Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Внутриклеточная сортировка макромолекул и сохранение клеточных компартментов
Структура хромосомы
После обсуждения структуры ДНК и белков, входящих в состав хромосомы, рассмотрим ее строение в целом. Оказывается, ДНК в хромосоме не только упакована с помощью гистонов в регулярно повторяющиеся нуклеосомы, но, кроме того, хитроумным способом организована вместе с другими белками в серию субдоменов, обладающих различными свойствами. Эти структуры более высокого порядка являются удивительной особенностью хроматина эукариотических клеток; до сих пор остается загадкой, как именно такие домены функционируют.
9.2.1.Хромосомы, по-видимому, состоят из серии петель [21]
Диаметр ядра обычно не превышает 5 мкм (5 х 104 см). Поскольку упаковка ДНК в хроматиновой фибрилле позволяет уменьшить ее линейные размеры до 1 мм, должны существовать другие, более высокие, уровни компактизации. Один из принципов дальнейшей конденсации хроматина был подсказан внешним видом некоторых особых хромосом - так называемых хромосом типа ламповых щеток из ооцитов многих животных и политенных хромосом определенных клеток насекомых. Хромосомы этих двух типов обладают хорошо выраженной петельной структурой, т. е. имеют серию петельных доменов, которые под углом отходят от основной оси хромосомы. Установлено, что хромосома бактерии E.coli (кольцевая молекула ДНК длиной около 0,1 см, лишенная гистонов) также уложена в виде петель. Существующие в настоящее время данные позволяют считать петельную укладку общим принципом структурной организации хроматина.
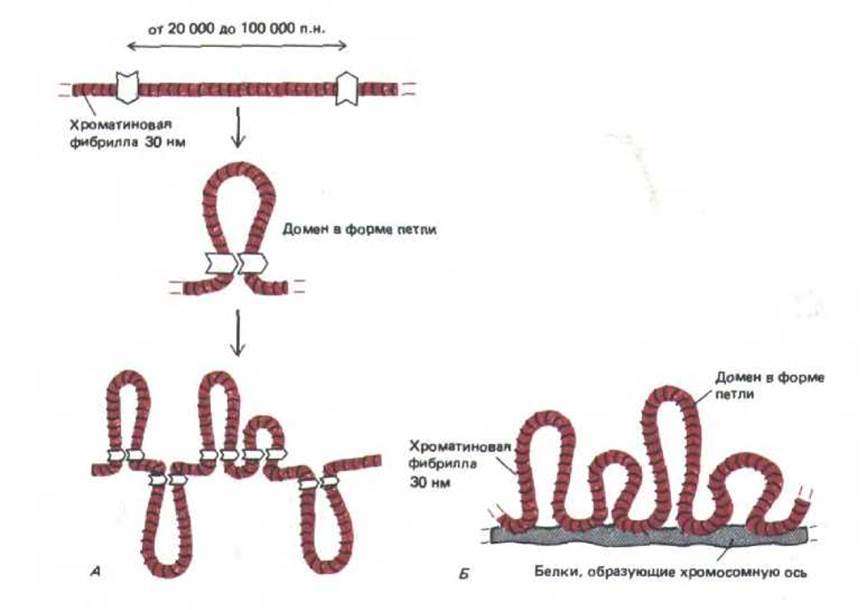
Рис. 9-34. Схематическое изображение участка хромосомы, имеющего петельную организацию. Каждая из петель содержит приблизительно от 20000 до 100000 пар оснований двухцепочечной ДНК, входящей в состав 30 нм-хроматиновой фибриллы. А. Модель упаковки, согласно которой на каждом конце петли находится сайт-специфический ДНК-связывающий белок. Б. Модель упаковки с участием хромосомной оси. В настоящее время неизвестно, как в действительности происходит упаковка, хотя цитологические данные свидетельствуют о том, что осевой участок изолированных митотических хромосом (место локализации концов петель) сильно обогащен ферментом, который в клетке содержится в большом количестве,-ДНК топоизомеразой ll.
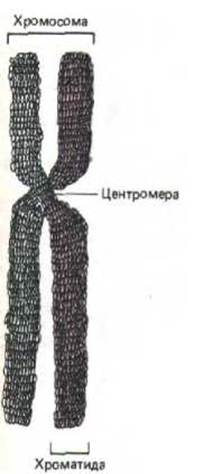
Рис. 9-35. Схематическое изображение типичной метафазной хромосомы. Каждая хроматида содержит одну из двух идентичных дочерних молекул ДНК (одна из них выделена цветом], образующихся в процессе репликации на более ранней стадии клеточного цикла.
Была выдвинута гипотеза, предполагающая, что петли хроматина формируются и поддерживаются с помощью ДНК-связывающих белков, которые узнают определенные нуклеотидные последовательности двух отдельных участков хроматиновой фибриллы и сближают их (рис. 9-34, А). В результате эти участки образуют устье петли. Петли могут сформироваться и иным путем при связывании ДНК с осью хромосомы (рис. 9-34, Б). По косвенным оценкам у таких таксономически отдаленных организмов, как дрозофила и человек, средняя длина петель весьма сходна, типичная петля содержит примерно от 20000 до 100000 пар оснований, что соответствует приблизительно 0,5 мкм фибриллы диаметром 30 нм. Если типичная хромосома человека состоит главным образом из петель, то в ее состав могло бы входить более 2000 таких доменов.
9.2.2. Митотические хромосомы состоят из максимально сконденсированного хроматина [22]
Большинство хромосом чрезвычайно растянуты и спутаны, что делает их невидимыми на всех фазах клеточного цикла за исключением митоза. В этот период хромосомы спирализуются, конденсируются и приобретают четкие формы. Эта суперспирализация, уменьшающая линейные размеры ДНК с 5 см по 5 мкм, сопровождается фосфорилированием всех молекул гистона Н1, присутствующих в клетке, по пяти сериновым остаткам. В связи с тем, что гистон Н1 связывает между собой нукдеосомные частицы (см. рис. 9-28), его фосфорилирование может играть ключевую роль в конденсации хромосом в процессе митоза. На рис. 9-35 изображена типичная митотическая хромосома на стадии метафазы. Две дочерние молекулы ДНК упаковываются порознь и образуют сестринские хроматиды, которые удерживаются вместе с помощью центромеры. Митотические хромосомы обычно несут на своей поверхности множество других молекул, в том числе большое количество рибонуклеопротеинов. Если удалить эту оболочку, на электронных микрофотографиях можно отчетливо увидеть, что каждая хроматида построена из хроматиновых петель, отходящих от центральной оси (рис. 9-36 и 9-37). Экспериментально доказано, что поперечная исчерченность, характерная для митотических хромосом, отражает в какой-то степени порядок расположения генов в молекуле ДНК. Различные способы упаковки длинной спирали ДНК представлены на рис. 9-38.
