Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Цитоскелет
Движение ресничек
После мышечного сокращения наиболее изученным видом клеточной подвижности является биение ресничек. Реснички - это миниатюрные волосовидные образования толщиной около 0,25 мкм, построенные из микротрубочек (микротрубочки - это вторая из трех главных груші нитевидных элементов цитоскелета). Реснички имеются у клеток многих типов и встречаются у большинства животных и некоторых низших растений. Их главная функция - создавать ток жидкости около поверхности клетки или продвигать клетку вперед сквозь толщу воды. Простейшие, например, используют реснички и для передвижения, и для сбора пищевых частиц. У человека огромное множество ресничек (10' и более на 1 см2), принадлежащих клеткам эпителия нижних дыхательных путей, непрерывно перемещает слизь с частицами пыли и остатками отмерших клеток вверх, к ротовой полости, где слизь проглатывается и удаляется. Реснички обеспечивают также передвижение яйцеклетки по яйцеводу, а сходная с ними структура - жгутик -движет сперматозоиды позвоночных.
Подобно тому как изучение мышечного сокращения вносит большой вклад в наше понимание двигательных процессов на основе актина и миозина в немышечных клетках, так и наши знания о механизме биения ресничек помогают понять, каким образом системы микротрубочек порождают движение иного рода - например, обеспечивают внутриклеточный транспорт или процессы, связанные с митозом.
11.3.1. Для ресничек и жгутиков характерны колебательные движения - волны изгиба
Реснички движутся координированно, образуя на поверхности клетки однонаправленные бегущие волны (рис. 11-49). Каждая ресничка работает подобно крошечному хлысту: активный удар вперед, при котором ресничка полностью выпрямляется и преодолевает сопротивление окружающей жидкости, сменяется пассивной фазой, во время которой она возвращается в исходное положение, изгибаясь при этом так, чтобы уменьшить сопротивление среды (рис. 11-50, А). Циклы соседних ресничек едва заметно сдвинуты во времени, что приводит к волнообразной картине, которую можно видеть под микроскопом.

Рис. 11-49, Микрофотографии ресничного эпителия из кишки морского червя, полученные с помощью панирующего электронного микроскопа. Хотя реснички распределены по всей поверхности равномерно, их биение координируется таким образом, что возникают ряды бегущих в одном направлении волн. Эти волны более отчетливо видны при меньшем увеличении справа).
Жгутики спермиев и многих простейших по своей внутренней структуре очень похожи на реснички, но обычно они гораздо длиннее, и характер движения у них иной: не удары хлыста, а бегущие квазисинусоидальные волны (рис. 11-50, Б). Тем не менее молекулярная основа движения здесь одна и та же. Следует, однако, отметить, что жгутики бактерий - это нечто совсем иное, чем реснички и жгутики эукариотических клеток (см. разд. 12.5.4).
11.3.2. Ресничка содержит пучок параллельных микротрубочек, образующих структуру типа 9+2
Биение реснички обусловлено изгибанием ее осевой структуры - так называемой аксонемы. Это сложный комплекс микротрубочек и связанных с ними белков. Микротрубочки обычно представляют собой полые белковые цилиндры с наружным диаметром 25 нм (см. ниже). В аксономе они видоизменены и расположены весьма характерным образом. Открытие этой системы явилось одним из самых впечатляющих результатов ранних электронно-микроскопических исследований: девять сдвоенных трубочек расположены по окружности, в центре которой находятся две одиночные микротрубочки (рис. 11-51). Такая структура типа 9 + 2 характерна для ресничек и жгутиков почти всех эукариотических организмов, от простейших до человека. Микротрубочки тянутся по всей длине аксонемы, которая обычно составляет около 10 мкм, в некоторых клетках может достигать 200 мкм.
