Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Введение в биологию клетки
Эволюция клетки
От прокариот - к эукариотам
Существует предположение, что все ныне живущие организмы произошли из единственной, возникшей несколько миллиардов лет назад первобытной клетки. Пережив своих конкурентов, эта клетка положила начало процессу клеточного деления и эволюции, который в конце концов создал зеленый покров Земли, изменил состав ее атмосферы и сделал ее родиной разумной жизни. Видимо, только так можно объяснить «фамильное сходство» между всеми организмами. На эволюционном пути имеется важная веха. Приблизительно 1,5 млрд. лет назад произошел переход от маленьких клеток со сравнительно простой внутренней структурой (так называемых прокариот, к которым относятся различные бактерии) к большим по размеру и значительно более сложно устроенным эукариотическим клеткам, подобным клеткам высших животных и растений.
1.2.1. Прокариотические клетки имеют простую структуру, но различаются по биохимическим свойствам [7]
Бактерии - наиболее простые организмы, обнаруженные в большинстве природных сред обитания. Это - сферические или удлиненные клетки обычно размером в несколько микрометров (рис. 1-13). Как правило, у них имеется жесткая защитная оболочка, называемая клеточной стенкой, под которой находится плазматическая мембрана, ограничивающая единственный цитоплазматический компартмент, содержащий ДНК, РНК, белки и малые молекулы. В электронном микроскопе содержимое таких клеток имеет вид матрикса различной плотности без явно выраженных организованных внутренних структур (см. рис. 1-8, А).
Бактерии малы и способны быстро размножаться путем простого бинарного деления. При избытке питательных веществ «выживание наиболее приспособленных» обычно означает выживание тех, которые быстрее всех делятся. В оптимальных условиях прокариотическая клетка может делиться каждые 20 минут и, таким образом, образовать до 5 млрд. клеток (что приблизительно равно населению земного шара) менее, чем за 11 часов. Благодаря способности быстро делиться бактериальные популяции с легкостью адаптируются к изменениям окружающей среды. Например, в лабораторных условиях популяция бактерий, поддерживаемая в большом сосуде, за несколько недель благодаря спонтанным мутациям и естественному отбору приобретает способность использовать в качестве источника углерода новые типы Сахаров.
В природе бактерии занимают невообразимое множество экологических ниш, и столь же многообразным оказывается их биохимическое строение. Различают две группы бактерий: эубактерии - часто встречающиеся формы, населяющие почву, воду и другие организмы, и архебактерии, встречающиеся в таких неудобных средах обитания, как болота, океанские глубины, очень соленые воды и горячие кислые источники (рис. 1-14).

Рис. 1-13. Некоторые прокариотические клетки, изображенные в одинаковом масштабе.

Рис. 1-14. Родственные связи между современными бактериями (стрелками показаны вероятные пути эволюции). Происхождение эукариотических клеток обсуждается дальше в тексте.
Существуют виды бактерий, способные питаться практически любыми органическими молекулами - сахарами, аминокислотами, жирами, углеводами, полипептидами и полисахаридами. Некоторые даже могут получать атомы углерода из СО2 и атомы азота из N2. Несмотря на относительно простое строение, бактерии живут на Земле дольше любых других организмов и превосходят по численности все другие типы клеток.
1.2.2. Развитие метаболических реакций
Бактерии, которая растет в растворе солей, содержащем в качестве единственного источника углерода глюкозу, необходимо осуществлять множество химических реакций. Она должна не только извлечь из глюкозы химическую энергию, необходимую для многих жизненно важных процессов, но и использовать атомы углерода для синтеза всех необходимых клетке органических молекул. Эти реакции катализируются сотнями ферментов, последовательно работающих в «цепях» химических реакций, так что продукт одной реакции служит субстратом для следующей. Такие цепи ферментативных реакций, называемые метаболическими путями, обсуждаются в следующей главе.
Вначале, когда жизнь на Земле только зародилась, в метаболических реакциях, видимо, не было необходимости: клетки могли жить и расти, питаясь окружающими их молекулами - наследием первобытного бульона. По мере истощения этих естественных ресурсов большое преимущество при отборе должны были получить организмы, вырабатывающие ферменты для образования органических молекул. Считается, что таким образом наличный комплект клеточных ферментов постепенно увеличивался, и в результате возникли метаболические пути современных организмов. Два возможных варианта эволюции метаболических путей проиллюстрированы на рис. 1-15.
Если эволюция метаболических путей шла путем последовательного добавления новых ферментативных реакций к существовавшим ранее, то, подобно самым старым годовым кольцам на срезе ствола, наиболее древние реакции должны находиться ближе всего к центру «метаболического древа» - там, где синтезируются наиболее существенные молекулярные «кирпичики». Такое центральное положение в метаболизме прочно занимают реакции с участием фосфатов Сахаров, в самом центре которых, видимо, находится последовательность реакций, называемая гликолизом и способная осуществлять расщепление глюкозы в отсутствие кислорода (т. е. анаэробно). Самые древние из метаболических путей должны были быть анаэробными, поскольку в атмосфере первобытной Земли кислорода не было. Практически во всех живых клетках протекают реакции гликолиза, сопровождающиеся образованием аденозинтрифосфата, или АТР, - соединения, используемого всеми клетками в качестве источника легкодоступной химической энергии.

Рис. 1-15. Схема, иллюстрирующая два возможных механизма возникновения метаболических путей. Клетка слева имеет в своем распоряжении запасы родственных соединений (А, Б, В, Г), возникшие благодаря пребиотическому синтезу. Одно из соединений (Г) метаболически полезно. По мере истощения запасов этого соединения преимущество при отборе получают те клетки, которые «учатся» синтезировать новый фермент, катализирующий образование Г из В. Последовательное повторение подобных этапов могло бы привести к развитию важных метаболических путей. В клетке, показанной справа, в изобилии имеется метаболически полезное соединение А. В процессе эволюции возникает фермент, который случайно оказывается способным превращать соединение А в соединение Б. Затем клетка претерпевает другие изменения, позволяющие ей использовать это новое вещество. Дальнейшее появление новых ферментов может в конце концов привести к возникновению длинной цепи реакций.
С находящимися в центре обмена веществ превращениями фосфатов Сахаров связаны сотни других химических реакций. Часть из них отвечает за синтез малых молекул, многие из которых в свою очередь используются в дальнейших реакциях синтеза больших, специфических для конкретного организма полимеров. Другие реакции участвуют в расщеплении сложных молекул пищи до более простых химических соединений. Одна из наиболее поразительных особенностей всех этих реакций - это то, что они протекают в любых типах организмов. Конечно, существуют и отличия: многие специфические продукты метаболизма присущи только некоторым родам или видам. Например, общая для всех аминокислота лизин синтезируется различными путями у бактерий, дрожжей и зеленых растений, и совсем не синтезируется у высших животных. Тем не менее в широком смысле подавляющее большинство реакций и катализирующих их ферментов характерно для всех живых существ - от бактерий до человека. Поэтому считается, что примитивные исходные клетки, породившие все живое, уже выполняли эти реакции. Ферменты, катализирующие основные метаболические реакции, по мере дивергенции организмов постепенно модифицировались, не изменяя при этом своей основной функции. Поэтому аминокислотные последовательности одного и того же типа фермента в различных современных организмах, содержат исключительно ценную информацию об эволюционном родстве этих видов. Полученные таким путем данные хорошо согласуются с результатами других исследований, например с изучением ископаемых остатков. Еще более богатый источник информации заключен в нуклеотидных последовательностях ДНК современных клеток. Сравнение высококонсервативных последовательностей, которые детерминируют основные функции (и, следовательно, очень медленно меняются в ходе эволюции), позволяет судить о родстве давно дивергировавших организмов (рис. 1-16). Более быстро эволюционировавшие последовательности могут использоваться для оценки эволюционных процессов у родственных видов (рис. 1-17). Можно надеяться, что применение этих методов позволит с беспрецедентной точностью проследить ход эволюционного процесса.