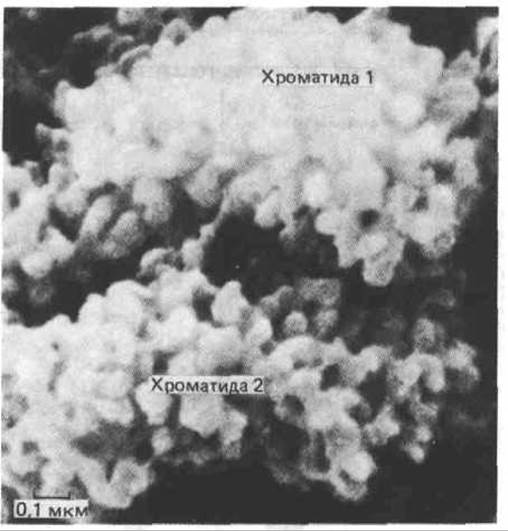
Рис. 9-36. Микрофотография участка типичной высококонденсированной митотической хромосомы, полученная с помощью сканирующего электронного микроскопа. Предполагается, что каждый выступ, напоминающий клубенёк, представляет собой конец отдельного петлевого домена. Обратите внимание, как хорошо различимы две идентичные спаренные хроматиды, схема которых представлена на рис. 9-35. (По Mardsen M. Р., Laemmli U.K. Cell 17: 849-858, 1979.)
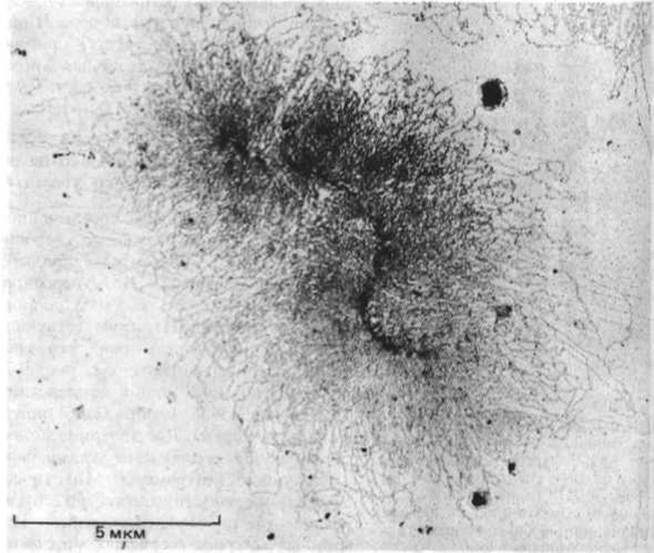
Рис. 9-37. Электронная микрофотография одиночной хроматиды митотическай хромосоми насекомого (Oncopelfus). Специальная обработка позволяет визуализировать петли хроматина, отводящае от центральной оси хромагиды. (С любезного разрешения Victoria Foe.)
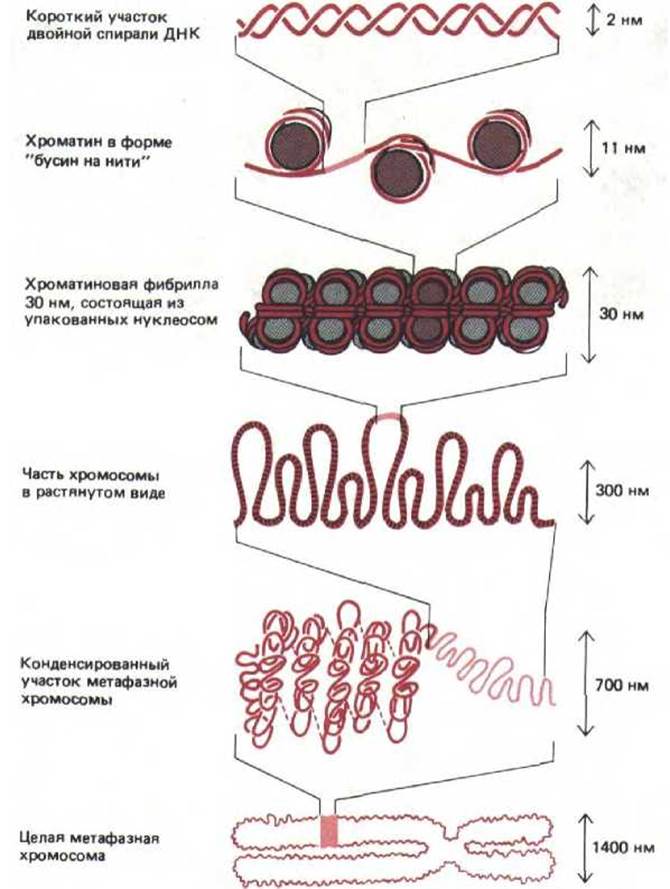
Рис. 9-38. Схема, иллюстрирующая различные уровни упаковки хроматина, которые, по-видимому, отражают последовательные этапы формирования высококонденсированной метафазной хромосомы.
В митотической хромосоме хроматин транскрипционно неактивен: синтез РНК с началом конденсации хромосом прекращается. По-видимому, РНК-полимераза не может в этих условиях продвигаться вперед по ДНК, хотя возможно это обусловлено и другими причинами.
9.2.3. Каждая митотическая хромосома содержит определенный набор очень больших доменов [23]
Набор из 46 митотических хромосом человека называют его кариотипом. Цитологические методы, разработанные в начале 70-х гг., позволяют безошибочно идентифицировать каждую хромосому в кариотипе человека. Для этого специально подготовленные препараты митотических хромосом обрабатывают красителями, способными флуоресцировать только при связывании с определенными типами последовательностей ДНК. Несмотря на то, что эти красители обладают очень низкой избирательностью [их использование позволяет в основном отличать участки ДНК, обогащенные АТ-парами (G-полосы) от участков с GC-богатыми последовательностями (R-полосы)], обработка ими митотических хромосом выявляет в каждой из них характерную картину чередования темных и светлых полос (сегментов) (рис. 9-39). Распределение этих полос специфично для каждого типа хромосом, что делает возможной их однозначную идентификацию (рис. 9-40).
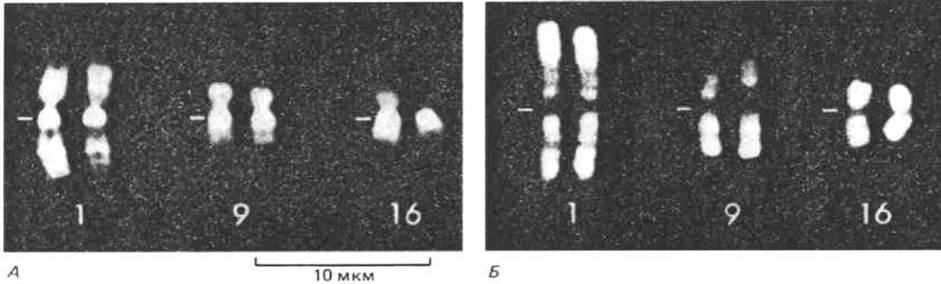
Рис. 9-39. Микрофотографии трех пар митотических хромосом человека, полученные с помощью флуоресцентного микроскопа. А. Окрашивание хромосом АТ-специфическим красителем Hoescht 33258 (G-полосы). Б. Окрашивание хромосом GC-специфическим красителем оливомицином (R-полосы). Черта указывает положение центромеры. Обратите внимание на то, что картины распределения сегментов (полос) в хромосомах на обеих фотографиях комплементарны: полосы, яркоокрашенные на А, на Б затемнены и наоборот. G-полосы проявляются и при окрашивании красителем Гимза (отсюда их название), а обозначение полос буквой R отражает тот факт, что они как бы обратны (reverse) G-пoлосам. (По K.F. Jorgenson, J.H. van de Sande and С. С. Zin, Chromosoma 68: 287-302, 1978.)