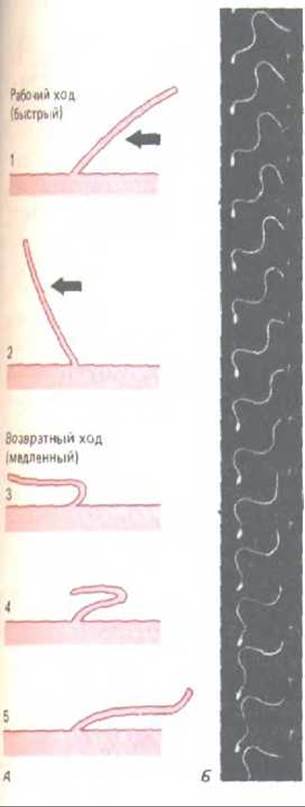
Рис. 11-50. Различный характер биения ресничек и жгутиков. А. Удар реснички (например, реснички эпителия дыхательных путей) напоминает гребное движение пловца. За рабочим ходом (стадии 1 и 2), при котором жидкость прогоняется по поверхности клетки, следует совершенно иной по характеру возвратный ход (стадии 3, 4, 5). Каждый цикл длится обычно 0,1-0,2 с и создает силу, перпендикулярную оси аксонемы. Б. Здесь для сравнения показано волнообразное движение жгутика у спермия оболочника. Клетка была заснята при стробоскопическом освещении с частотой 400 вспышек в 1 с. Обратите внимание, что волны, имеющие постоянную амплитуду, непрерывно движутся от основания жгутика к его концу. В результате клетка движется прямо вперед, как бы отталкиваясь от своей аксонемы, т.е. совершенно иначе, чем в случае с ресничкой. (Б-с любезного разрешения С. J. Brokaw.)
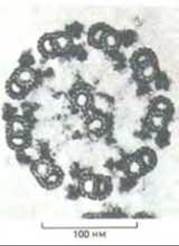
Рис. 11-51. Электронная микрофотография жгутика зеленой водоросли Chlamydomonas (поперечный срез) (с любезного разрешения Lewis Tilney). Видна характерная структура типа 9 + 2, свойственная почти всем ресничкам и жгутикам эукариот. Схема, показывающая основные компоненты этой структуры, представлена на рис. 11-53.
Если микротрубочки центральной пары одинаковы, и каждая представляет собой отдельный полный цилиндр, то наружные дублеты состоят из двух слившихся микротрубочек - одной полной и одной неполной (субфибриллы А и В соответственно). По линии контакта эти субфибриллы имеют участок общей стенки. На поперечных срезах видно, что полная микротрубочка дублета образована кольцом из 13 субъединиц, а неполная (субфибрилла В) - только из 11.
11.3.3. Микротрубочки - полые цилиндры, образованные молекулами тубулина [31]
Микротрубочки состоят из молекул тубулина, каждая из которых представляет собой гетеродимер, образованный двумя прочно связанными глобулярными субъединицами. Эти субъединицы - родственные белки (около 450 аминокислот в каждом), получившие название а- и ß-тубулинов. Хотя тубулин присутствует практически во всех клетках эукариот, главным источником его для биохимических исследований служит головной мозг позвоночных: 10-20% растворимого белка, экстрагируемого из мозга большинством методов, составляет тубулин; и это неудивительно, так как отражает высокое содержание микротрубочек в длинных аксонах и в дендритах нервных клеток.
В ходе сборки микротрубочек молекулы тубулина образуют линейные протофиламенты, в которых а-тубулин одного димера контактирует с ß-тубулином следующего. Целая микротрубочка содержит 13 таких протофиламентов, уложенных параллельно бок о бок вокруг центральной области, которая на электронных микрофотографиях кажется пустой (рис. 11-52). Так как все протофиламенты уложены параллельно и имеют одинаковую ориентацию, микротрубочки, подобно актиновым филаментам, являются полярными структурами, у которых есть плюсконцы, растущие быстро, и минус-концы, растущие медленно (см. схему 11-2). Плюс-концы микротрубочек находятся на кончике реснички.
Подобно актину и многим другим белкам цитоскелета, тубулин у большинства организмов закодирован целым семейством близко родственных генов. У одноклеточной зеленой водоросли Chlamydomonas оказалось два гена а-тубулина и два гена ß-тубулина, а у плодовой мушки Drosophila-no четыре и того и другого. Аминокислотные последовательности тубулинов у разных организмов также очень сходны, хотя они не столь консервативны, как у актина: ß-тубулин дрожжей, например, на 70% идентичен с куриным ß-тубулином, тогда как актины этих видов имеют более 90% идентичных аминокислотных остатков.