Рис. 1-16. Эволюционное родство организмов по данным секвенирования (определения нуклеотидных последовательностей) генов рибосомной РНК малой субъединицы. Эти гены содержат высококонсервативные последовательности, меняющиеся столь медленно, что их можно использовать для «измерения» филогенетического родства между представителями всех групп живых организмов. Из этих данных следует, что в ходе эволюции эукариотических клеток линии растений, животных и грибов дивергировали от общего предка сравнительно поздно. (По М. L. Sogin, H.J. Elwood, J. H. Gunderson, Proc. Natl. Acad. Sci. USA, 83, 1383-1387, 1986.)
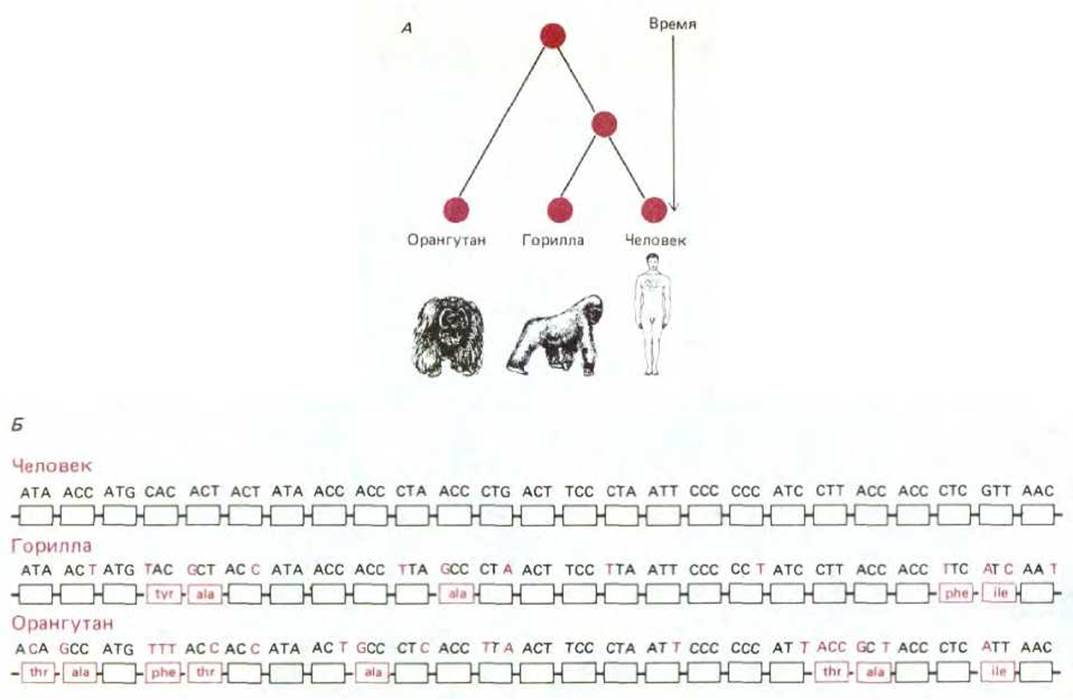
Рис. 1-17. К кому ближе человек, к горилле или орангутану? Ответ на этот вопрос можно получить, проанализировав последовательности ДНК этих видов и построив родословную, представленную в верхней части рисунка. Для выяснения филогенетического родства между близкородственными организмами обычно используют митохондриальную ДНК, поскольку она эволюционирует примерно в 5-10 раз быстрее, чем ядерная ДНК. В нижней части рисунка приведена последовательность первых 75 нуклеотидов одного и того же гена (митохондриальный ген, кодирующий субъединицу NAD-дегидрогеназы) для каждого из видов. Цветными буквами отмечены участки, по которым горилла и орангутан отличаются от человека. Прямоугольники под нуклеотидными последовательностями обозначают аминокислоты в соответствующих белках. Названия аминокислот, если они отличаются от соответствующих аминокислот человека, также выделены цветом. Проведенный анализ показывает, что последовательность гориллы отличается от последовательности человека на 10%, а орангутана - на 17%. Предполагая, что эти различия обусловлены случайными мутациями, появляющимися с одинаковой частотой в линии каждого вида, можно вывести родословную, показанную на схеме А. (Данные из W. M. Brown, Е. М. Prager, A. Wang, А. С. Wilson, J. Mol. Biol., 18; 225-239, 1982.)
1.2.3 Цианобактерии способны фиксировать СО2 и N2
Самые ранние стадии метаболизма появились в ходе эволюции для того, чтобы восполнить недостачу органических молекул, образовавшихся еще в пребиотических условиях. Что же произошло, когда запас подобных соединений полностью истощился? В этих условиях значительное преимущество при отборе должны были приобрести организмы, способные использовать атомы углерода и азота (в виде СО2 и N2) из атмосферы. Но углекислый газ и молекулярный азот хотя и имеются в изобилии, очень стабильны. Их превращение в доступную для усвоения форму (органических молекул, таких, как простые сахара) требует большого количества энергии и значительного числа сложных химических реакций.
Для использования СО2 в ходе эволюции возник механизм фотосинтеза, в процессе которого СО2 превращается в органическое соединение за счет энергии солнечного излучения. Взаимодействие солнечного света с пигментной молекулой - хлорофиллом - переводит электрон в более высокое энергетическое состояние. Энергия, высвобождаемая при обратном переходе электрона на более низкий энергетический уровень, направляется молекулами белка на проведение химических реакций.
По всей вероятности, одной из первых реакций с использованием солнечного света было фосфорилирование нуклеотидов с образованием АТР - богатого энергией соединения. Другой важный этап в ходе эволюции - создание «восстановительной силы» (восстановительных эквивалентов). Дело в том, что атомы азота и углерода в СО2 и N2 атмосферы находятся в инертном окисленном состоянии, и один из путей сделать их более реакционноспособными, с тем чтобы они могли участвовать в биосинтезе, - это восстановить их, т.е. передать им электроны. Восстановление идет следующим образом. Хлорофилл, используя энергию солнечного света, отбирает электроны у слабых доноров электронов и переносит их на сильные доноры электронов, которые в свою очередь используются для восстановления СО2 и N2. Анализ механизмов фотосинтеза у современных бактерий позволяет сделать вывод о том, что одним из первых источников электронов был H2S, конечным продуктом обмена (метаболическим отходом) которого должна была быть элементарная сера. Значительно позже развился куда более сложный, но, как оказалось, более полезный процесс «извлечения» электронов из Н2О. В результате в качестве отхода в земной атмосфере начал накапливаться кислород.
В современном мире основной путь, по которому углерод и азот включаются в органические молекулы и поступают в биосферу, реализуется в процессе жизнедеятельности цианобактерий (называемых также синезелеными водорослями). К этой группе относятся наиболее автономные из ныне живущих организмов. Они способны «фиксировать» СО2 и N2 и существовать только за счет воды, воздуха и солнечного света, причем механизмы, с помощью которых это достигается, в своих общих чертах, по-видимому, не изменились более чем за 1 млрд. лет. Вместе с другими бактериями, обладающими некоторыми из этих способностей, цианобактерий создали условия, в которых могли развиваться более сложные типы организмов: как только одна группа организмов преуспела в синтезе всего диапазона органических компонентов клетки из неорганических веществ, другие организмы получили возможность существовать, питаясь первичными продуцентами и продуктами их жизнедеятельности.