При анализе хромосом человека, проведенном с помощью флуоресцентных красителей, на ранних стадиях митоза, когда хромосомы значительно менее компактны, чем в метафазе, удалось подсчитать, что весь гашюидный набор содержит по меньшей мере 2000 четко выраженных полос, соответствующих АТ-богатым последовательностям ДНК.
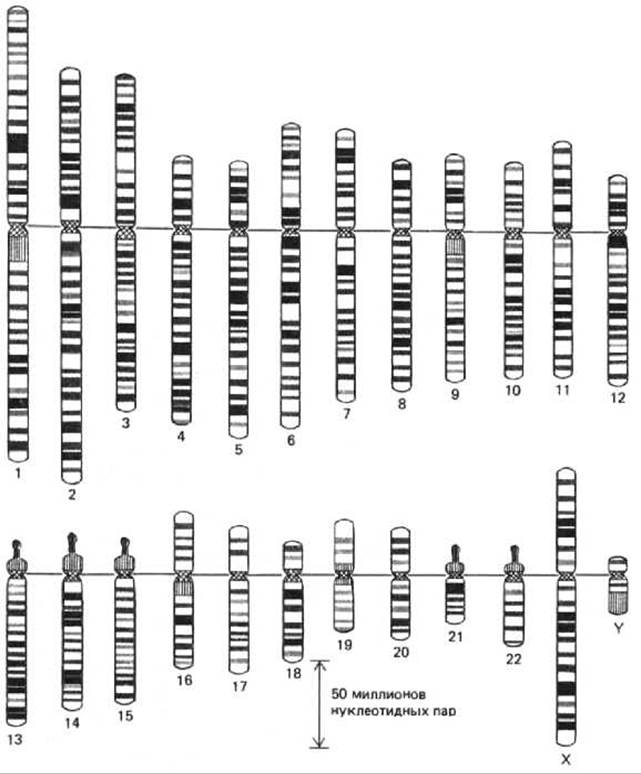
Рис. 9-40. Стандартная карта распределения сегментов в каждой из хромосом, составляющих человеческий кариотип, на стадии прометафазы митоза. Хромосомы от 1-й до 22-й пронумерованы в примерном соответствии с размером. Диплоидная клетка человека содержит по две хромосомы каждого типа плюс две Х-хромосомы (у женщин) или Х- и Y-хромосомы (у мужчин). 850 полос, указанных на рисунке - это G-полосы, обнаруживаемые при окрашивании реагентами, специфичными к АТ-последовательностям ДНК. Выделенные цветом «головки» на хромосомах 13, 14, 15, 21 и 22 указывают на расположение генов, кодирующих большие рибосомные РНК, цветными линиями отмечено положение центромеры на каждой хромосоме. (С изменениями по U. Franke, Cytogenet. Cell Genet. 31: 24-32, 1981.)
В ходе митоза происходит дальнейшая конденсация хромосом и, как следствие, утолщение полос и уменьшение их числа.
Наличие поперечной исчерченности составляет общую черту миготических хромосом даже таких далеких друг от друга видов, как человек и дрозофила. Более того, картина распределения полос в хромосомах почти не изменилась за долгие периоды эволюции. Например, почти каждой хромосоме человека соответствует аналог в кариотипе шимпанзе, гориллы и орангутана (хотя в результате слияния одной пары хромосом у человека имеется 46, а не 48 хромосом, как у обезьян), причем картина распределения полос у них практически одинакова. Все это лишний раз указывает на большое значение пространственной организации ДНК в составе хромосом для экспрессии соответствующих генов, а само существование полос, возможно, отражает какие-то черты функциональной организации хроматина. Почему образуются такие полосы, до сих пор является загадкой. Даже наиболее тонкие полосы, изображенные на рис. 9-40, должны содержать не менее 30 петель, а суммарный нуклеотидный состав столь длинных последовательностей ДНК (более миллиона пар оснований, что соответствует размеру среднего бактериального генома), по-видимому, близок к среднестатистическому. Известно, что как АТ-богатые, так и GC-богатые полосы содержат гены.
9-13
9.2.4. ДНК хромосом типа ламповых щеток в интерфазе состоит из серии различающихся доменов [24]
Несмотря на высокий порядок упаковки хроматина, нити его в период интерфазы слишком тонки и спутанны, чтобы можно было ясно увидеть целиком всю хромосому. Тем не менее, существуют определенные типы клеток, в которых общую структуру интерфазных хромосом различить можно. Например, спаренные в мейозе хромосомы растущих ооцитов (незрелые яйцеклетки), активно синтезируют РНК и образуют необычайно жесткие и протяженные петли хроматина, покрытые вновь транскрибируемой РНК, которая упакована в плотные комплексы РНК-белок. В связи с тем, что ДНК покрыта такими комплексами, хромосомы (их называют хромосомами типа ламповых щеток) хорошо видны даже под световым микроскопом (рис. 9-41).
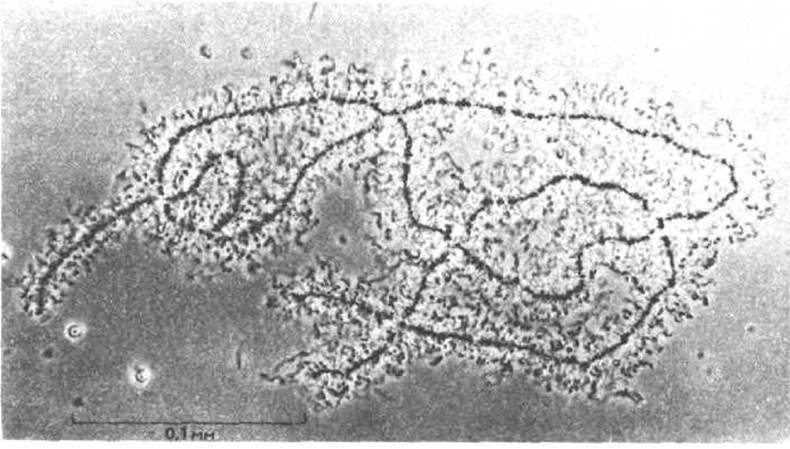
Рис. 9-41. Световая микрофотография хромосом типа ламповых щеток из ооцита земноводного. На ранних стадиях дифференцировки ооцитов каждая хромосома перед началом мейоза рсплицируется, ад гем гомологи спариваются с образованием четыреххроматидной протяженной структуры, изображенной на фотографии. Стадия хромосом типа ламповых щеток может длиться месяцы и даже годы. В это время в ооците создается запас мРНК и других веществ, необходимых для его последующего развития в новую особь. Обратите внимание, что каждая хромосомная ось имеет в длину приблизительно 400 мкм, тогда как длина большинства митотических хромосом составляет менее 10 мкм. (С любезного разрешения Joseph G. Gall.)