Рис. 11-52. А. Электронная микрофотография поперечного среза микротрубочки, где видно кольцо из 13 субъединиц, каждая из которых соответствует отдельной молекуле тубулина. Б. Электронная микрофотография микротрубочки (негативный контраст). В и Г. Схемы строения микротрубочки, показывающие, как молекулы тубулина образуют стенку цилиндра. В-13 молекул на поперечном разрезе микротрубочки; Г - вид сбоку короткого отрезка микротрубочки с уложенными в продольные ряды (протофиламенты) молекулами тубулина. Каждый из 13 протофиламентов состоит из цепи молекул тубулина, представляющих собой aß-гетеродимеры. Микротрубочка-полярная структура; разные концы молекулы тубулина (а и ß) обращены к разным концам микротрубочки. [С любезного разрешения Richard Linck (А) и Rolley Williams (Б); Г-по данным Linda Amos.]
Необычайная эволюционная консервативность актина и тубулина может, по крайней мере отчасти, быть следствием структурных ограничений, которые накладываются связыванием их с многочисленными (и разнообразными) белками. Молекулы тубулина, так же как и актина, взаимодействуют не только между собой, но и со многими вспомогательными белками. Как мы увидим, эти белки модифицируют свойства микротрубочек и соединяют их с другими структурами клетки. По-видимому, большинство случайных мутационных изменений нарушают хотя бы одну из функций микротрубочек или актиновых филаментов и поэтому оказываются вредными для организма.
Все ныне известные тубулины, будучи смешаны in vitro, образуют одинаковые микротрубочки. Тем не менее кажется вероятным, что некоторые вариации в структуре тубулинов имеют для клетки функциональное значение. В частности, у высших позвоночных участки тубулинов обоих типов (а и ß), содержащие необычно много кислых аминокислотных остатков, обнаруживают явные тканеспецифические различия. Эта область в молекулах тубулина, как полагают, участвует в связывании вспомогательных белков, и изменения ее аминокислотной последовательности могут изменять функции микротрубочек, влияя на связывание этих белков.
11.3.4. Вдоль стенки дублета микротрубочек проходит длинная тонкая нить [32]
Хотя большинство микротрубочек состоит, по-видимому, только из субъединиц тубулина, для построения специальных видов микротрубочек (каковы, например, дублеты микротрубочек в ресничке) используются дополнительные белки. Если заставить микротрубочки ресничек или жгутиков диссоциировать в разбавленном солевом растворе, то из такой смеси удается выделить особенно устойчивые фрагменты субфибриллы А-ленты, состоящие из двух-четырех протофиламентов. Помимо тубулина эти фрагменты содержат белок тектин, образующий длинные нити толщиной 2-3 нм, видимо, родственные промежуточным филаментам. Тектиновые филаменты вытянуты вдоль стенки дублета микротрубочек и, вероятно, способствуют образованию общей стенки А- и В-субфибрилл. Как полагают, эти филаменты или какие-то еще не известные нитевидные молекулы определяют расположение на микротрубочках специальных периодических структур, которые будут описаны ниже.
11.3.5. Аксонема ресничек и жгутиков содержит белковые связки, «ручки» и «спицы» [33]
В аксонеме с микротрубочками связано много других белковых структур, взаимодействие которых обеспечивает ресничку энергией и дает возможность использовать эту энергию для волнообразных движений. Пожалуй, самые важные из этих структур - короткие боковые выступы («ручки»), отходящие от каждого дублета микротрубочек внешнего кольца по направлению к соседнему дублету (рис. 11-53). Пары таких выступов располагаются по всей длине субфибриллы А с интервалами в 24 нм. Они состоят из белка, называемого динеином, и, как мы увидим, играют важную роль в движении ресничек и жгутиков. Другой белок - нексин - образует между соседними дублетами поперечные связи, расположенные на несколько большем расстоянии друг от друга, чем динеиновые «ручки»; по-видимому, они весьма эластичны и стягивают аксонему по окружности, ограничивая скольжение соседних микротрубочек.

Рис. 11-53. Схема поперечного среза реснички (соответствующая микрофотографии на рис. 11-51). Вдоль всей реснички с определенной периодичностью расположены различные структуры, отходящие от микротрубочек (см. табл. 11-2).
От каждого наружного дублета внутрь отходит радиальная «спица», доходящая до внутреннего чехла, окружающего центральную пару одиночных микротрубочек (рис. 11-53). Если смотреть на аксонему сбоку, то все эти структуры - динеиновые ручки, нексиновые связки, радиальные «спицы» и отростки центрального чехла - предстанут в виде боковых выступов, повторяющихся с характерной для каждой из этих структур периодичностью (табл. 11-2).
