого нуклеотидные последовательности. Таким образом, инфицирующая цепь не может индуцировать размножение вируса без предобразованной репликазы. У РНК- вирусов с позитивным геномом, например у вируса полиомиелита, дело обстоит иначе: здесь вирусная РНК может выступать в роли мРНК, и у этих вирусов «голый» геном инфекционен.
Синтез вирусной РНК начинается всегда на 3'-конце РНК-матрицы (т. е. с 5'-конца новой молекулы РНК) и идет до тех пор, пока не будет достигнут 5'-конец матрицы. Никаких механизмов, которые корректировали бы синтез вирусной РНК, нет, и частота ошибок здесь примерно та же, что и при транскрипции ДНК (в среднем одна ошибка на 104 нуклеотидов). Однако отсутствие корректирующих механизмов серьезным образом не сказывается на репликации из-за небольших размеров РНК-хромосомы вируса. Геномы всех РНК-вирусов невелики в сравнении с геномами крупных ДНК-содержащих вирусов, и это является прямым следствием того, что у них механизм репликации более примитивен.
У всех ДНК-вирусов репликация инициируется в точке начала репликации, к которой присоединяются специальные инициаторные белки, способные привлекать к себе репликационные ферменты клетки-хозяина (см. разд. 5.3.9). Существует много различных путей репликации вирусных геномов. Их сложность объясняется необходимостью так или иначе решить проблему репликации концов простой линейной молекулы ДНК, при том что имеющаяся ДНК-репликаза не способна начинать синтез в отсутствие затравки. Проблема эта решается у ДНК-вирусов различными способами; есть вирусы с кольцевой хромосомой, которая вообще не имеет концов; у некоторых вирусов с линейным геномом терминальные нуклеотидные последовательности повторяются или концы хромосом образуют петли; наконец, имеются вирусы, геном которых несет особые терминальные белки, способные непосредственно «включать» ДНК-полимеразу.

Рис. 5-72. Хромосома бактериофага Т4, на которой обозначены больше 30 генов, участвующих в репликации его ДНК. Геном бактериофага Т4 насчитывает свыше 160000 пар нуклеотидов, кодирующих более 200 различных белков, в том числе и белков, участвующих в репликации ДНК (некоторые из них здесь отмечены). Среди остальных белков много таких, которые участвуют в сборке головки и хвостового отростка (см. рис. 568, А).