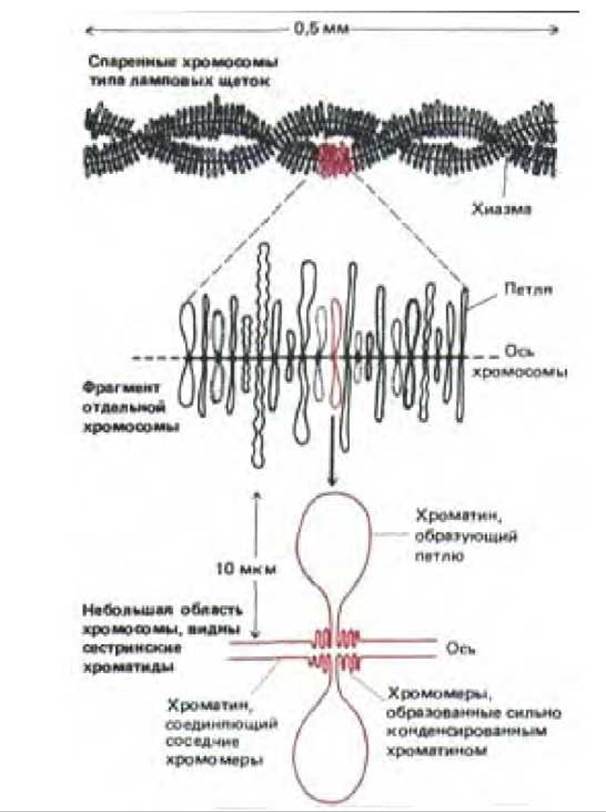
Рис. 9-42. Строение хромосомы типа ламповых щеток. У многих земноводных набор хромосом типа ламповых щеток содержит в сумме около 10000 петель хроматина, хотя большая часть ДНК остается в сильно конденсированном состоянии и располагается в хромомерах. Каждая петля соответствует определенной последовательности ДНК. В каждой клетке содержится по четыре копии каждой петли, поскольку структура, представленная в верхней части рисунка, состоит из двух спаренных гомологичных хромосом, а каждая хромосома представлена двумя сестринскими хроматидами. Такие структуры, состоящие из четырех нитей, характерны для данной стадии развития ооцита (стадия диплонемы мейоза).
Схема строения хромосом типа ламповых щеток приведена на рис. 9-42. Большие петли, состоящие из деконденсированного хроматина, отходят в стороны от оси хромосомы. Опыты по гибридизации нуклеиновых кислот показали, что определенная петля всегда содержит одну и ту же последовательность ДНК, которая во время роста ооцита располагается строго определенным образом. Следовательно, эти петли соответствуют фиксированным единицам упаковки хроматина, который деконденсировался и стал транскрипционно активным. Поскольку петля среднего размера содержит приблизительно 100 000 пар оснований, каждая петля может соответствовать одной петле хроматина, описанного выше (см. разд. 9.2.1). Многие петли постоянно транскрибируются по всей длине, другие содержат протяженные участки хроматина, который не транскрибируется вовсе. Большая часть хроматина не входит в состав петель и остается в сильно конденсированном состоянии в хромомерах; этот хроматин, как правило, не транскрибируется. Короткие области хроматина, которые не обладают высокой степенью конденсации и активно не транскрибируются, соединяют соседние хромомеры вдоль хорошо выраженной оси хромосомы.
Хромосомы типа ламповых щеток являются необычными в том отношении, что уровень их транскрипции выше, а большинство образующихся транскриптов РНК длиннее, чем те, которые синтезируются на других хромосомах. Однако есть данные, что молекула ДНК, входящая в состав любых интерфазных хромосом, тоже подразделяется на различные области, каждая из которых отделена от своих соседей границей. По-видимому, и в данном случае хроматин в разных областях упакован по-разному (например, в виде петель, хромомеров или хроматина, входящего в состав оси между хромомерами).
9.2.5. В политенных хромосомах также можно увидеть упорядоченные участки интерфазного хроматина [25]
Структура хроматина на уровне одиночных петель очень хорошо различима и в некоторых клетках насекомых. Многие клетки личинок мух вырастают до необычайно большого размера, претерпевая несколько циклов репликации ДНК, которые не сопровождаются клеточным делением. В результате они содержат в несколько тысяч раз больше ДНК, чем обычная клетка. Такие гигантские клетки называются полшлоидными в том случае, если число наборов хромосом у них превышает норму. Ситуация может быть и иной: гомологичные хромосомные пары могут не отделяться друг от друга, а формировать единые огромные хромосомы (политенные хромосомы). Тот факт, что в отдельных гигантских клетках насекомых хромосомы могут переходить из политенного в полишюидное состояние, свидетельствует о том, что эти два хромосомных статуса тесно связаны между собой и что структура политенных хромосом в принципе аналогична структуре нормальных хромосом.