1.2.4. Бактерии могут осуществлять аэробное окисление молекул пищи
Многие сегодня справедливо обеспокоены влиянием человеческой деятельности на окружающую среду. Однако в прошлом другие организмы, хотя и значительно медленнее, тоже вызывали радикальные изменения условий на Земле. Лучше всего это видно на примере состава земной атмосферы, которая с появлением фотосинтеза превратилась из практически лишенной молекулярного кислорода смеси газов в смесь, в которой содержание кислорода составляет 21%.
Учитывая высокую химическую активность кислорода и его способность реагировать с большинством компонентов цитоплазмы, можно сделать вывод, что для многих ранних организмов кислород, видимо, был токсичен (как и для многих современных анаэробных бактерий). Однако именно благодаря высокой реакционной способности кислород способен выступать в роли «поставщика» химической энергии, и не удивительно, что в ходе эволюции организмы использовали это свойство. С помощью кислорода живые существа способны более полно окислять молекулы пищи. Например, в отсутствие кислорода глюкоза может быть расщеплена только до молочной кислоты или этилового спирта, конечных продуктов анаэробного гликолиза. В присутствии же кислорода глюкоза полностью расщепляется до Н2О и СО2. Таким способом можно получить значительно больше энергии из каждого грамма глюкозы. Энергия, высвобождаемая при аэробном окислении молекул пищи, называемом обычно дыханием, используется для синтеза АТР, подобно тому как у фотосинтезирующих организмов АТР образуется за счет солнечной энергии. В обоих случаях происходит ряд последовательных реакций переноса электронов, которые создают разность концентраций ионов Н+ внутри и снаружи небольших ограниченных мембранами компартментов. Полученный таким образом градиент концентрации Н+ служит источником энергии для синтеза АТР. На сегодняшний день дыхание характерно для подавляющего большинства организмов, включая и большинство прокариот.
1.2.5. Клетки эукариот содержат несколько характерных органелл
Как же повлияло накопление молекулярного кислорода в атмосфере на анаэробные организмы, положившие начало жизни на Земле? В мире, богатом кислородом, который не мог быть ими использован, такие организмы оказались в невыгодных условиях. Некоторые из них, без сомнения, вымерли. Другие либо развили способность к дыханию, либо нашли экологические ниши, практически лишенные кислорода, и продолжили в них анаэробное существование. По всей вероятности, существовал и третий класс организмов, который выбрал значительно более хитрую и неизмеримо более богатую отдаленными последствиями стратегию выживания. Считается, что организмы этого класса вступили в симбиоз с аэробными клетками, а затем образовали с ними прочную ассоциацию. Это - наиболее привлекательное объяснение возникновения современных клеток эукариотического типа (схема 1-1), о которых в основном и пойдет речь в книге.
Эукариотические клетки по определению и в отличие от прокариотических имеют ядро (по-гречески «карион»). Ядро, в котором находится большая часть клеточной ДНК, ограничено двойной мембраной (рис. 1-18). Таким образом, компартмент, содержащий ДНК, отделен от остального содержимого клетки - цитоплазмы, где протекает большинство метаболических реакций. В самой цитоплазме различают множество характерных органелл. Среди них особенно выделяются два типа-митохондрии и хлоропласти (рис. 1-19 и 1-20). Каждая из этих органелл окружена собственной двойной мембраной, отличающейся по химическим свойствам от мембраны, окружающей ядро. Митохондрии - почти универсальный компонент эукариотических клеток, тогда как хлоропласты встречаются лишь в тех эукариотических клетках, которые способны к фотосинтезу, т. е. в клетках растений, но не животных и грибов. Считается, что обе органеллы имеют симбиотическое происхождение.
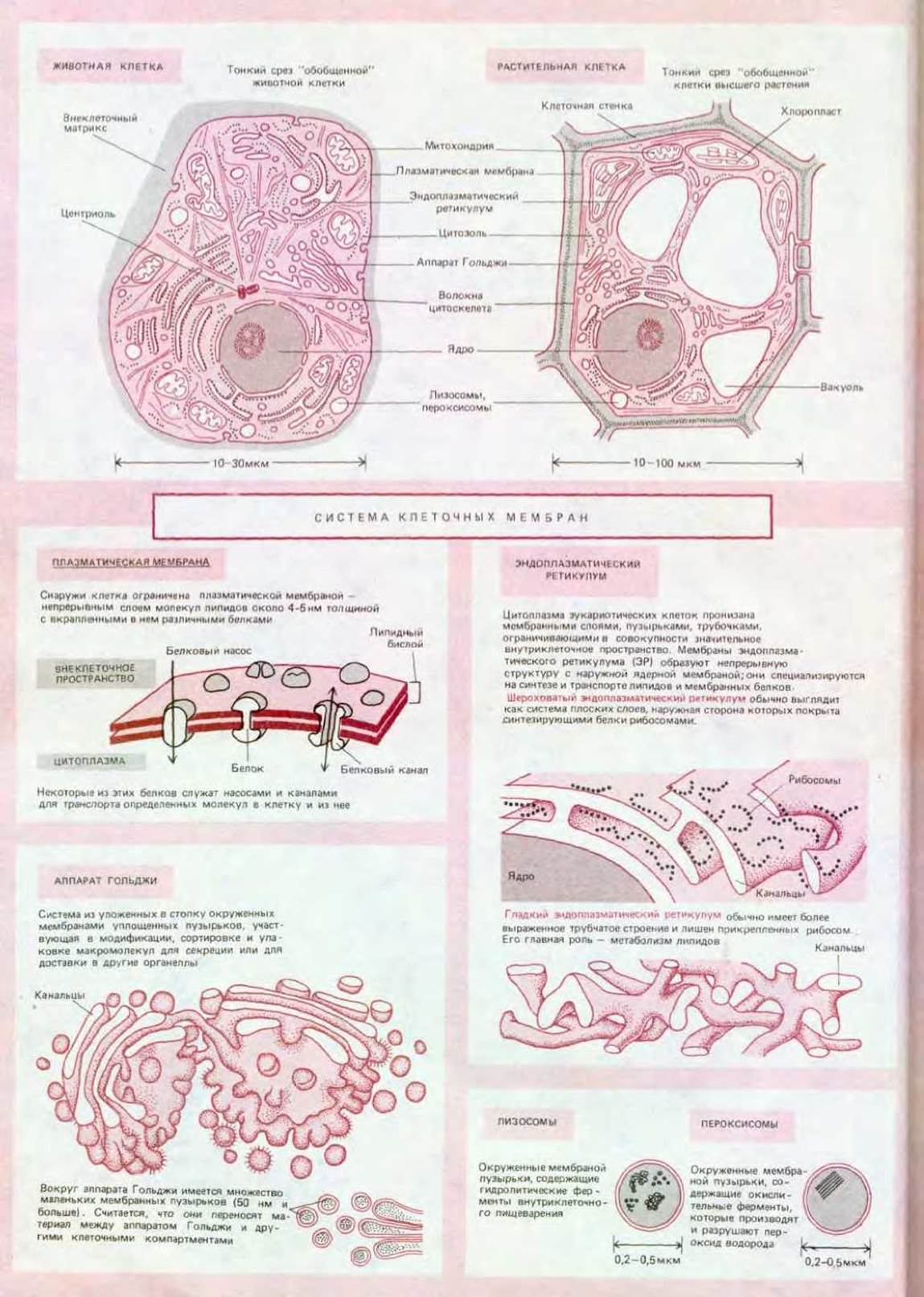
Схема 1-1. Основные органеллы эукариотических клеток.