Таблица 11-2. Основные белковые структуры аксонемы
|
Компоненты аксонемы (периодичность расположения вдоль аксонемы) |
Функция |
|
Тубулин (8 нм) Динеиновые ручки (24 нм) |
Главный компонент микротрубочек Выступают из дублетов микротрубочек, взаимодействуют с соседними дублетами, вызывая изгибание |
|
Нексиновые связки (86 нм) |
Удерживают соседние дублеты микротрубочек вместе |
|
Радиальные спицы (29 нм) |
Тянутся от каждого из 9 наружных дублетов по направлению к центральной паре |
|
Выступы внутреннего чехла (14 нм) |
Отходят в виде ряда боковых ручек от центральной пары микротрубочек; вместе с радиальными спицами регулируют характер биения реснички |

Рис. 11-54. Электронная микрофотография изолированной аксонемы (из реснички Tetrahymena), подвергнутой кратковременному воздействию трипсина с целью частично разрушить белковые связи, поддерживающие ее нормальную структуру. После обработки АТР отдельные дублеты микротрубочек скользят относительно друг друга, что ведет к значительному-вплоть до 9-кратного-увеличению длины аксонемы. (F.D. Warner, D.R. Mitchell, J. Cell Biol. 89: 35-44, 1981. С разрешения Rockefeller University Press.)
11-18
11.3.6. Аксонема движется благодаря скольжению микротрубочек [34]
Если жгутик отделить от клетки с помощью лазерного луча, он сохраняет способность производить волнообразные движения. Это означает, что двигательный аппарат находится в самой аксонеме жгутика, а не в его основании (как в случае жгутиков бактерий, см. разд. 12.5.4). Действительно, даже изолированная аксонема может совершать движения в солевом растворе, содержащем АТР.
Изгибание ресничек и жгутиков обусловлено взаимным скольжением микротрубочек. Это было показано в экспериментах, где изолированную аксонему обрабатывали протеолитическими ферментами, которые разрушают нексиновые связки и радиальные спицы, оставляя неповрежденными динеиновые ручки и сами микротрубочки. Если к такой частично «переваренной» аксонеме добавить АТР в концентрации всего лишь 10 мкМ, она начинает удлиняться (причем длина ее может превысить первоначальную в 9 раз); это происходит из-за того, что образующие аксонуму волокна (дублеты) телескопически выдвигаются из ослабленной структуры (рис. 11-54). По-видимому, соседние наружные дублеты активно скользят относительно друг друга, будучи освобождены от скрепляющих боковых сшивок (например, нексиновых). В интактной же структуре это скользящее движение преобразуется в изгиб, что схематически показано на рис. 11-55.
11.3.7. За скольжение ответствен динеин [35]
Если дублеты микротрубочек способны активно скользить друг по другу, то должна существовать сила, вызывающая это движение. Эта сила не может создаваться нексиновыми сшивками, так как после их разрушения путем протеолиза способность к скольжению сохраняется. Между тем динеиновые ручки при протеолизе не разрушаются. Обычно ручки, отходящие от каждого наружного дублета аксонемы, не дотягиваются до соседнего дублета, однако они приходят в соприкосновение с ним, когда ресничка израсходует весь свой запас АТР (рис. 11-56).
Диненн - это крупный белковый комплекс, содержащий две или три (в зависимости от источника) глобулярные головки, соединенные с общим основанием тонкими гибкими нитями (рис. 11-57). Каждая глобулярная головка обладает АТРазной активностью, которая усиливается примерно в шесть раз при ассоциации с микротрубочкой. Вся динеиновая ручка состоит из одной молекулы динеина. Кинетические исследования показывают, что головки, видимо, вызывают скольжение микротрубочек в ресничке с помощью механизма, который в своей основе похож на работу миозиновых головок в мышце (разд. 11.1.10): однонаправленное - от плюс-конца к минус-концудвижение головок динеина по микротрубочке обеспечивается повторными циклами конформационных изменений в каждой головке, обусловленных связыванием и гидролизом АТР. Это движение и создает силу, которая стремится сдвинуть соседний дублет микротрубочек к концу аксонемы (см. рис. 11-55).