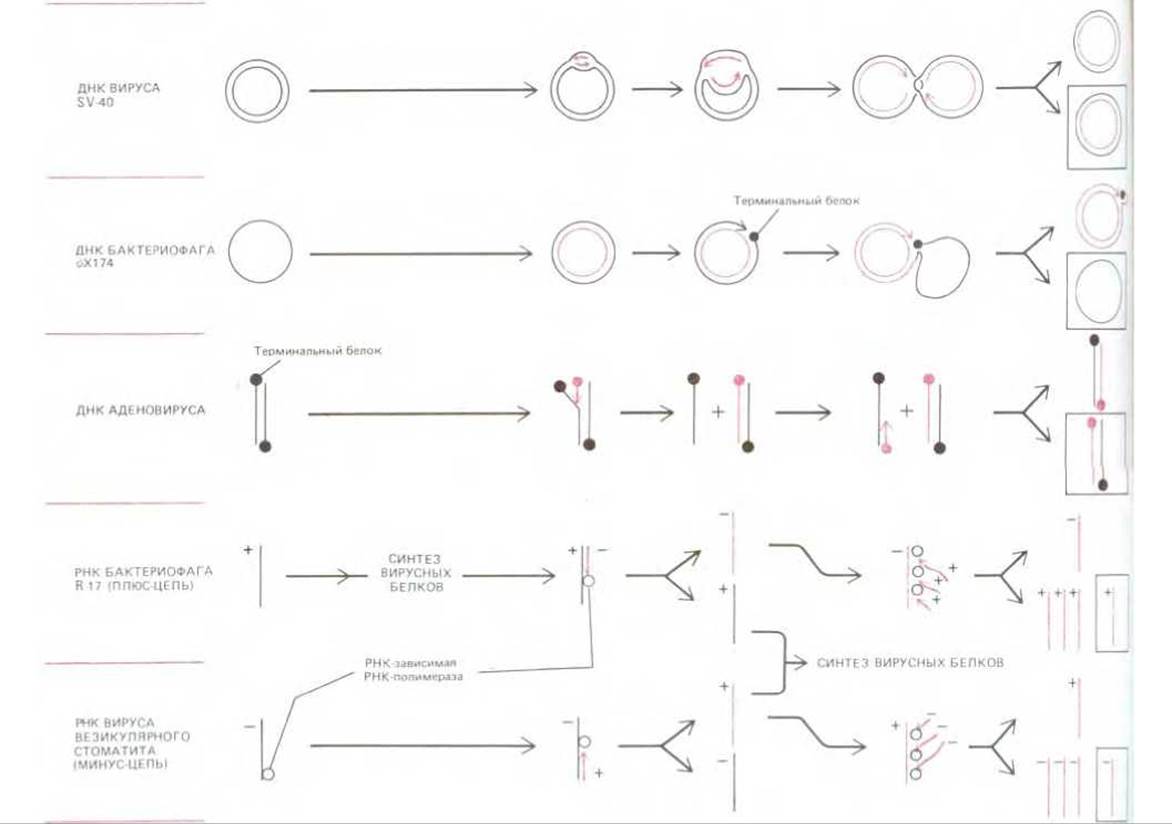
Рис. 5-73. Примеры, иллюстрирующие разнообразные способы репликации вирусных геномов. В двух случаях, как мы видим, к концам цепей ДНК ковалентно присоединены терминальные белки, эти белки играют важную роль в соответствующем процессе репликации. Обратите внимание на главное различие между РНК-вирусами с позитивными и негативными геномами: оно заключается в том, что вирусы с «минус»-цепью, прежде чем образовать вирусные белки, должны синтезировать «плюс»-цепь. Для этой цели капсид РНК-вируса должен нести в себе одну или несколько молекул вирусной РНК-зависимой РНК-полимеразы (репликазы). Справа в рамке показан для каждого случая конечный РНК- или ДНК-продукт, идентичный инфицирующему вирусному геному, представленному слева.
На рис. 5-73 представлены некоторые способы репликации геномов, встречающиеся у вирусов.
5-43
5.5.6. Хромосомы вирусов способны включаться в хромосомы клетки-хозяина [48]
Проникновение вирусной хромосомы в клетку далеко не всегда приводит к немедленному размножению вирусных частиц. Многие вирусы могут существовать в латентной форме: их геномы при этом хотя и присутствуют в клетке, остаются неактивными; образования дочерних
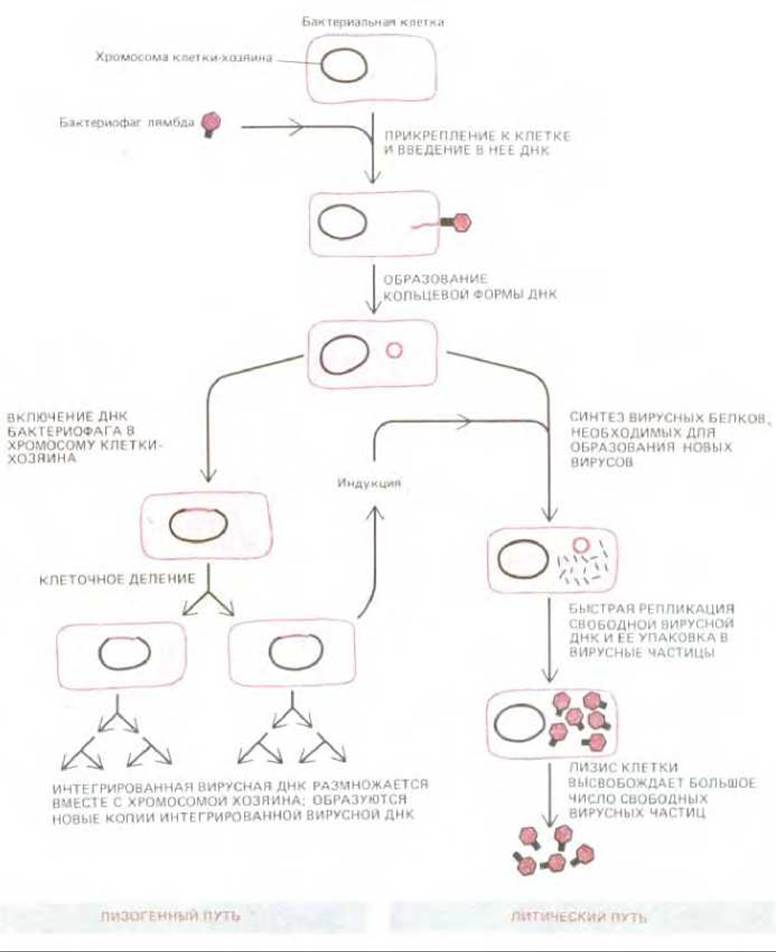
Рис. 5-74. Жизненный цикл бактериофага λ. Геном бактериофага состоит приблизительно из 50000 пар нуклеотидов и кодирует около 50 белков. Его двухцепочечная ДНК способна существовать как в линейной, так и в кольцевой форме. Развитие бактериофага может пойти, как это здесь изображено, и по литическому, и по лизогенному пути. Повреждение ДНК клетки, находящейся в лизогенном состоянии, вынуждает интегрированную фаговую ДНК (профаг) выйти из хромосомы хозяина и начать литический цикл. Включение фаговой ДНК в хромосому хозяина и выход из хромосомы осуществляются путем сайт-специфической генетической рекомбинации, катализируемой особым белком бактериофага λ, так называемой интегразой.