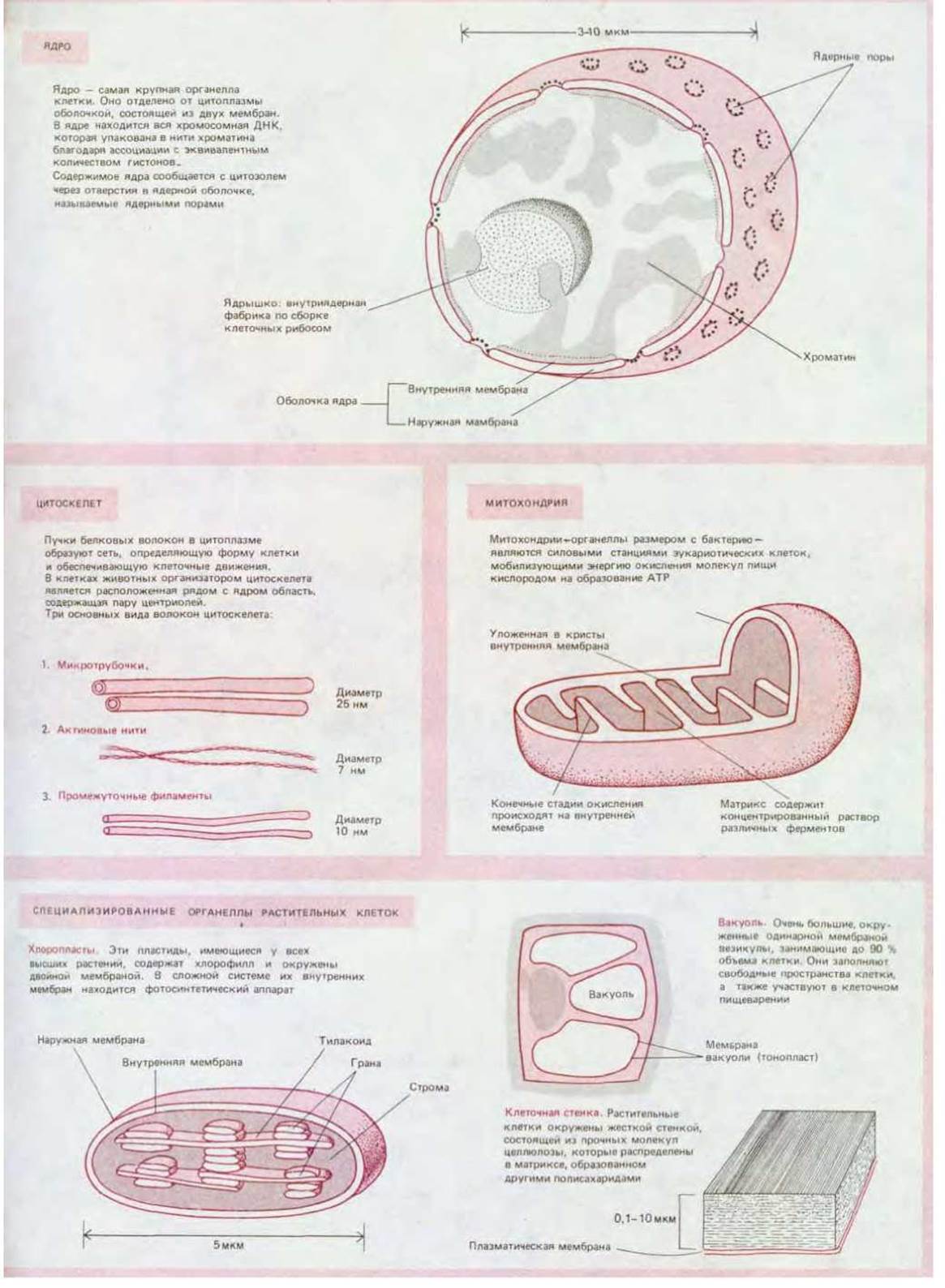

Рис. 1-18. В ядре содержится большая часть ДНК эукариотической клетки. Ядро отчетливо видно на электронной микрофотографии тонкого среза клетки млекопитающего. (С любезного разрешения Daniel S. Friend.)
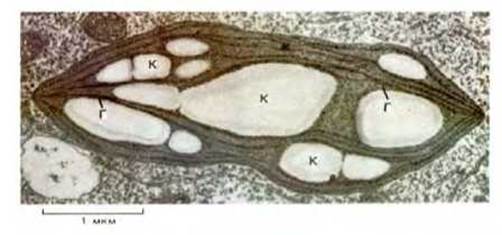
Рис. 1-19. Электронная микрофотография хлоропласта клетки мха. Видна разветвленная система внутренних мембран. Уплощенные мембранные мешочки, содержащие хлорофилл, организованы в стопки, или граны (Г). Этот хлоропласт содержит также большие скопления крахмала (К). (С любезного разрешения J. Burgess.)

Рис. 1-20. Митохондрии осуществляют окислительное расщепление питательных веществ во всех эукариотических клетках. Как видно на этой электронной микрофотографии, их наружная мембрана гладкая, а внутренняя очень извилистая. (С любезного разрешения Daniel S. Friend.)
1.2.6. Эукариотические клетки зависят от митохондрий, осуществляющих окислительный метаболизм
Митохондрии во многом похожи на свободноживущие прокариотические организмы: например, они напоминают бактерий по форме и размеру, содержат ДНК, производят белок и размножаются делением. Разрушив эукариотические клетки и разделив их компоненты, можно показать, что митохондрии ответственны за дыхание и что ни в каких других частях клетки этот процесс не происходит. Без митохондрий клетки животных и грибов были бы анаэробами, зависимыми в своих энергетических потребностях от сравнительно малоэффективного и архаичного процесса гликолиза. Многие современные бактерии могут дышать, причем механизм этого дыхания имеет явное сходство с дыханием у митохондрий, и есть все основания думать, что эукариотические клетки являются потомками примитивных анаэробных организмов, которые выжили в богатом кислородом мире, поглотив аэробных бактерий. Они поддерживали их в состоянии симбиоза ради присущей им способности потреблять атмосферный кислород и производить энергию. Анализ некоторых современных организмов свидетельствует о возможности такого эволюционного события. Существует несколько сотен видов одноклеточных эукариот, которые напоминают гипотетический предковый эукариотический организм тем, что живут в условиях дефицита кислорода (например, в кишечнике животных) и совсем не имеют митохондрий. Сравнение нуклеотидных последовательностей, проведенное в последнее время, дает основание предполагать, что одна группа таких организмов, микроспоридии, очень рано в эволюции ответвилась от линии, ведущей к другим эукариотическим клеткам. Другой представитель эукариот, амеба Pelomyxa palustris, хотя и лишен митохондрий, все же осуществляет окислительный метаболизм, «приютив» в своей цитоплазме аэробных бактерий и установив с ними постоянные симбиотические отношения. Таким образом, существование микроспоридий и Pelomyxa свидетельствует о реальности двух этапов в эволюции эукариот, к которым относимся и мы сами.
Приобретение митохондрий должно было иметь много последствий. Например, у прокариотических клеток плазматическая мембрана тесно связана с образованием энергии, в то время как у эукариотических клеток эта важнейшая функция передана митохондриям. Кажется вполне вероятным, что освобождение плазматической мембраны эукариотической клетки от этой функции позволило ей приобрести новые свойства. В частности, поскольку эукариотическим клеткам не нужно поддерживать высокий градиент Н+ на своей мембране (что необходимо для производства АТР у прокариот), у них появляется возможность использовать контролируемые изменения в ионной проницаемости плазматической мембраны в целях клеточной сигнализации. Следовательно, одновременно с возникновением эукариот в плазматической мембране появляются ионные каналы. В настоящее время у высших организмов эти каналы опосредуют сложные процессы передачи электрических сигналов (в частности, в нервной системе), а у одноклеточных свободноживущих эукариот, таких, как простейшие, они во многом определяют их поведение (см. ниже).