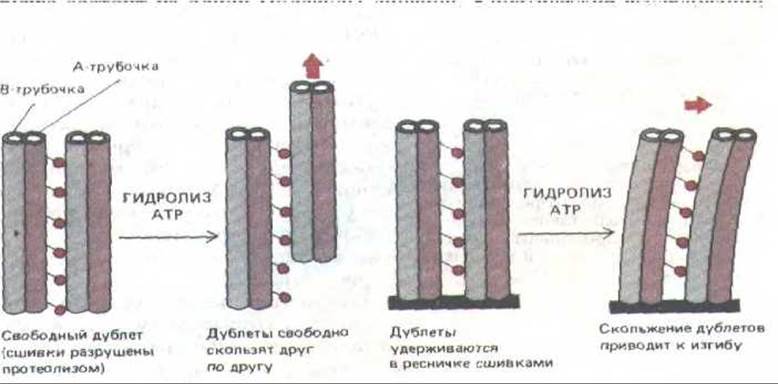
Рис, 11-55. Взаимное скольжение двух наружных дублетов микротрубочек (слева) приводит к изгибу, если дублеты скреплены на одном из концов (справа). Основания молекул динеина соединены только с А-трубочкой, оставляя головки свободными, так что они могут контактировать с соседней В-трубочкой. По-видимому, иная структура В-трубочки мешает связыванию с ней основания молекулы динеина. Такая асимметрия в организации молекул динеина необходима, чтобы предотвратить бесплодное «перетягивание каната» между соседними микротрубочками; возможно, именно поэтому каждая из девяти внешних микротрубочек представляет собой дублет А-В.
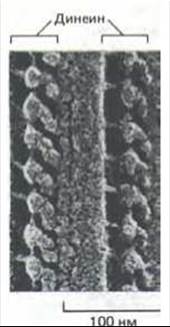
Рис. 11-56. Электронная микрофотография реснички (после замораживания-травления). Видны динеиновые ручки, отходящие от дублета микротрубочек через равные интервалы. (С любезного разрешения John Heuser.)

Рис. 11-57. Динеин - крупный белковый комплекс (мол. масса около 2 млн.), в его состав входят от 9 до 12 полипептидных цепей, самая большая из которых с мол. массой около 450 000. Основание молекулы прочно связано с А-трубочкой (это связывание независимо от АТР), тогда как большие глобулярные головки имеют участки для АТР-зависимого присоединения к другой микротрубочке. Когда головки гидролизуют связанный с ними АТР, они движутся по направлению к минус-концу этой второй микротрубочки, вызывая таким образом относительный продольный сдвиг двух соседних дублетов в ресничке или жгутике (см. рис. 11-55).
11-17
11-19
11.3.8. Скольжение микротрубочек должно регулироваться, чтобы оно могло вызвать изгиб ресничек [36]
Если бы все динеиновые ручки активировались одновременно, как головки миозина в сокращающейся мышце, то аксонема попросту закрутилась бы в тугую спираль. Поэтому для того, чтобы возник локальный изгиб реснички и чтобы этот изгиб распространялся в виде волны от ее основания до самого кончика, должны существовать специальные регуляторные механизмы, координирующие активность молекул динеина. Эта регуляция не может зависеть от потоков Са2+ или других ионов, поскольку аксонема способна изгибаться и после удаления плазматической мембраны. Судя по всему, активация отдельных динеиновых ручек зависит от механических перемещений других компонентов аксонемы, и сигнал передается динеину через белок-белковые взаимодействия. Важную роль играет также упругость жгутика, который в отсутствие активной работы стремится восстановить равновесную конфигурацию всей структуры.
11-20
11.3.9. Аксонему можно изучать генетическими методами [37]
Если выделить аксонемы в очищенном виде и подвергнуть их белки анализу с помощью двумерного электрофореза в полиакриламидном геле (разд. 4.4.5), в них можно обнаружить около 200 различных полипептидов. Изучение функций этих белков и их локализации в аксонеме значительно облегчается при использовании для этой цели мутантних организмов. Излюбленный объект для таких исследований - одноклеточная зеленая водоросль Chlamydomonas reinhardtii, имеющая два жгутика, с помощью которых она плавает (рис. 11-58). Было выделено много мутантов этой водоросли с нарушенной подвижностью. У некоторых из них имеются дефекты в механизме сборки жгутика, и поэтому жгутики не образуются или получаются рудиментарными; у других жгутики есть, но они неподвижны или движутся очень медленно. На электронных микрофотографиях таких мутантных жгутиков можно видеть различные структурные аномалии. У одного класса неподвижных мутантов единственным заметным изменением оказалась потеря динеиновых ручек. У другого класса нет только радиальных спиц, тогда как у третьего отсутствуют одновременно центральная пара микротрубочек и центральная капсула. Изолированные, свободные от мембран аксонемы от мутантов всех трех классов не способны двигаться в присутствии АТР.