частиц не происходит. О способности вирусов существовать в латентной форме стало известно, когда выяснилось, что многие бактерии, казалось бы неинфицированные, продуцируют бактериофаг, если их подвергнуть облучению ультрафиолетом. После этого были проведены эксперименты, которые показали, что у таких лизогенных бактерий в хромосому включена полная хромосома вируса. Эти интегрированные вирусные хромосомы стали называть провирусами (профагами).
Бактериофаги, способные включаться в бактериальные хромосомы, называются лизогенизирующими бактериофагами. Наиболее полно изучен среди них бактериофаг лямбда (λ), о ферменте которого, лямбда-интегразе, мы уже говорили. Когда бактериофаг λ заражает подходящую клетку Е. coli, он обычно размножается в ней и образует несколько сотен дочерних фаговых частиц, которые выходят наружу в момент лизиса клетки; это так называемый литический путь инфекции. Гораздо реже линейные инфицирующие молекулы ДНК замыкаются в кольцо и включаются в кольцевую хромосому бактерии-хозяина путем сайт-специфической рекомбинации (см. разд. 5.4.7). По завершении такой интеграции образовавшаяся лизогенная бактерия, несущая хромосому бактериофага X в виде профага, размножается, как обычно, до тех пор, пока на нее не воздействует какой-нибудь повреждающий внешний фактор, например ультрафиолетовые лучи или ионизирующее излучение. Это воздействие вынуждает интегрированный профаг выйти из хромосомы хозяина и начать обычный, свойственный вирусу цикл размножения. Интегрированный профаг, таким образом, не осужден на гибель вместе с поврежденной клеткой-хозяином; у него есть шанс ускользнуть из нее и перейти в соседнюю неповрежденную клетку Е. coli (рис. 5-74).
5.5.7. Непрерывный синтез вирусных белков может превращать нормальные клетки в раковые [49]
В животных клетках, как и у бактерий, для размножения вирусов существует помимо литического еще и другой путь. Те животные клетки, в которых ДНК-вирусы размножаются литическим путем, ведущим к гибели клетки, принято называть пермиссивными. Клетки, в которых размножение вирусов блокируется, называются непермиссивными; вирусная хромосома в таких случаях либо включается в геном клетки-хозяина и в дальнейшем размножается вместе с ним, либо образует плазмиду - кольцевую молекулу ДНК, репликация которой регулируется и не ведет к гибели клетки. Иногда это вызывает в непермиссивных клетках определенное генетическое изменение, в результате которого начинается их неконтролируемый рост, т.е. нормальные клетки превращаются в раковые. Соответствующий ДНК-вирус называют в таких случаях опухолевым ДНК-вирусом, а превращение, о котором идет речь, - вирусной неопластической трансформацией. Среди опухолевых ДНК-вирусов наиболее полно изучены два представителя паповавирусов, а именно SV40 и вирус полиомы. Выяснилось, что их трансформирующая способность зависит от нескольких вирусных белков, кооперативное действие которых переводит покоящиеся клетки из Go-фазы в S-фазу (см. разд. 3.3). В пермиссивных клетках этот переход в S-фазу делает доступными вирусу все репликационные ферменты клетки-хозяина, необходимые для синтеза вирусной ДНК. В непермиссивной клетке синтез провирусом этих вирусных белков подавляет часть нормальных регуляторных механизмов и самой клетки, и всего ее потомства.