1.2.7. Хлоропласты представляют собой потомков «захваченных» прокариотических клеток
Хлоропласты осуществляют фотосинтез в значительной степени так же, как прокариоты-цианобактерии, солнечный свет у них поглощается присоединенным к мембранам хлорофиллом. Некоторые Хлоропласты по строению во многом напоминают цианобактерии, например сходными могут быть их размеры и способ укладки в слои хлорофиллсодержащих мембран (рис. 1-20). Показано также, что хлоропласты размножаются делением, а нуклеотидная последовательность их ДНК почти полностью гомологична определенным участкам бактериальной хромосомы. Все это наводит на мысль, что хлоропласты и цианобактерии имеют общего предка и что хлоропласты произошли от прокариот, «захваченных» когда-то эукариотическими клетками. Прокариоты осуществляли фотосинтез для клеток-хозяев в обмен на предоставляемые последними «приют» и питание. Симбиоз фотосинтезирующих клеток с другими типами клеток - явление достаточно частое, и ряд современных эукариотических клеток содержат в себе истинные цианобактерии (рис. 1-21).
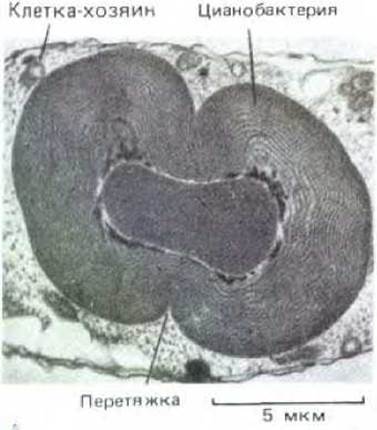
Рис. 1-21. Организм, близкородственный современным цианобактериям, но являющийся облигатным симбионтом другой клетки (вместе эти два организма известны как Cyanophora paradoxa). Показан момент деления «цианобактерии». (С любезного разрешения Jeremy D. Pickett-Heaps.)
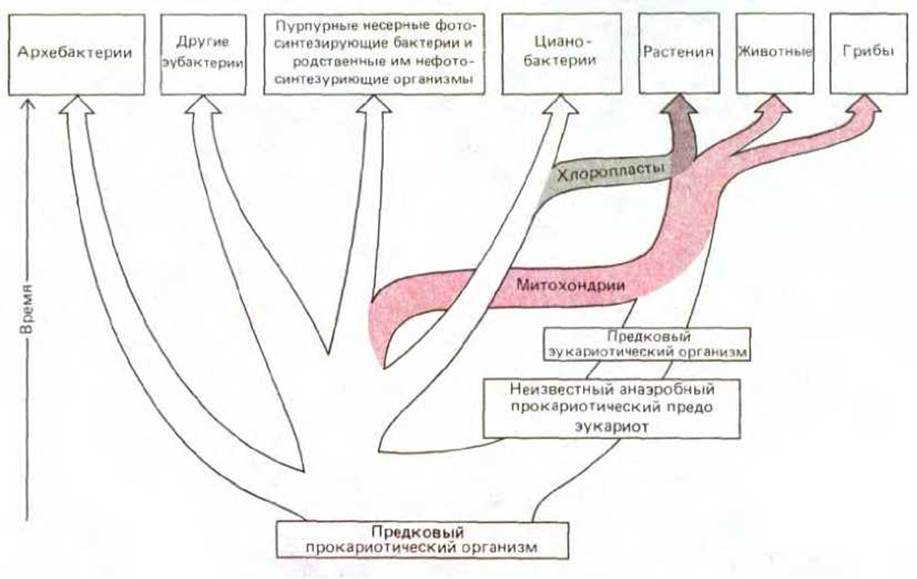
Рис. 1-22. Гипотеза о происхождении эукариот путем симбиоза аэробных и анаэробных прокариот
Таблица 1-1. Сравнение прокариотических и эукариотических организмов
|
Прокариоты |
Эукариоты |
|
|
Организмы |
Бактерии и цианобактерии |
Протисты, грибы, растения и животные |
|
Размер клеток |
Обычный линейный размер 1-10 мкм |
Обычный линейный размер 10 100 мкм |
|
Метаболизм |
Анаэробный или аэробный |
Аэробный |
|
Органеллы |
Немногочисленны или отсутствуют |
Ядро, митохондрии, хлоропласты, эндоплазматический ретикулум и др. |
|
ДНК |
Кольцевая ДНК в цитоплазме |
Очень длинная ДНК с большим количеством некодирующих участков организована в хромосомы и окружена ядерной мембраной |
|
РНК и белки |
РНК и белки синтезируются в одном компартменте |
Синтез и процессинг РНК происходят в ядре, синтез белков в цитоплазме |
|
Цитоплазма |
Отсутствие цитоскелета, движения цитоплазмы, эндо- и экзоцитоза |
Имеются цитоскелет из белковых волокон, движение цитоплазмы, эндоцитоз и экзоцитоз |
|
Деление клеток |
Бинарное деление |
Митоз (или мейоз) |
|
Клеточная организация |
Преимущественно одноклеточные |
Преимущественно многоклеточные с клеточной дифференцировкой |
На рис. 1-22 показано эволюционное происхождение эукариот в соответствии с симбиотической теорией. Следует отметить, что митохондрии и хлоропласты, проявляя определенное сходство с современными аэробными бактериями и цианобактериями, в то же время во многих отношениях отличаются от них. Например, количество ДНК в этих органеллах очень мало, большинство составляющих их молекул синтезируется вне органелл и лишь затем в них транспортируется. Если считать, что митохондрии и хлоропласты действительно возникли из симбиотических бактерий, то следует признать, что они претерпели значительные эволюционные изменения и стали весьма зависимыми от своих хозяев.
Для современных эукариот характерно не только наличие митохондрий, им присущ целый ряд особенностей, отличающих их от прокариот (табл. 1-1). Вместе все эти особенности наделяют эукариотические клетки большим количеством различных потенциальных возможностей, и трудно сказать, какая из них возникла раньше других. Заметим, что важнейшим шагом на пути эволюции было появление митохондрий в анаэробных эукариотических клетках, поскольку вместе с ними клетки получали эффективный источник энергии и могли направить ее на усложнение своих функций.
1.2.8. Эукариотические клетки содержат множество различных внутренних мембран
Объем эукариотических клеток, как правило, в 1000 и более раз превышает объем клеток прокариот. Соответственно больше в эукариотических клетках и разнообразного клеточного материала, например содержание ДНК в клетках человека в 1000 раз превышает ее количество в клетках бактерий. Известно, что именно на мембране протекает ряд важнейших реакций, связанных с поступлением в клетку сырья для метаболизма и выходом соответствующих продуктов во внеклеточное пространство. Вот почему большой объем эукариотических клеток требует значительного увеличения поверхности их мембраны по сравнению с клетками прокариот. Но согласно законам геометрии, при простом увеличении размеров какого-либо предмета его объем возрастает как куб линейного размера, а площадь поверхности - лишь как квадрат. Поэтому для сохранения необходимого соотношения площади поверхности и объема большие эукариотические клетки вынуждены увеличивать свою поверхность за счет изгибов, складок и других усложнений формы мембраны.
Сложность строения мембран - одна из основных особенностей всех эукариотических клеток. Мембраны окружают ядро, митохондрии и (у растений) хлоропласты. Они образуют лабиринт эндоплазматического ретикулума (рис. 1-23), где синтезируются липиды и мембранные белки, а также материал, предназначенный для экспорта из клетки. Мембраны формируют стопки уплощенных пузырьков, составляющих аппарат Гольджи (рис. 1-24), который тоже участвует в синтезе и транспорте различных органических молекул. Мембраны окружают лизосомы, содержащие запас ферментов, необходимых для внутриклеточного пищеварения, и таким образом защищают от действия этих ферментов белки и нуклеиновые кислоты самой клетки. Точно так же мембраны окружают пероксисомы, где в процессе окисления различных молекул образуются и разлагаются опасные высоко-реакционноспособные перекиси (пероксиды). Мембраны образуют также маленькие везикулы (пузырьки) и большие, заполненные жидкостью вакуоли (у растений). Все эти окруженные мембранами структуры соответствуют определенным компартментам цитоплазмы. В совокупности они занимают почти половину объема типичной живой клетки. Цитоплазматический компартмент, лишенный всех окруженных мембранами органелл, обычно называют цитозолем.