Наиболее ценными для понимания работы жгутиков оказались такие мутанты, которые могут двигаться несмотря на утрату некоторых компонентов аксонемы. Выделены, например, медленно плавающие мутанты, у которых нет внутренних или внешних динешювых ручек; следовательно, для создания движущего усилия достаточно присутствия одного-любого - типа динеина, но в норме каждый из них вносит свой вклад в подвижность жгутика. Более неожиданными оказались вторичные мутации, которые восстанавливают подвижность неподвижных жгутиков, лишенных центрального дублета или радиальных спиц, без восстановления исчезнувших компонентов. Существование таких мутантов позволяет думать, что отсутствие центрального дублета или комплекса радиальных спиц каким-то образом выключает активность динеиновых ручек, «замораживая» мотор жгутика, а мутация во втором гене «отключает выключатель» и восстанавливает биение аксонемы. Интересно, что подвижные двойные мутанты с дефектной сердцевиной могут двигаться только по жгутиковому типу и не способны к обычным для Chlamydomonas хлыстообразным ударам, характерным для ресничек. По-видимому, ресничный тип движения требует более сложной структуры, чем волнообразное движение типичного жгутика (см. рис. 11-50).
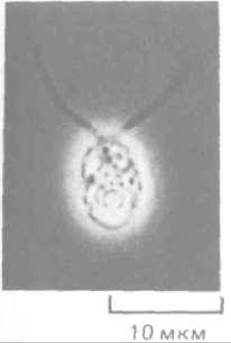
Рис. 11-58. Одноклеточная зеленая водоросль Chlamydomonas reinhardtii. Этот организм плавает с помощью двух жгутиков, которые совместно производят хлыстообразные движения, напоминающие гребок пловца (в отличие от жгутика спермия). (С любезного разрешения John Hopkins.)
Дефекты ресничек и жгутиков встречаются и у человека, например при различных формах наследственного мужского бесплодия, обусловленного неподвижностью спермиев. В зависимости от типа генетической аномалии жгутик спермия может быть лишен динеиновых ручек, головок радиальных спиц или центральной капсулы с одной или обеими центральными микротрубочками. Точно такие же дефекты обнаруживаются в клетках ресничного эпителия этих индивидуумов, и нередко они страдают хроническими респираторными заболеваниями - рецидивирующим бронхитом и хроническим синуситом: из-за нарушенной функции ресничного эпителия у них не происходит надлежащего удаления слизи из бронхов и придаточных пазух носа. Примечательно, что около половины людей, страдающих таким синдромом неподвижных ресничек, имеют также весьма редкую особенность - situs inversus viscerum, т. е. обратную симметрию тела, когда сердце расположено справа, печень и аппендикс слева и т.д. (весь комплекс аномалий носит название синдрома Картагенера). В связи с этим было высказано предположение, что однонаправленное биение ресничек на ранних стадиях развития организма может играть ключевую роль в определении нормальной асимметрии нашего тела.
Рассмотрев движение ресничек и жгутиков, мы перейдем теперь к вопросу о том, как они образуются.
11.3.10. Центриоли выполняют в клетке две различные функции [38]
Можно оторвать у Chlamydomonas пару ее жгутиков так, что центриоли сохранятся; в этом случае жгутики быстро образуются вновь. Почти все необходимые для этого белковые компоненты имеются в растворимой форме в цитоплазме клетки; из них и строятся новые жгутики. Некоторые стадии сборки могут происходить и в бесклеточных экстрактах: молекулы тубулина полимеризуются в микротрубочки (этот процесс мы подробнее рассмотрим в разделе 11.4), а динеиновые ручки могут снова прикрепляться к аксонеме, с которой они были предварительно смыты раствором с высокой ионной силой. Однако сами по себе белки аксонемы не способны восстановить характерную структуру 9 + 2. Для этого необходима «затравка», играющая роль матрицы, на которой происходит дальнейший рост. В клетке такой затравкой служит иентриоль.