5.5.8. Опухолевые РНК-вирусы принадлежат к классу ретровирусов [50]
Особым образом ведут себя опухолевые РНК-вирусы: проникновение их в пермиссивную клетку часто ведет одновременно и к нелетальному для клетки высвобождению дочерних вирусных частиц (отпочковывающихся от клеточной поверхности), и к стойкому генетическому изменению в инфицированной клетке, которое превращает эту клетку в раковую. Как может заражение вирусом вызвать стойкое генетическое изменение, было непонятно до тех пор, пока не был открыт фермент обратная транскриптаза, с помощью которого цепи инфицирующей РНК этих вирусов транскрибируются в комплементарные им цепи ДНК. Опухолевые РНК-вирусы, к которым относится первый хорошо изученный опухолевый вирус, а именно вирус саркомы Рауса, являются представителями крупного класса вирусов, так называемых ретровирусов. Название это отражает тот факт, что часть их жизненного цикла составляет процесс, обратный нормальной транскрипции, т. е. транскрипции ДНК в РНК. К ретровирусам относится и вирус СПИДа (спонтанно приобретенного иммунодефицита).
Рис. 5-75 иллюстрирует жизненный цикл одного из ретровирусов. Фермент обратная транскриптаза представляет собой необычную форму ДНК-полимеразы, способную использовать в качестве матрицы и РНК. и ДНК; этот фермент закодирован в РНК ретровируса и при образовании дочерних вирусных частиц всегда упаковывается в их капсид. Когда одноцепочечная РНК ретровируса проникает в клетку, обратная транскриптаза сначала синтезирует ДНК-копию этой цепи РНК, в результате чего образуется гибридная ДНК-РНК-спираль, которую тот же фермент использует затем для образования двойной спирали, состоящей уже из двух цепей ДНК. Эта ДНК-копия вирусного РНК-генома включается в хромосому клетки-хозяина. Включению способствует закодированный вирусом фермент, осуществляющий сайт-специфическую рекомбинацию; он узнает определенную нуклеотидную последовательность в вирусной ДНК и катализирует включение вирусной ДНК практически в любой участок хромосомы клетки-хозяина (см. рис. 5-67, Б). Следующий этап инфекционного процесса составляет транскрипция интегрированной вирусной ДНК РНК-полимеразой клетки-хозяина; в результате появляется большое число молекул вирусной РНК, идентичных исходному инфицирующему геному. Завершается процесс трансляцией этих молекул РНК с образованием капсидных и мембранных белков, а также обратной транскриптазы; наконец, происходит сборка новых вирусных частиц, окруженных оболочкой, которые отпочковываются от клеточной мембраны (см. рис. 5-75).
Как РНК-, так и ДНК-содержащие опухолевые вирусы вызывают неопластическую трансформацию клеток, потому что присутствие в клетке вирусной ДНК индуцирует синтез новых белков, нарушающих регуляцию клеточного деления. Гены, кодирующие синтез таких белков, называются онкогенами. У опухолевых ДНК-вирусов онкогены обычно кодируют нормальные вирусные белки, необходимые для размножения вируса. Иначе обстоит дело у опухолевых РНК-вирусов: онкогены, которые они несут, представляют собой модифицированные формы нормальных генов клетки-хозяина - они для размножения вируса не требуются. Поскольку в капсид ретровируса может уместиться лишь некоторое ограниченное количество РНК, необходимые онкогенные последовательности нуклеотидов часто замещают собой существенную часть генома ретровируса и вирус оказывается дефектным. Мы расскажем позже (см. разд. 13.4.2 и разд. 21.2.1), почему изучение вирусных онкогенов послужило ключом к пониманию причин и природы рака, а также к познанию тех механизмов, которые в норме регулируют рост и деление клеток у многоклеточных организмов.
5.5.9. Некоторые транспозоны очень сходны с ретровирусами [51]
Поскольку многие вирусы способны включаться в хромосомы клеток-хозяев и выходить из них, можно предположить, что все крупные геномы содержат то или иное число различных провирусов. Вполне вероятно, что в этих геномах имеется также и ряд мобильных последовательностей ДНК, которые не образуют вирусных частиц и не могут покинуть клетку; их называют транспозонами. Транспозоны (размеры их колеблются от нескольких сотен до десятков тысяч нуклеотидных пар) обычно присутствуют в клетке в нескольких копиях. Они представляют собой нечто вроде крошечных паразитов, таящихся в хромосомах. Время от времени любой такой транспозон активируется и под влиянием собственного фермента, катализирующего сайт-специфическую рекомбинацию, переходит в какой-нибудь иной участок ДНК в пределах той же клетки. Этот процесс носит название транспозиции. Ферменты, катализирующие транспозицию, - их называют транспозазами - по большей части закодированы в ДНК самого транспозона. Большинство транспозонов переходит с места на место крайне редко (у бактерий приблизительно 1 раз на 105 генераций), так что их очень трудно отличить от неподвижных участков хромосомы. Не знаем мы также и того, что служит пусковым механизмом для этого процесса.