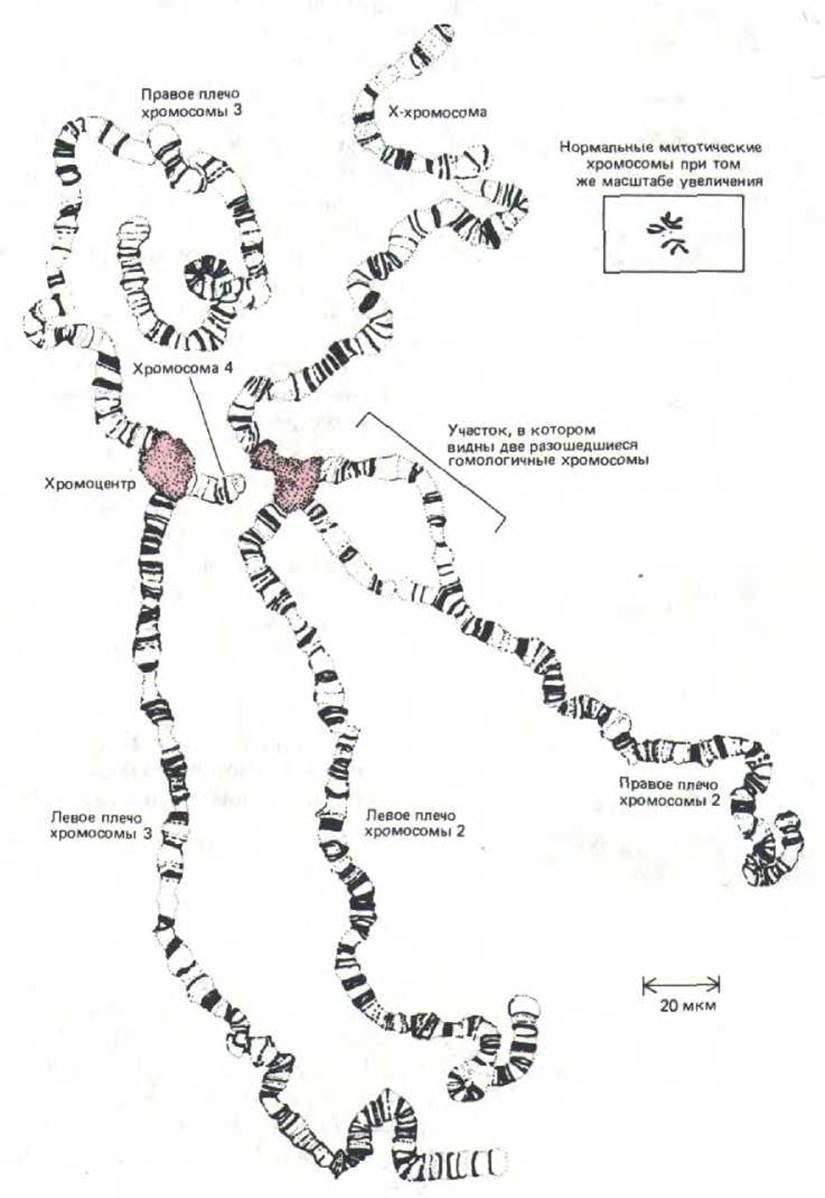
Рис. 9-43. Полный набор политенных хромосом из клетки слюнной железы дрозофилы. Эти хромосомы были расправлены и подготовлены для наблюдения путем «раздавливания» материала на предметном стекле. На рисунке представлены четыре пары хромосом. Каждая хромосома тесно спарена со своим гомологом (так, что каждая пара кажется единой структурой), чего не наблюдается в большинстве интерфазных ядер. Четыре пары хромосом связаны друг с другом своими центромерными зонами, образующими один большой «хромоцентр» (окрашенная область). На данном препарате хромоцентр разделился на две части в процессе подготовки к микроскопированию. Необходимо отметить, что при выстраивании бок о бок многих нитей хроматина произошло значительное раскручивание каждой молекулы ДНК. (С изменениями по Т. S. Painter, J.Hered. 25: 465-476, 1934.)

Рис. 9-44. Световая микрофотография участка политенной хромосомы из клетки слюнной железы дрозофилы. Видна характерная картина распределения дисков. Эти диски обнаруживаются в интерфазных хромосомах и являются отличительной чертой гигантских политенных хромосом; их не следует отождествлять с гораздо более «грубыми полосами, показанными на рис. 9-40, которые обнаруживаются в нормальных митотических хромосомах с помощью специальных красителей. (С любезного разрешения Joseph G. Gall.)

Рис. 9-45. Электронная микрофотография небольшого участка полигенной хромосомы дрозофилы. На тонком срезе видно, что разные хромосомные диски, весьма различающиеся по толщине (В), разделены междисковыми участками (I), хроматин которых гораздо менее сконденсирован. (С любезного разрешения Viekko Sorsa.)
Благодаря своей большой величине, а также тому, что плотная и строго упорядоченная упаковка индивидуальных хроматшювых нитей в составе политенной хромосомы препятствует их запутыванию, эти хромосомы очень хорошо видны в световой микроскоп. Как и хромосомы типа ламповых щеток, политенные хромосомы в интерфазе активно синтезируют РНК. Явление политении наиболее глубоко было изучено на четырех хромосомах клеток слюнных желез личинок дрозофилы. В этих клетках ДНК реплицируется 10 раз подряд, дочерние хромосомы не разделяются, в результате чего образуются гигантские хромосомы, содержащие 1024 (210) тесно прилегающие друг к другу индивидуальные хроматиновые нити.
При наблюдении окрашенных политенных хромосом в световой микроскоп хорошо заметны перемежающиеся поперечные полосы: темные (диски) и светлые (междисковые участки) (рис. 9-43 и 9-44). Каждый диск и междисковый участок состоят из 1024 идентичных последовательностей ДНК, расположенных рядом друг с другом. Около 85% ДНК в политенных хромосомах содержится в дисках и 15%-в меж дисковых участках. Хроматин каждого диска при окрашивании выглядит более темным, так как он более конденсирован, чем хроматин междисковых участков (рис. 9-45). Полагают, что диск состоит из петли, которая многократно сложена (рис. 9-46). В зависимости от размера отдельные полосы содержат от 3000 до 300000 нуклеотидных пар. Поскольку каждый диск можно идентифицировать исходя из его толщины и расположения в хромосоме, все диски могут быть пронумерованы, что дает возможность составить «карту» политенной хромосомы. Во всем геноме дрозофилы содержится примерно 5000 дисков и 5000 междисковых участков.
9.2.6. Отдельные домены хроматина в политенных хромосомах могут разворачиваться и вновь упаковываться как отдельные единицы [26]
Задолго до того, как были получены первые данные о структуре хроматина, изучение политенных хромосом позволило сформулировать гипотезу, согласно которой транскрипция генов сопровождается значительными изменениями в упаковке ДНК: отдельный диск хромосомы вздувается при активации содержащихся в нем генов и вновь конденсируется, когда гены становятся неактивными.
Для того чтобы выявить транскрибируемые области на политенной хромосоме, можно ввести в клетки радиоактивный предшественник РНК [3Н]-уридин, а затем локализовать растущий транскрипт РНК с помощью радиоавтографии (рис. 9-47). Именно таким способом было установлено, что большая часть активного хроматина находится в де-конденсированном состоянии и образует характерные хромосомные пуфы.
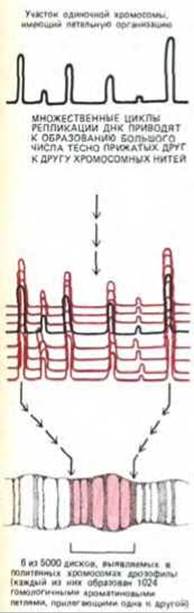
Рис. 9-46. Схема, показывающая, каким образом плотно прилегающие один к другому гомологичные петельные домены могут стать причиной появления дисков в политенных хромосомах. В составе каждого диска петли хроматиновых фибрилл находятся в очень тесном контакте друг с другом, формируя гораздо более конденсированную структуру, чем показано на этом рисунке.
Одним из основных факторов, контролирующих активность генов в политенных хромосомах дрозофилы, является гормон экдизои, встречающийся у насекомых. Уровень этого гормона в ходе развития личинки периодически поднимается и снижается, индуцируя транскрипцию разнообразных генов, которые кодируют белки, необходимые личинке для линьки и для окукливания. По мере прохождения через определенные стадии развития, возникают новые и исчезают старые пуфы, что связано с активацией и затуханием активности транскрипционных единиц и с синтезом разнообразных мРНК и белков (рис. 9-48). Изучение отдельного пуфа, размер которого относительно мал (но соответствующая ему полоса на хромосоме все-таки различима), дает основание предполагать, что каждый пуф образуется при разворачивании одного-единственного диска на хромосоме (рис. 9-49). Электронномикроскопический анализ показал, что ДНК пуфов находится в гораздо менее конденсированном состоянии, чем это свойственно хроматиновой фибрилле диаметром 30 нм (рис. 9-50). По-видимому, отдельная петля, которая, как полагают, упакована в диск на хромосоме (рис. 9-46), при транскрипции деконденсируется как самостоятельная единица.