Рис. 1-23. Электронная микрофотография тонкого среза клетки млекопитающего, на которой виден гладкий и шероховатый эндоплазматический ретикулум (ЭР). Участки гладкого ЭР участвуют в липидном обмене, участки шероховатого, усыпанные рибосомами, являются местами синтеза белков. Синтезированные белки покидают цитозоль и входят в некоторые другие компартменты клетки. (С любезного разрешения George Palade.)

Рис. 1-24. Электронная микрофотография среза клетки млекопитающего. Виден аппарат Гольджи. Эта структура состоит из нескольких слоев уплощенных мембранных пузырьков (см. также схему 1-1). Аппарат Гольджи участвует в синтезе и упаковке материала, предназначенного для секреции из клетки, а также в транспорте новосинтезированных белков в отведенный для них клеточный компартмент. (С любезного разрешения Daniel S. Friend.)
Все перечисленные нами мембранные структуры находятся внутри клетки. Как же в таком случае они могут влиять на площадь ее поверхности? Ответ заключается в том, что между внутриклеточными, окруженными мембранами структурами и внеклеточной средой происходит обмен. Он осуществляется с помощью двух уникальных для эукариотических клеток процессов: эндоцитоза и экзоцитоза. При эндоцитозе некоторые участки наружной поверхностной мембраны впячиваются и отрываются, образуя цитоплазматические мембранные пузырьки, содержащие вещества, которые находились во внешней среде или были адсорбированы на поверхности клетки. Таким способом в цитоплазму могут попасть весьма крупные частицы и даже целые клетки (фагоцитоз). Экзоцитоз - обратный процесс, при котором внутриклеточные мембранные пузырьки сливаются с плазматической мембраной, высвобождая тем самым свое содержимое во внешнюю среду. Благодаря описанному механизму, расположенные глубоко внутри клетки и окруженные мембранами компартменты увеличивают эффективную поверхность клетки, участвующую в обмене веществ с внешней средой.
Как мы увидим в дальнейших главах, различные мембраны и окруженные ими компартменты эукариотических клеток стали высокоспециализированными: одни предназначены для секреции, другие - для всасывания, третьи - для определенных биосинтетических процессов и т. д.
1.2.9. Эукариотические клетки имеют скелет
Чем больше клетка, чем сложнее и специализированнее ее внутренние структуры, тем больше необходимость контролировать положение и перемещение этих структур. Все эукариотические клетки имеют внутренний скелет цитоскелет, определяющий форму клеток, их способность двигаться и перемещать органеллы из одной части клетки в другую. Цитоскелет образован сетью белковых волокон. Наиболее важные среди них - это актиновые нити и микротрубочки (рис. 1-25), которые, очевидно, возникли на очень ранних этапах эволюции, так как встречаются у всех эукариот практически в неизменном виде. И те, и другие участвуют в механизмах клеточных движений, например, актиновые нити (филаменты) обеспечивают мышечное сокращение, а микротрубочки являются основными структурными и силовыми элементами, обусловливающими движение ресничек и жгутиков - длинных выростов на поверхности некоторых клеток, биения которых напоминают удары бича.

Рис. 1-25. На этой микрофотографии, полученной с помощью техники глубокого травления, видна сеть актиновых филаментов, подстилающая плазматическую мембрану животной клетки. (С любезного разрешения John Heuser.)
Актиновые нити и микротрубочки существенны также для внутренних перемещений, которые имеют место в цитоплазме эукариотических клеток. Так, микротрубочки в форме митотического веретена - важнейшая часть аппарата, обеспечивающего правильное распределение ДНК в процессе деления эукариотических клеток. Без микротрубочек эукариотические клетки просто не могли бы воспроизводиться, поскольку движение путем простой диффузии было бы либо слишком медленным, либо слишком неупорядоченным. Существует предположение, что большинство органелл эукариотических клеток прямо или косвенно прикреплены к цитоскелету и могут перемещаться только вдоль его направляющих элементов.
1.2.10. К царству простейших относятся наиболее сложные из известных клеток
Насколько сложно может быть устроена одиночная эукариотическая клетка, легче всего проиллюстрировать на примере протистов (рис. 1-26). Это свободноживущие одноклеточные эукариоты, возникшие разными эволюционными путями (см. рис. 1-16) и демонстрирующие чрезвычайное разнообразие форм и поведения: они могут быть фотосинтезирующими и хищными, подвижными и прикрепленными. Часто их характеризует сложное строение, например они могут включать чувствительные щетинки, фоторецепторы, жгутики, ногоподобные отростки, ротовой аппарат, жалящие иглы и мышечноподобные пучки сократительных волокон. Хотя все протисты являются одноклеточными, они могут быть организованы столь же сложно, как многие многоклеточные организмы. Все это особенно верно в отношении группы протистов, называемых простейшими, или «первыми животными».
Didinium - хищное простейшее. Оно имеет округлое тело диаметром около 150 мкм, окаймленное двумя рядами ресничек, передний край уплощен и имеет одиночный рылообразный выступ (рис. 1-27). Didinium быстро плавает в воде благодаря синхронным биениям своих ресничек. Встретив подходящую жертву (обычно другое простейшее, Paramecium), хищник выпускает из своего рыльца множество маленьких парализующих жал. Затем Didinium прикрепляется к Paramecium и пожирает ее, выворачиваясь как полый шар, чтобы поглотить клетку почти такого же размера, как он сам. Большую часть этого сложного поведения плавание, парализацию и поглощение жертвы - обеспечивают цитоскелетные структуры, подстилающие цитоплазматическую мембрану. Этот кортикальный слой включает, например, параллельные пучки микротрубочек, образующие сердцевину каждой реснички и обусловливающие способность к биениям.
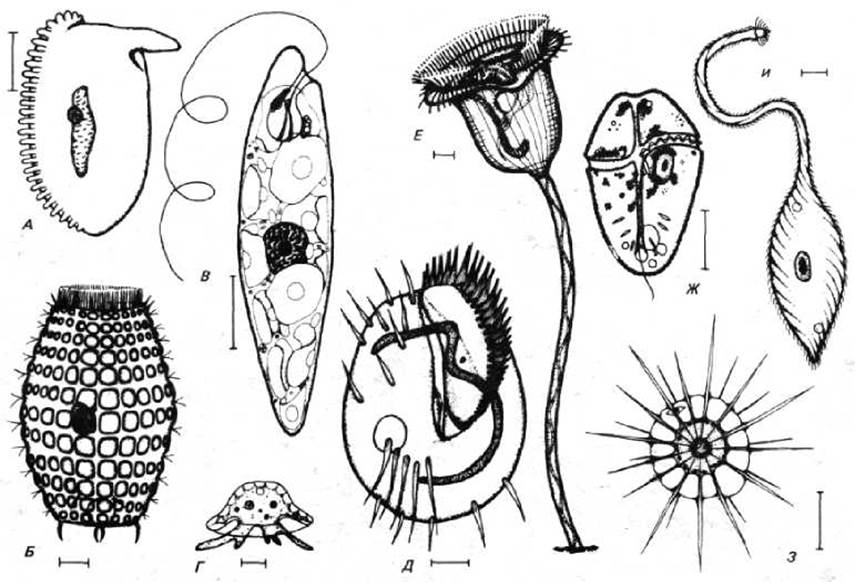
Рис. 1-26. Набор простейших, демонстрирующий огромное разнообразие этого класса одноклеточных организмов. Рисунки сделаны с разным увеличением, но для каждого организма приводится шкала в 10 мкм. Организмы А, Б, Д, Е и И принадлежат к реснитчатым, В - к эвгленовидным, Г - амебам, Ж - жгутиковым, З - солнечникам. (Из М. A. Sleigh, The Biology of Protozoa. London: Edward Arnold, 1973.)