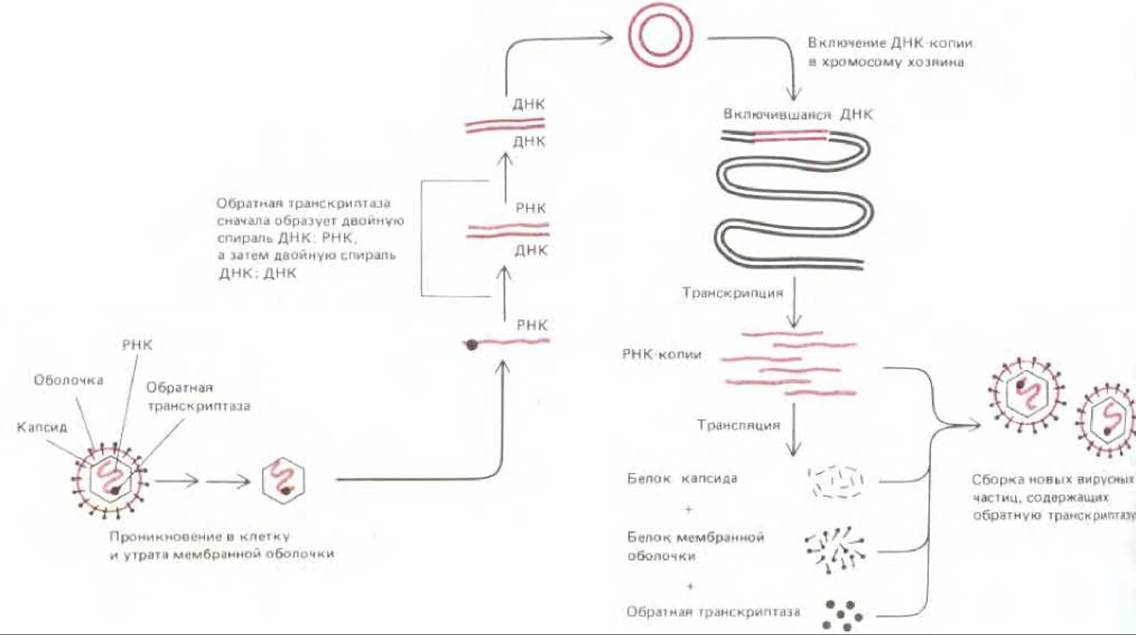
Рис. 5-75. Жизненный цикл одного из ретровирусов. Геном ретровируса представляет собой молекулу РНК, насчитывающую около 8500 нуклеотидов; в каждой вирусной частице упаковано две такие молекулы. Фермент обратная транскриптаза представляет собой ДНК-полимеразу, которая образует сначала ДНК-копию вирусной молекулы РНК, а затем вторую цепь ДНК, так что в итоге получается ДНК-копия РНК-генома. Включение этой двойной спирали ДНК в хромосому клетки-хозяина, катализируемое вирусным белком, необходимо для синтеза новых молекул вирусной РНК, осуществляемого РНК-полимеразой клетки-хозяина.
Механизмы транспозиции могут быть различными. У одного крупного семейства транспозонов используемый для этой цели механизм полностью совпадает с частью жизненного цикла ретровируса. Такие элементы - их называют ретротранспозонами - известны у столь далеких друг от друга организмов, как дрожжи, плодовая мушка и млекопитающие. Одним из наиболее хорошо изученных ретротранспозонов является обнаруженный у дрожжей элемент Туl. На первом этапе его транспозиции происходит транскрипция всего элемента с образованием РНК-копии, состоящей более чем из 5000 нуклеотидов. В этом РНК-транскрипте закодирована обратная транскриптаза, которая синтезирует кольцевую двухцепочечную ДНК-копию молекулы РНК через промежуточное соединение - гибридную двойную спираль РНК-ДНК точно так же, как это происходит в клетке, инфицированной ретровирусом (см. рис. 5-75). Аналогия сохраняется и далее, когда кольцевая ДНК включается в случайно выбранный участок хромосомы. Как и у ретровируса, включение осуществляется по типу сайт-специфической рекомбинации, представленной на рис. 5-67, Б; для этого, по всей вероятности, используется транспозаза, также закодированная в упомянутом выше длинном РНК-транскрипте. При всем поразительном сходстве с ретровирусом элемент Туl отличается от него тем, что будучи лишен функциональной белковой оболочки, он может перемещаться лишь внутри какой-нибудь одной клетки или может быть передан ее потомкам.