9.2.7. Гены на политенной хромосоме расположены, вероятно, как в дисках, так и в междисковых участках [27]
Фиксированное расположение дисков и междисковых участков на политенной хромосоме дрозофилы навело цитогенетиков на мысль, что каждый диск, возможно, соответствует отдельному гену. Анализ мутаций не только подтвердил это предположение, но позволил генетикам подсчитать число жизненно важных генов у дрозофилы; их оказалось приблизительно 5000, т. е. столько же, сколько дисков на хромосомах. Например, при попытке индуцировать мутации, картируемые на небольшом участке хромосомы (содержащем около 50 различных дисков), генетическими методами было выявлено около 50 жизненно важных генов. Хотя данный метод и не позволяет определить, где именно локализован конкретный ген, в диске или в междисковом участке, эти наблюдения дают основание для следующего вывода: обычный диск может содержать последовательности ДНК, кодирующие один-единственный жизненно важный белок.
Однако последующие эксперименты заставили усомниться в правильности гипотезы «один диск - один ген». Например, была клонирована значительная область генома дрозофилы размером 315000 нуклеотидных пар, затем отдельные ее фрагменты использовали в качестве зондов для идентификации мРНК, синтезирующихся на этом участке. Количество отдельных мРНК, как выяснилось, в три раза превышает число дисков. Весьма вероятно, что большая часть этих мРНК соответствуют генам, влияющим на приспособленность мух именно в природных условиях. В лабораторных условиях, в отсутствие селективного отбора, мутации по многим из этих генов могут не улавливаться. Недавно было установлено, что мРНК синтезируется как в дисках, так и в междисковых участках.

Рис. 9-47. Синтез РНК на гигантской политенной хромосоме из слюнных желез насекомого Chironomus tentans. На представленном радиоавтографе хромосомы, меченной [3Н]-уридином, области синтеза РНК покрыты темными гранулами серебра в соответствии с активностью каждого участка. (По С. Felling, Chromosoma 15: 71-122, 1964.)

Рис. 9-48. Серия фотографий, иллюстрирующая последовательные стадии возникновения и регрессии пуфов в политенных хромосомах Drosophila melanogaster. Представлен фрагмент левого плеча хромосомы 3, содержащий пять крупных пуфов. Каждый из этих пуфов активен лишь в течение короткого периода: зафиксированная на фотографиях последовательность событий занимает 22 ч и повторяется в каждом поколении мух. (С любезного разрешения Michael Ashburner.)
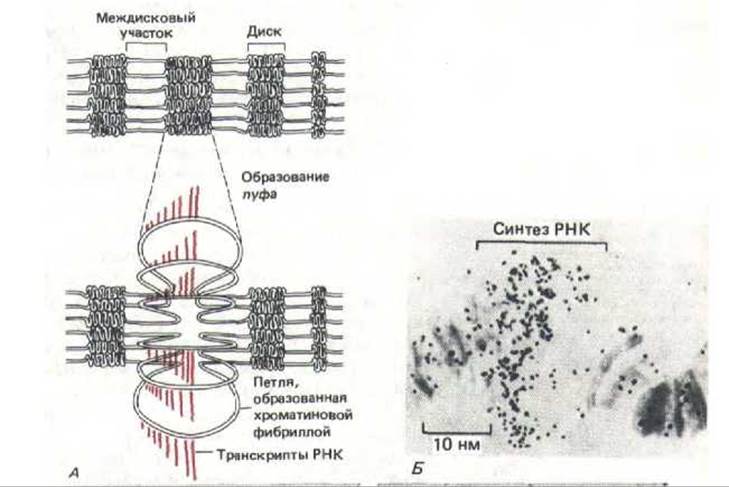
Рис. 9-49. А. Упрошенная схема процесса образования пуфа на политенной хромосоме. Б. Радиоавтограф отдельного пуфа, в котором на указанном участке синтезировалась РНК; в результате этот участок оказался меченным [3Н]-уридином. (С любезного разрешения Jose Bonner.)
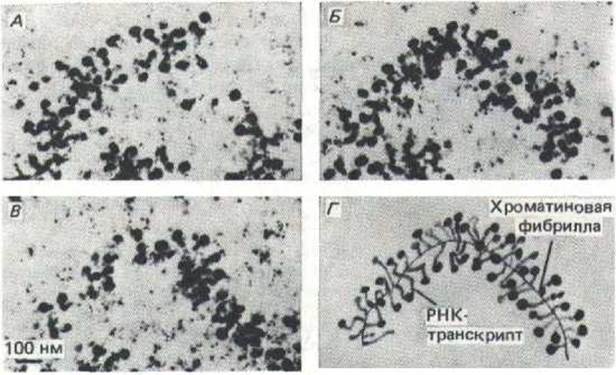
Рис. 9-50. Электронные микрофотографии серии тонких срезов крупного хромосомного пуфа, показывающие конформацию необычно длинной транскрипционной единицы. На схеме Г изображена пространственная модель исследуемого участка данной транскрипционной единицы. Концы транскриптов РНК, похожие на клубеньки, прикреплены к одиночной хроматиновой фибрилле. «Клубеньки» на 5'-конце большинства транскриптов образуются вследствие упаковки молекулы РНК в рибонуклеопротеиновые частицы. (По К. Andersson, В. Bjokroth and В. Daneholt, Exp. Cell Res. 130: 313-326, 1980.)
Хотя структурная организация хромосом наиболее четко выявляется при исследовании необычных интерфазных хромосом, таких как политенные или хромосомы типа ламповых щеток, вероятно, все хромосомы высших эукариот имеют подобную организацию. Изучение политенных хромосом дрозофилы показало, что гены могут располагаться и в области конденсированного хроматина дисков, и в менее конденсированных междисковых участках; кроме того установлено, что в каждом диске может содержаться несколько генов. Для чего же в таком случае каждый длинный тяж хроматина в хромосоме подразделяется на большое число различающихся областей? Хотя ответ и неизвестен, вполне вероятно, что такое строение дает возможность: 1) сохранять организацию ДНК; 2) отделять гены друг от друга, и таким образом избегать биологических «помех»; 3) регулировать транскрипцию генов (например, конститутивно экспрессируемые гены «домашнего хозяйства» могут располагаться в междисковых участках, тогда как гены, специфические для определенного типа клеток, могут находиться в дисках).