Хищное поведение Didinium, а также свойства, благодаря которым оно реализуется: крупный размер, способность к фагоцитозу и способность двигаться, преследуя добычу, присущи исключительно эукариотам. Весьма вероятно, что эти свойства возникли очень рано в эволюции эукариот и именно это сделало возможным захват ими бактерий для «одомашнивания» в качестве митохондрий и хлоропластов.
1.2.11. Гены можно включать и выключать
Заметим, что простейшие при всей сложности их организации - это далеко не вершина эволюции эукариот. Более впечатляющие достижения на эволюционном пути были получены не путем концентрирования всяческих усложнений в одиночной клетке, а путем разделения обязанностей между различными типами клеток. Эволюция создала многоклеточные организмы, клетки которых, хотя и имеют одинаковое происхождение, претерпевают дифференцировку, в результате чего они становятся специализированными для выполнения определенных функций и образуют соответствующие органы.

Рис. 1-27. Приведенная электронная микрофотография получена с помощью сканирующего электронного микроскопа. Видно, как простейшее пожирается другим простейшим. Простейшие - это одноклеточные животные, которым присуще поразительное разнообразие форм и поведения. Ресничная инфузория Didinium (слева) имеет два периферических кольца подвижных ресничек и рылообразный выступ спереди, с помощью которого она захватывает добычу. Справа показано, как Didinium заглатывает другое простейшее, Paramecium. (С любезного разрешения D. Barlow.)
Различные специализированные типы клеток одного и того же высшего растения или животного часто выглядят совершенно по-разному (схема 1-2). Это кажется парадоксальным, поскольку все клетки многоклеточного организма являются потомками одной и той же клетки-предшественницы, а именно оплодотворенной яйцеклетки. Общее происхождение подразумевает наличие одинаковых или сходных генов. Как же возникают различия? В редких случаях клетки при специализации теряют часть генетического материала крайним примером могут служить эритроциты млекопитающих, у которых в ходе дифференцировки теряется ядро. Однако подавляющее большинство клеток почти всех видов растений и животных сохраняет всю генетическую информацию, содержащуюся в оплодотворенной яйцеклетке. В основе специализации лежат не потеря или приобретение генов, а изменение в их экспрессии.
Даже бактерии не синтезируют постоянно все возможные типы белков и способны регулировать уровень происходящих в них процессов в соответствии с условиями внешней среды. Например, белки, необходимые для усваивания (метаболизма) лактозы, производятся некоторыми бактериями только в том случае, если этот сахар имеется в среде в виде единственного углевода. Другие бактерии, попав в неблагоприятные условия, прекращают большинство нормальных метаболических процессов и образуют споры, имеющие плотную непроницаемую наружную стенку и цитоплазму измененного состава.
У эукариотических клеток развились куда более сложные механизмы контроля экспрессии генов, затрагивающие целые системы взаимодействующих генных продуктов. Как внешние, так и внутренние сигналы активируют или подавляют группы генов. При дифференцировке клеток должны координированно измениться и состав мембран, и цитоскелет, и секретируемые продукты, и даже метаболизм. Сравните, например, приспособленную к сокращению клетку скелетной мышцы и остеобласт - секретирующий твердый матрикс кости - у одного и того же животного (схема 1-2). Столь радикальные различия в типе клеток обусловлены стабильными изменениями в экспрессии генов. Механизмы, контролирующие такие изменения, развились у эукариот до беспрецедентной для прокариот степени.
1.2.12. Клетки эукариот содержат значительно больше ДНК, чем это необходимо для кодирования белков
В эукариотических клетках очень много ДНК. Как мы уже говорили, в клетках человека ее почти в 1000 раз больше, чем в типичной бактериальной клетке, а в клетках некоторых амфибий в 10 раз больше, чем клетках человека (рис. 1-28). Однако, по-видимому, лишь малая часть этой ДНК (возможно, около 1 % в клетках человека) действительно кодирует белки. Зачем же нужны остальные 99% ДНК? Одна из гипотез состоит в том, что она просто увеличивает массу ядра. По другой гипотезе, эта ДНК - собрание бесполезных для клетки последовательностей, веками накапливавшихся в ней в результате использования клеточных механизмов синтеза для собственного размножения. Действительно, в ДНК многих видов обнаружены так называемые мобильные (подвижные) элементы - последовательности, способные внезапно «перепрыгивать» из одного участка ДНК в другой и даже встраивать в новые места свои дополнительные копии. Мобильные элементы могут, таким образом, размножаться, подобно медленно развивающейся инфекции, составляя все более увеличивающуюся часть генетического материала.

Схема 1-2. Некоторые типы клеток позвоночных животных.
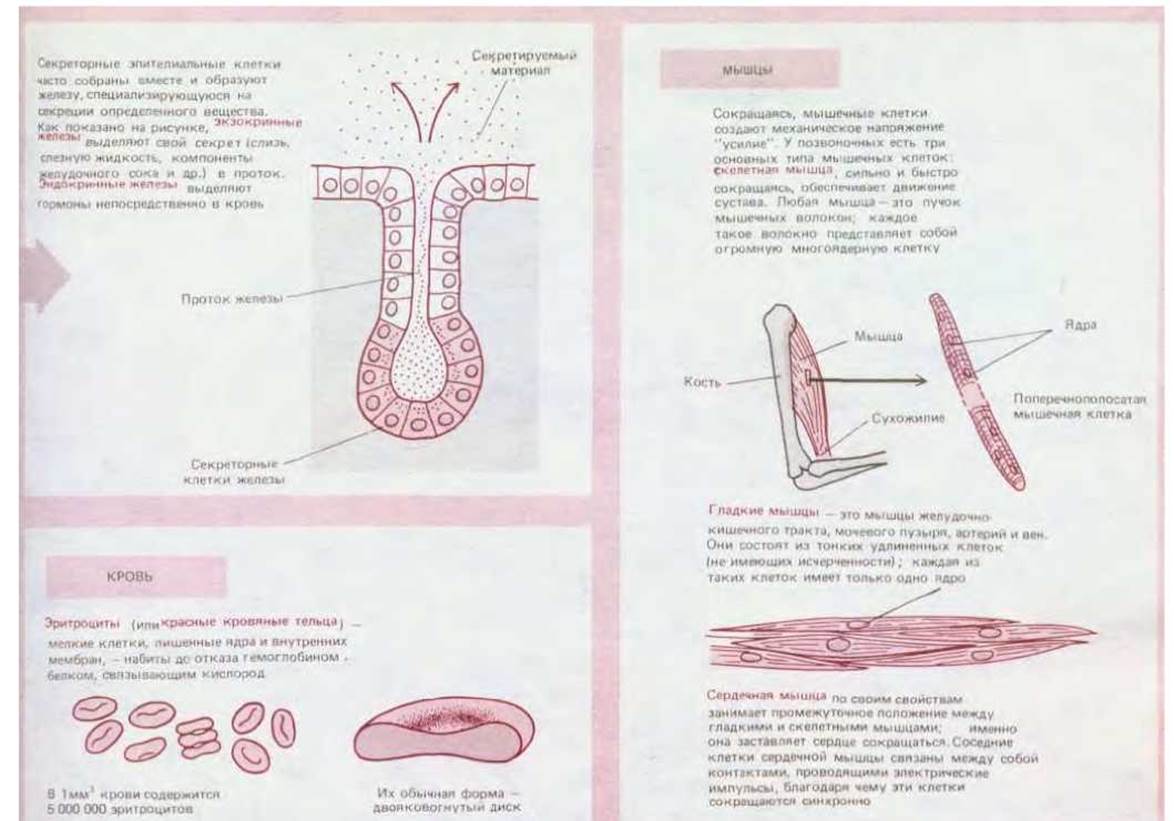
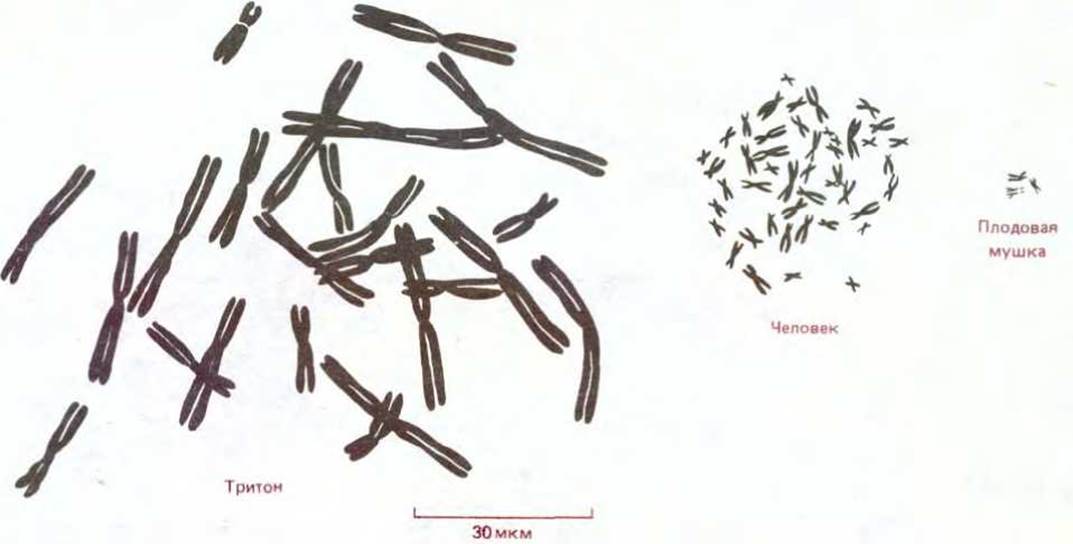