5-44
5.5.10. У другой группы транспозонов переход совершается непосредственно из одного участка генома в другой [52]
Многие транспозоны отличаются от ретротранспозонов тем, что они, по-видимому, никогда не существуют вне хромосомы клетки- хозяина; транспозазы, катализирующие их перемещение, действуют на ДНК транспозона в то время, как он пребывает в интегрированном состоянии в геноме хозяина. Транспозаза, как полагают, связывается с короткими нуклеотидными последовательностями, повторяющимися в инвертированном виде на обоих концах транспозона, и тем самым сближает эти концы для последующей рекомбинации, которую она катализирует (рис. 5-76). У некоторых транспозонов механизм транспозиции сводится всего лишь к разрыву и воссоединению ДНК; два конца транспозона присоединяются при этом к концам хромосомы в другом ее участке, там, где в ней возник ступенчатый разрыв (см. рис. 5-67, Б). Такие транспозоны переходят непосредственно из одного участка хромосомы в другой без сопутствующей репликации ДНК. Однако при сшивании хромосомы в том месте, откуда был удален транспозон, ее нуклеотидная последовательность нередко нарушается, т. е. в этом участке хромосомы возникает мутация.
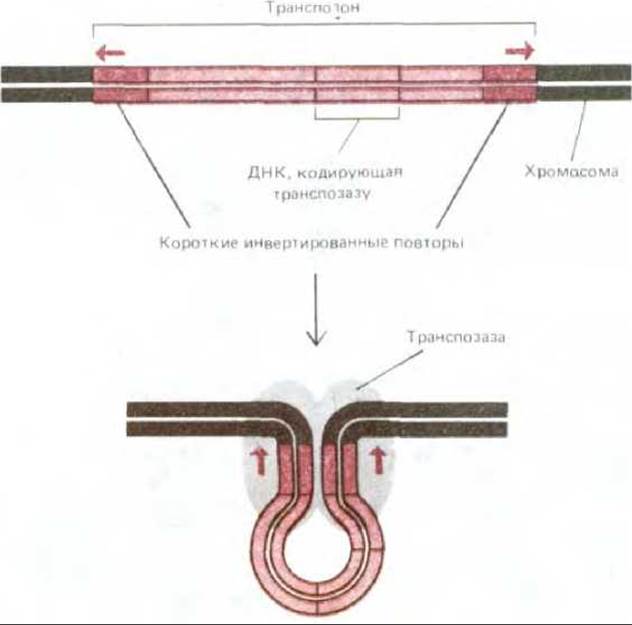
Рис. 5-76. Структура транспозона, перемещающегося непосредственно из одного участка хромосомы в другой. Узнаются транспозоны этого типа по наличию у них на концах двух инвертированных повторяющихся последовательностей ДНК. Как показали эксперименты, именно эти повторяющиеся последовательности (длиной иногда не более 20 нуклеотидов) требуются для того, чтобы находящаяся между ними ДНК могла быть перенесена на новое место специальным ферментом транспозазой. На схеме показан белковый комплекс, образующийся на первом этапе процесса транспозиции. Дальнейшие события включают разрыв ДНК на концах инвертированных повторяющихся последовательностей и ряд других этапов, которые можно рассматривать как модификацию схемы, представленной на рис. 5-67, Б (см. также рис. 5-77).
Существуют также транспозоны, при перемещениях реплицирующиеся. Лучше других изучен пример, в котором сайт-специфическая рекомбинация «запускает» локальный синтез ДНК; одна копия реплицировавшегося транспозона включается при этом в какой-либо новый случайно выбранный участок хромосомы, а другая остается в старом участке (рис. 5-77). Механизм процесса очень близок к тому, который мы только что описали для нерепликативного пути. Известны транспозоны, способные использовать как гот, так и другой путь.
Перемещаясь сами, транспозоны всех типов прихватывают иногда и соседние нуклеотидные последовательности генома хозяина или вызывают в них перестройки. Часто результатом оказываются делеции в соседних участках генома или перенос соседних последовательностей на новое место. Присутствие в хромосомах транспозонов делает их нуклеотидные последовательности менее стабильными, нежели это считалось ранее; возможно, именно эти элементы несут ответственность за многие изменения в геномах, существенные для эволюции.