9-14
9.2.8. В транскрипционно активных областях хроматин менее конденсирован [28]
При обработке ядер клеток позвоночных ДНКазой I (той самой, которая при более низких концентрациях выявляет сверхчувствительные к нуклеазе области, лишенные нуклеосом) было показано, что определенная часть хроматина, содержащего активные гены, находится в де-конденсированном состоянии. Этот вывод подтверждает результаты, полученные на политенных хромосомах насекомых. При определенной концентрации ДНКазы в первую очередь подвергается деградации около 10% генома позвоночных. В разных клетках одного и того же организма нуклеаза расщепляет различные последовательности ДНК, которые соответствуют различным типам продуцируемых этими клетками РНК. С помощью гибридизации со специфическими ДНК-зондами удалось выяснить, что деградировавшие последовательности относятся главным образом к тем областям генома, которые в данном типе клеток активно транскрибируются. Интересно, что даже гены, транскрибирующиеся в каждом поколении клеток лишь несколько раз, обладают чувствительностью к нуклеазе. Этот факт говорит о том, что сверхчувствительным к нуклеазе участок становится скорее всего вследствие особого состояния хроматина, а не собственно транскрипции (рис. 9-51). Хроматин в таком чувствительном к нуклеазе состоянии часто называют активным хроматином. Полагают, что нуклеосомы, входящие в его состав, упакованы менее плотно. Даже если нуклеазной обработке подвергаются митотические хромосомы, расщепленным оказывается преимущественно активный хроматин. По-видимому, и в митозе, несмотря на чрезвычайно плотную упаковку генома, между активным и неактивным хроматином сохраняются некоторые различия.
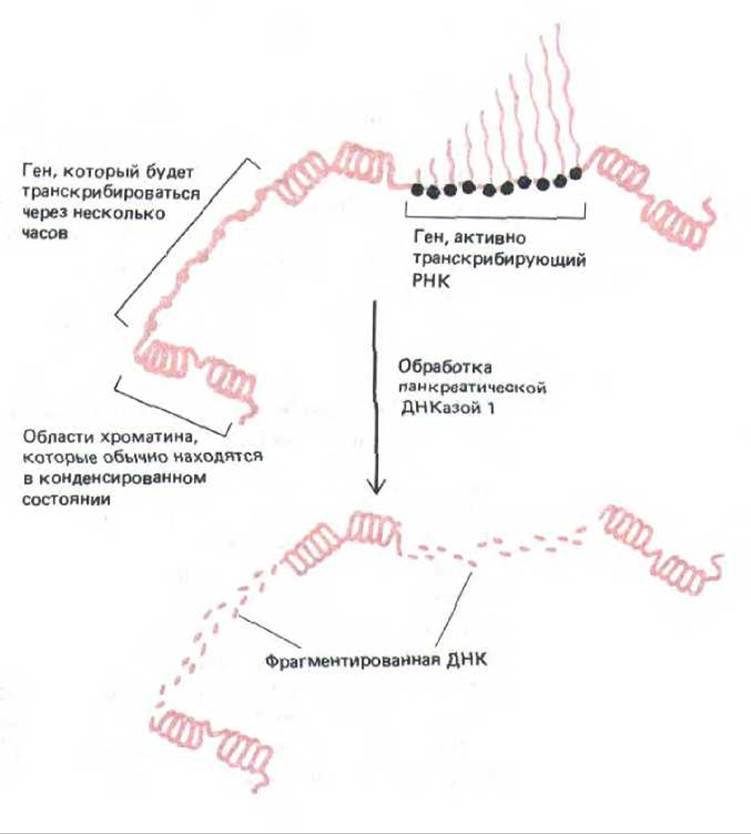
Рис. 9-51. Обработка хроматина панкреатической ДНКазой I. Вначале фермент разрезает сайты, обладающие повышенной чувствительностью к нуклеазе (не показано), затем деградации подвергается последовательность ДНК, в которую входят активно транскрибируемые и потенциально активные гены.
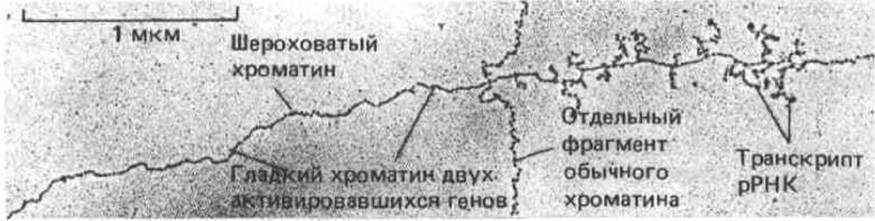
Рис. 9-52. Электронная микрофотография расправленного хроматина из клеток эмбриона насекомого (Oncopeltus). Видны значительные изменения в структуре хроматина двух тандемно расположенных генов рРНК. По-видимому, у этих эмбрионов синтезу рРНК предшествует переход хроматина из состояния «бусины на нитке» к гладкой форме. Возможно, гладкий хроматин представляет собой область, где ДНК «соскочила» с нуклеосом в ходе подготовки к транскрипции. Следует отметить, что структура хроматина, входящего в состав гена, расположенного левее от транскрибируемой области, подверглась изменениям, хотя в данный момент он не транскрибируется. (По V. Е. Foe, Cold Spring Harbor Symp. Quant. Biol. 42: 723-740, 1978.)
Электронномикроскопический анализ, как правило, не выявляет никаких изменений в строении нуклеосом, входящих в состав активного хроматина. Однако есть и поразительное исключение: у некоторых зародышей насекомых в области активных генов рРНК можно заметить протяженные участки хроматина, как будто лишенного нуклеосом (рис. 9-52). Согласно одной из гипотез, в таких участках каждая нуклеосома открывается, образуя развернутую структуру. Вероятно, при этом происходит раскручивание двух витков спирали ДНК, которые в обычном состоянии намотаны на нуклеосому. По-видимому, эти структурные изменения способствуют эффективности транскрипции.
9-15
9.2.9. Активный хроматин обладает особыми биохимическими свойствами [29]
Для того чтобы разобраться в строении хроматина, сконденсированного в разной степени, необходимо выделить и охарактеризовать хромосомные белки, присущие каждому из этих состояний. В отношении активного хроматина на этом пути были достигнуты определенные успехи. Установлено, что 1) гистон Н1 не очень тесно связан по крайней мере с некоторой частью активного хроматина; 2) четыре гистона, образующие нуклеосому, содержатся в обычных количествах, но характеризуются необычно высоким уровнем ацетилирования лизиновых остатков, расположенных вблизи аминоконца этих белков. Ацетильные группы присоединяются к ним с помощью фермента гистоновой ацетилазы и удаляются гистоновой деацетилазой, причем каждая ацетильная группа существует в среднем около 10 минут; 3) нуклеосомы в активном хроматине селективно связывают два близкородственных небольших хромосомных белка HMG14 и HMG17. Так как эти белки присутствуют только в активном хроматине, их количество строго соответствует тому, которое требуется для связывания примерно одной из каждой 10 нуклеосом. Аминокислотные последовательности обоих белков крайне консервативны, что свидетельствует о важности их функции; 4) у некоторых организмов, например у Tetrahymena, активный хроматин в значительной мере обогащен минорной формой гистона Н2А. Подобная разновидность гистона найдена также у дрозофилы, курицы и человека. Любое из описанных свойств может играть важную роль в разворачивании хроматина и таким образом способствовать синтезу РНК, однако для подтверждения этой гипотезы требуется проведение прямых экспериментов. Дальнейшие биохимические исследования можно проводить с использованием новых методов очистки активного хроматина на основе селективного связывания с аффинным матриксом, который содержит антитела, специфические либо к белку HMG, либо к ацетилированному лизину. В гл. 10 обсуждаются механизмы контроля образования активного хроматина.