Центриоль - это небольшая цилиндрическая органелла толщиной около 0,2 мкм и длиной 0,4 мкм. Стенку центриоли образуют девять групп из трех слившихся микротрубочек (9 триплетов), причем каждый триплет наклонен в сторону центральной оси под углом 45° к огибающей окружности - как лопатки турбины (рис. 11-59). Соседние триплеты центриоль (называемая также базальным тельцем) образует нижнюю часть аксонемы реснички. Справа схематическое изображение центриоли. Центриоль состоит из девяти триплетов микротрубочек, причем каждый триплет (c b a) содержит одну полную микротрубочку и две примыкающие к ней неполные микротрубочки. Особые белки образуют поперечные сшивки, поддерживающие цилиндрическую структуру (выделены цветом). (С любезного разрешения D. Т. Woodrum, R. W. Linck.) соединены между собой через определенные интервалы, а на электронных микрофотографиях часто видны бледные белковые «спицы», идущие к каждому триплету из центральной области, что напоминает колесо телеги (рис. 11-59, А). Нередко центриоли бывают объединены в пары, где они расположены под прямым углом друг к другу (рис. 11-60).
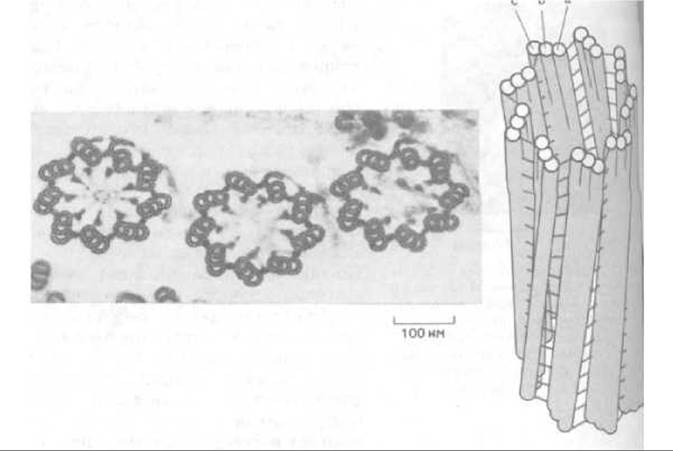
Рис. 11-59. Электронная микрофотография поперечного среза, проходящего через три центриоли в кортексе простейшего.
Центриоль - неизменный компонент аксонемы реснички, и в этом случае ее по традиции называют базальным тельцем. Специальные отростки, называемые исчерченными корешками, соединяют эту центриоль с другими компонентами цитоскелета. При образовании или регенерации каждый дублет микротрубочек аксонемы берет начало от двух из трех микротрубочек триплета центриоли, так что девятилучевая симметрия центриоли сохраняется в строении аксонемы. Данные радиоавтографии указывают на то, что тубулин и другие белки аксонемы пристраиваются к дистальному концу всей структуры, т. е. к плюс-концам микротрубочек. Как в аксонеме возникает центральная пара микротрубочек, не известно; в центриоли этой пары нет.
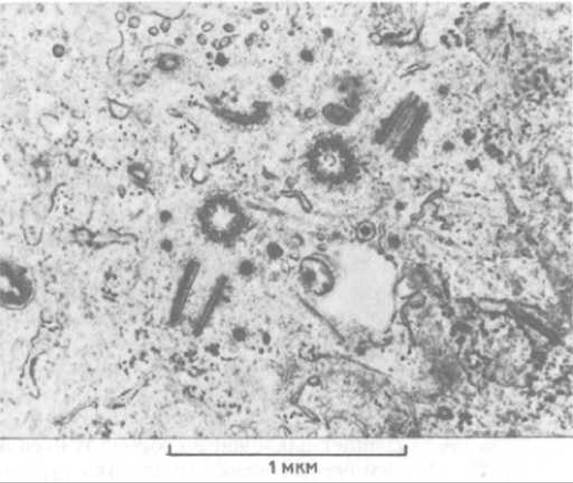
Рис. 11-60. На этой электронной микрофотографии показаны две новые центриоли, которые образовались в результате репликации двух материнских центриолей. На срезе одна центриоль в каждой паре видна в поперечном сечении, а другая в продольном, т.е. в каждой паре центриоли расположены перпендикулярно друг другу. (М. McGill, D. P. Highfield, Т. М. Monahan, R. В. Brinkky. J. Ultrastruct. Res. 57: 43-53, 1976.)