Можно ли приписать транспозонам еще одну важную эволюционную роль, а именно можно ли считать их древнейшими предками вирусов? Что касается ретровирусов, то они, по-видимому, действительно произошли от ретротранспозонов. Однако нельзя упускать из виду, что все ныне существующие транспозоны теснейшим образом зависят от метаболизма ДНК, у первых же клеток геномы состояли, как полагают, не из ДНК, а из РНК. Очевидно поэтому, путь к самым истокам происхождения вирусов следует искать именно в метаболизме РНК.
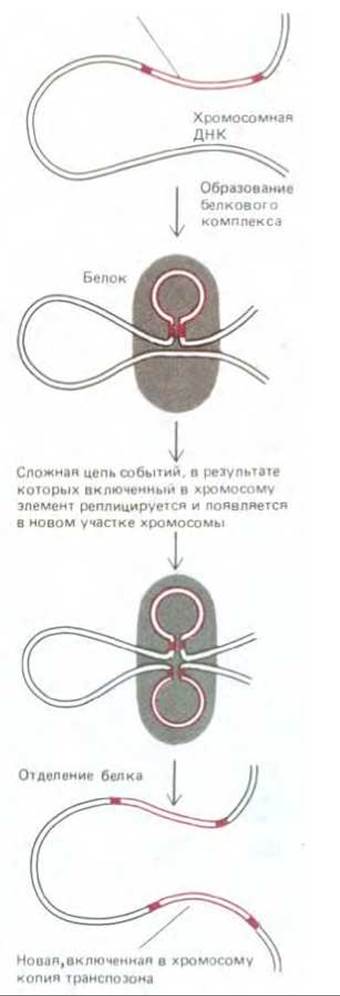
Рис. 5-77. Схема, иллюстрирующая перемещение в хромосоме одного из типов транспозонов. Транспозон этого типа реплицируется в процессе транспозиции, так что, появляясь в новом участке хромосомы, он в то же время в виде одной из копий остается и в старом участке. Две инвертированные повторяющиеся последовательности ДНК на концах транспозона представлены здесь красными прямоугольниками. В начале процесса транспозиции транспозаза разрывает одну из двух цепей ДНК на обоих концах транспозона, чтобы мог начаться синтез ДНК, для которого транспозон служит матрицей. Синтез идет путем добавления нуклеотидов к 3'-концам нуклеотидных последовательностей ДНК хромосомы.
Известны и последующие этапы этого процесса, но весь он в целом слишком сложен для того, чтобы рассматривать его здесь.
5.5.11. Большинство вирусов, вероятно, возникло в процессе эволюции из плазмид [53]
Даже самые крупные вирусы не способны осуществлять биосинтетические процессы самостоятельно; они зависят в этом отношении от клеток-хозяев. Среди известных нам вирусов ни один не имеет собственных рибосом и не обладает способностью синтезировать АТР, в котором он нуждается. Ясно поэтому, что клетки должны были появиться в процессе эволюции раньше вирусов. Предшественниками вирусов были, вероятно, небольшие фрагменты нуклеиновых кислот, которые приобрели способность размножаться независимо от хромосом их клеток-хозяев. Такие независимо размножающиеся элементы - их назвали плазмидами - реплицируются самостоятельно. Плазмиды встречаются как в ДНК-, так и в РНК-форме, и так же как вирусы, они содержат особую нуклеотидную последовательность, которая служит точкой начала репликации. Однако в отличие от вирусов они не способны синтезировать белок для построения белковой оболочки и, не имея такой оболочки, не могут свободно переходить из клетки в клетку. Многие из них не способны также включаться в хромосому клетки-хозяина.
Возможно, что первые РНК-плазмиды напоминали собой вироиды, встречающиеся в некоторых растительных клетках. Эти небольшие кольцевые молекулы РНК (не более 300-400 нуклеотидов) размножаются, хотя они и не кодируют никаких белков (см. рис. 10-61). Не имея капсида, вироиды существуют лишь как голые молекулы РНК и переходят от растения к растению только в том случае, когда и донорная клетка, и клетка-реципиент оказываются поврежденными, т. е. когда между ними не существует мембранного барьера, который вироид не способен преодолеть. Под давлением естественного отбора такие независимо реплицирующиеся элементы могли, очевидно, включать в себя те нуклеотидные последовательности клетки-хозяина, которые облегчали их самостоятельное размножение, в том числе и некоторые последовательности, кодирующие белки. Некоторые известные нам плазмиды действительно достаточно сложны: в них закодированы белки и молекулы РНК, регулирующие их размножение, а также белки, регулирующие их распределение между дочерними клетками. Самые крупные среди известных плазмид представляют собой кольцевые двухцепочечные спирали ДНК, насчитывающие свыше 100000 пар нуклеотидов.