9.2.10. Гетерохроматин сильно конденсирован и транскрипционно неактивен [30]
Доказательства существования второй необычной формы хроматина были получены в 1930 г., когда при наблюдении в световом микроскопе интерфазного ядра клетки высших эукариот была обнаружена особая конденсированная форма хроматина, названная гетерохроматином (менее конденсированная часть хроматина была названа эухроматином). Гетерохроматин в интерфазе остается необычайно компактным, сохраняя такую структуру, которую большая часть хроматина приобретает только в митозе. Позднее обнаружилось, что как и митотический хроматин, Гетерохроматин транскрипционно неактивен (рис. 9-53). Полагают, что в большинстве клеток около 90% хроматина транскрипционно неактивно. Хотя относительная устойчивость такого неактивного хроматина к обработке нуклеазой свидетельствует в пользу того, что этот хроматин более конденсирован, чем 10% в транскрипционно активных областях, лишь часть его (возможно от 10 до 20%) обладает сильно конденсированной конформацией, известной под названием гетерохроматин. Таким образом, гетерохроматин, вероятно, представляет собой особый класс транскрипционно инертного хроматина, несущего определенные функции. У млекопитающих и многих других высших эукариот ДНК, окружающая каждую центромеру, состоит из относительно простых повторяющихся нуклеотидных последовательностей; именно такие «сателлитные ДНК» составляют основную часть гетерохроматина этих организмов.

Рис. 9-53. Радиоавтограф тонкого среза ядра клетки, подвергнутой импульсному мечению [3Н]-уридином для выявления областей активного синтеза РНК (гранулы серебра). Светлые участки - области гетерохроматина, который обычно концентрируется вдоль внутренней поверхности ядерной оболочки. Светлая окраска участков гетерохроматина обусловлена особенностями метода приготовления препарата. Синтез РНК наблюдается главным образом в эухроматиновых участках, окружающих области гетерохроматина. (С любезного разрешения Stan Fakan.)
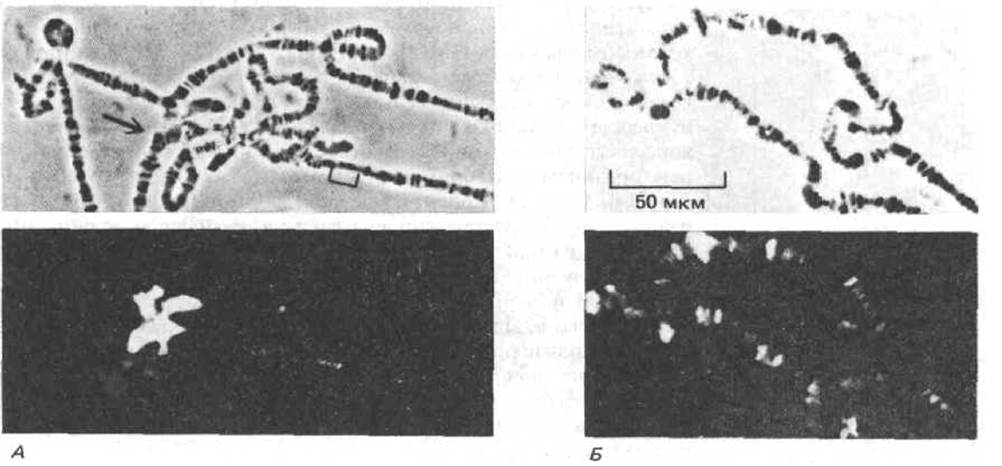
Рис. 9-54. Участок политенной хромосомы дрозофилы в фазово-контрастном (вверху) и флуоресцентном (внизу) микроскопе после иммунофлуоресцентного мечения гетерохроматина (слева) и активного хроматина (справа). А. Окрашивание моноклональными антителами, специфичными к белкам, содержащимся в гетерохроматиновых областях. Окрашены хромоцентр (указан стрелкой) и несколько отдельных дисков. Б. Окрашивание моноклональными антителами, специфичными к белкам, содержащимся в активном хроматине. Окрашивается часть дисков. (Л-по Т.С. James and S.C.R. Elgin, Моl. Cell Biol. 6: 3862 3872, 1986; Я-по G.C. Howard, S.M Abmayr, Z.A. Shinefeld, V.Z. Sato and S.C.R. Elgin, J. Cell Biol. 88: 219-225, 1981. © 1981 Rockefeller University Press.)
У дрозофилы в виде гетерохроматина организованы как хроматин центромеры, так и случайно разбросанные короткие участки. Подобные гетерохроматиновые области недореплицируются на ранних стадиях синтеза ДНК, т. е. в тот период, когда происходит образование политенной хромосомы. Таким образом, эти последовательности в политенных хромосомах представлены в относительно меньших количествах. Биохимические особенности этого типа гетерохроматина можно изучать на молекулярном уровне, проводя связывание с антителами, выявляющими белки хромосом, которые присутствуют только в гетерохроматине (рис. 9-54). Перспективным для биохимического анализа гетерохроматина представляется и клонирование генов, кодирующих специфические для него белки.
Заключение
Все хромосомы во время митоза подвергаются значительной конденсации. Специальное окрашивание выявляет на митотических хромосомах полосы, благодаря которым можно совершенно точно идентифицировать каждую хромосому. Эти полосы, насчитывающие миллионы нуклеотидных пар ДНК, выявляют в хромосоме значительную гетерогенность, природа которой пока еще неясна.
Во время интерфазы хромосомы обычно подвергаются деконденсации в такой степени, что их структура становится трудноразличимой. Замечательным исключением являются специализированные хромосомы типа ламповых щеток в ооцитах позвоночных и политенные хромосомы гигантских секреторных клеток насекомых. Изучение этих двух разновидностей интерфазных хромосом показало, что каждая длинная молекула ДНК, входящая в состав хромосомы, состоит из большого числа дискретных, по-разному упакованных доменов. Основная характерная черта этих хромосом — наличие значительного количества петель хроматина, большая часть которых содержит от 20 000 до 100 000 нуклеотидных пар. Как в хромосомах типа ламповых щеток, так и в политенных хромосомах наименее конденсированными являются области, наиболее активно синтезирующие РНК. Судя по чувствительности к ДНКазе, лишь 10% ДНК находится в относительно деконденсированном состоянии и транскрибируется. Такой «активный» хроматин имеет иные биохимические свойства, нежели те, которыми обладают более конденсированные участки.