Те центриоли, которые образуют базальные тельца ресничек, выполняют в клетке весьма специализированную функцию, так как реснички сами по себе - структуры специализированные. Наряду с этим почти во всех животных клетках имеется пара центриолей, которая служит как бы срединным элементом центросомы, или клеточного центра. Центросома (разд. 13.5.2) организует цитоплазматические микротрубочки в интерфазных клетках, а в делящихся клетках удваивается и дает начало двум полюсам митотического веретена (мы обсудим это в следующем разделе). Иногда центриоли могут выполнять поочередно то одну функцию, то другую: у Chlamydomonas, например, перед каждым митозом оба жгутика исчезают, а базальные тельца покидают свое место, чтобы стать полюсами веретена.
11.3.11. Новые центриоли обычно возникают путем дупликации уже имеющихся [39]
На фоне непрерывного увеличения массы животной клетки в ходе клеточного цикла выделяются два дискретных акта дупликации: удвоение числа хромосом (репликация ДНК) и удвоение центриолей. В культивируемых фибробластах последний процесс приблизительно совпадает по времени с началом синтеза ДНК. Прежде всего происходит разделение двух «половинок» центриолярной пары, а затем на каждой такой «половинке» достраивается дочерняя центриоль - снова под прямым углом к исходной (рис. 11-60). Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка затем становится матрицей при сборке триплетов, свойственных зрелой центриоли.
У позвоночных ресничные клетки могут нести сотни ресничек, и центриоли клеток-предшественников обеспечивают образование необходимого числа базальных телец. Например, при дифференцировке клеток ресничного эпителия яйцеводов и трахеи пара центриолей перемещается со своего обычного места около ядра в апикальную область клетки, где будут формироваться реснички. Там, вместо того чтобы образовать, как обычно, одну дочернюю центриоль, каждая материнская центриоль дает начало многочисленным электроноплотным «сателлитам». Из этих сателлитов и образуются затем базальные тельца, которые мигрируют к плазматической мембране, чтобы инициировать там рост ресничек.
Однако известны случаи, когда центриоли, по всей видимости, возникают de novo. Так, хотя неоплодотворенные яйцеклетки многих животных лишены функционирующих центриолей и для первого митотического деления (после оплодотворения) используют центриоли спермиев (разд. 15.4.8), в определенных экспериментальных условиях (резкое нарушение ионного баланса или электрическая стимуляция) они могут образовывать различное число центриолей. Каждая такая центриоль инициирует образование небольшой фигуры звезды, и одна из этих звезд может затем использоваться клеткой для деления; при этом из неоплодотворенной яйцеклетки развивается гаплоидный организм (такой ход событий называется партеногенезом - см. разд. 13.4.8). Вероятно, в цитоплазме неоплодотворенных яйцеклеток имеются какие-то предшественники центриолей, которые при особых обстоятельствах могут превращаться в новые настоящие центриоли.
Необычный способ удвоения центриолей и их непрерывность в длинном ряду клеточных поколений заставили в свое время предположить, что центриоли представляют собой полностью автономные, самореплицирующиеся органеллы. Хотя сейчас мы знаем, что это не так и при определенных условиях они могут образовываться в цитоплазме de novo, возможно все же, что какая-то часть информации, необходимая для формирования центриолей, содержится в них самих (подобно тому как размножение митохондрий и хлоропластов зависит от их собственных, внехромосомных генов-см. разд. 7.5). У Chlamydomonas группа генов, которые кодируют белки, участвующие в создании структуры базальных телец и аксонем, находится в дискретном генетическом элементе, и этот элемент передается дочерним клеткам независимо от основных хромосом; его природу и локализацию, однако, еще предстоит выяснить.
Заключение
Реснички и жгутики эукариот содержат цилиндрический пучок из девяти дублетов микротрубочек. Скольжение дублетов относительно друг друга преобразуется в изгиб реснички или жгутика. Силу, сдвигающую дублеты, создают боковые динеиновые ручки, которые тянутся от каждого дублета к соседнему; они используют для этого энергию гидролиза АТР. Ряд вспомогательных белков «увязывает» дублеты в цилиндрическую структуру и ограничивает амплитуду их скольжения. Другие вспомогательные белки образуют своего рода «молекулярна-механическое реле», регулирующее активность динеина таким образом, что изгибание реснички совершается циклически, и это обеспечивает характерное для ресничек биение. Сложная структура аксонемы образуется путем самосборки белковых компонентов, а нуклеацию процесса сборки осуществляет центриоль (базальнеє тельце), которая служит матрицей для формирования специфической структуры аксонемы - системы дублетов микротрубочек типа 9 + 2.