Рис. 1-28. Сравнение хромосом тритона, человека и плодовой мушки, представленных в одном масштабе. Если судить по общему размеру хромосом, на одну клетку у человека приходится больше ДНК, чем у плодовой мушки, но меньше, чем у тритона. Некоторые растения, такие, как лилия, имеют больше ДНК, чем некоторые животные. (По данным Н. С. Macgregor, Philos. Trans. R. Soc. Lond. (Biol.), 283, 309-318, 1978.)
Но эволюция использует все возможности. Вне зависимости от происхождения некодирующей ДНК сейчас она, наверняка, выполняет какую-то важную функцию. Например, часть этой ДНК играет структурную роль, позволяя генетическому материалу конденсироваться или «упаковываться» определенным образом. Другая часть «лишней» ДНК - регуляторная и участвует во включении и выключении генов, направляющих синтез белков, играя, таким образом, ключевую роль в сложных механизмах регуляции экспрессии генов эукариотической клетки.
1.2.13. Генетический материал эукариотических клеток упакован очень сложно
ДНК эукариотических клеток имеет столь большую длину, что риск ее запутывания или разрыва очень велик. Вероятно, по этой причине в процессе эволюции возникли гистоны - характерные для эукариот белки, связывающиеся с ДНК и скручивающие ее в компактные и более удобные для ряда клеточных процессов хромосомы (рис. 1-29). Уплотнение хромосом является существенной частью подготовки эукариотических клеток к делению (рис. 1-30). ДНК всех эукариот (за одним небольшим исключением) связана с гистонами. Об исключительно важном значении этих белков свидетельствует тот факт, что они ведут себя крайне консервативно в процессе эволюции: некоторые из гистонов гороха идентичны (с точностью до аминокислоты) соответствующим гистонам коровы.
В эукариотических клетках с ДНК кроме гистонов связано множество других белков. Некоторые из них, влияя на способность ДНК взаимодействовать с разнообразными молекулами, изменяют набор экспрессирующихся генов, отличающий один тип специализированных клеток от другого.

Рис. 1-29. Схема, показывающая, как положительно заряженные белки, называемые пистонами, способствуют скручиванию ДНК в хромосомах.
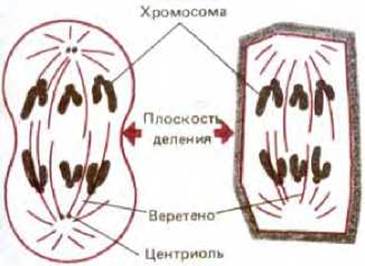
Рис. 1-30. Схематическое изображение эукариотической клетки в митозе. Слева - животная клетка, справа - растительная. Ядерная оболочка разрушена, а реплицированная ДНК сконденсирована в два полных набора хромосом. Митотическое веретено, состоящее в основном из микротрубочек, распределяет по одному набору хромосом в каждую из новообразующихся клеток.
Мембраны, окружающие ядра эукариотических клеток, защищают связанный с ДНК тонкий механизм контроля от многих происходящих в цитоплазме химических изменений. Кроме того, они позволяют пространственно разобщить две ключевые стадии экспрессии генов: 1) синтез РНК по матрице ДНК (транскрипцию ДНК) и 2) использование этих последовательностей РНК для синтеза определенных белков (трансляцию РНК). В прокариотических клетках нет такой компартментации и трансляция РНК с образованием белка происходит по мере образования РНК при транскрипции, начинаясь раньше, чем завершился синтез РНК. У эукариот, напротив (за исключением митохондрий и хлоропластов, которые в этом отношении, как и в других, ближе к бактериям), указанные этапы пути от гена к белку строго разобщены: транскрипция происходит в ядре, трансляция - в цитоплазме. РНК, прежде чем включиться в процессы синтеза белка, должна покинуть ядро. Для этого, находясь в ядре, РНК претерпевает сложный процесс созревания (процессинг), в ходе которого одни части молекулы РНК удаляются, а другие модифицируются.
Сложность организации генома в клетках эукариот обусловливает присутствие в них более тонких и разнообразных механизмов регуляции нежели те, которые обнаруживаются в клетках бактерий.
Заключение
Все существующие ныне клетки подразделяются на два типа: прокариотические (бактерии и их близкие родственники) и эукариотические. Считают, что первые близки в общих чертах с самыми ранними клетками-прародительницами. Несмотря на сравнительную простоту строения, клетки прокариот весьма разнообразны в биохимическом отношении, например, у бактерий можно обнаружить все основные метаболические пути, включая три главных процесса получения энергии - гликолиз, дыхание и фотосинтез. Эукариотические клетки больше по размеру и имеют более сложную организацию, чем клетки прокариот. Они содержат больше ДНК и различных компонентов, обеспечивающих ее сложные функции. ДНК эукариот заключена в окруженное мембраной ядро, а в цитоплазме находится много других окруженных мембранами органелл. К ним относятся митохондрии, осуществляющие окончательное окисление молекул пищи, а также (в растительных клетках) хлоропласты, в которых идет фотосинтез. Целый ряд данных свидетельствует о происхождении митохондрий и хлоропластов от ранних прокариотических клеток, ставших внутренними симбионтами большей по размеру анаэробной клетки. Другая отличительная особенность эукариотических клеток - это наличие цитоскелета из белковых волокон, организующего цитоплазму и обеспечивающего механизм движения.