Первый вирус возник, вероятно, когда у плазмиды появился ген, кодирующий белок капсида. Однако в капсиде умещается ограниченное количество нуклеиновой кислоты, и, значит, число генов, которые вирус может в себе заключать, лимитируется размерами капсида. Будучи вынуждены оптимальным образом использовать свой небольшой геном, некоторые мелкие вирусы (вроде бактериофага ФХІ74) обзавелись в ходе эволюции перекрывающимися генами, в которых часть нуклеотидной последовательности, кодирующей один какой-нибудь белок, используется (с той же или с иной рамкой считывания) для кодирования второго белка. У других вирусов в процессе эволюции возникли более крупные капсиды, что должно было дать им возможность приобретать новые полезные гены.
Обладая уникальной способностью к переносу ДНК через видовые барьеры, вирусы, почти несомненно, играли важную роль в эволюции тех организмов, которые они инфицируют. Многие вирусы часто вступают в рекомбинацию как друг с другом, так и с хромосомами клеток-хозяев; при этом они захватывают случайные фрагменты хромосом и переносят их в другие клетки или другие организмы. Кроме того, включившиеся в геном хозяина (интегрированные) копии вирусной ДНК (провирусы) становятся постоянными компонентами генома у большей части организмов. Примеры таких провирусов мы находим в семействе бактериофагов X и среди так называемых эндогенных ретровирусов, многочисленные копии которых обнаруживаются в геномах позвоночных. Интегрированная вирусная ДНК часто видоизменяется так, что способность образовывать полноценный вирус у нее утрачивается, но она все еще может кодировать белки, причем некоторые из них оказываются полезными для клетки. Вирусы, следовательно, так же, как и половой процесс, создают возможности для ускорения эволюции, открывая для нее такой путь, как смешение генофондов различных организмов.
С помощью вирусов отдельные последовательности ДНК переносятся из генома в геном (феномен трансдукции). Таким образом, вирусы можно использовать как инструмент для переноса генов из одной клетки в другую. Вирусы, а также очень близкие к ним плазмиды и транспозоны играют важную роль в биологии клетки и по другим причинам. Благодаря тому что они относительно просто устроены, изучение процесса их размножения продвигается чрезвычайно быстро, а полученные при этом данные позволяют судить об основных генетических механизмах клеток. Кроме того, вирусы и плазмиды стали главными элементами в технологии рекомбинантной ДНК. К рассмотрению этой последней темы мы теперь и перейдем.
Заключение
Вирусы — это инфекционные частицы, которые состоят из молекул ДНК или РНК (они образуют геном вируса), упакованных в белковый капсид; у некоторых вирусов капсид окружен еще и мембранной оболочкой, основу которой составляет липидный бислой. Строение вирусного генома и способы его репликации у разных вирусов сильно варьируют. Вирус способен размножаться только в клетке-хозяине, используя для этого ее генетические механизмы. Обычно вирусная инфекция завершается лизисом инфицированной клетки и высвобождением потомства вируса. Однако некоторые вирусы могут включаться в хромосому клетки, не вызывая лизиса последней. Здесь вирусные гены (в форме провируса) реплицируются вместе с генами хозяина. Считается, что многие вирусы возникли в ходе эволюции из плазмид, которые представляют собой самореплицирующиеся молекулы ДНК или РНК, не способные окружать себя белковой оболочкой.
Транспозоны - это нуклеотидные последовательности ДНК, отличающиеся от вирусов тем, что размножаясь только в клетках-хозяевах или в их потомстве, они, подобно плазмидам, не могут покинуть клетку. От плазмид же транспозоны отличаются тем, что они обычно реплицируются лишь в составе хромосомы хозяина, как ее неотъемлемая часть. Некоторые транспозоны, однако, весьма напоминают ретровирусы: они перемещаются в новые участки генома в результате обратной транскрипции промежуточного РНК-соединения. Существуют и транспозоны, способные оказаться в новом участке хромосомы, сохранив свою копию и на прежнем месте. Хотя и вирусы, и транспозоны могут рассматриваться как паразиты, они полезны тем, что вызываемые ими перестройки нуклеотидных последовательностей ДНК нередко играют важную роль в эволюции клеток и организмов.