Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Межклеточная сигнализация
Сигнализация с участием внутриклеточных рецепторов: механизмы действия стероидных гормонов
Множество онтогенетических и физиологических процессов у самых разных организмов - от грибов до человека-регулируется небольшим числом стероидных гормонов, синтезируемых из холестерола. Будучи сравнительно небольшими (мол. масса около 300) гидрофобными молекулами, эти гормоны проходят через плазматическую мембрану путем простой диффузии. Оказавшись внутри клетки-мишени, стероидный гормон каждого типа прочно, но обратимо связывается со своим специфическим рецепторным белком. Присоединение гормона ведет к аллостерическому изменению конформации рецепторного белка (процесс, называемый активацией рецептора), что повышает сродство последнего к ДНК; это позволяет рецептору связываться со специфическими генами в ядре и регулировать их транскрипцию. Аналогичным образом действуют гормоны щитовидной железы (тиреоидные гормоны), связываясь со своими рецепторами, которые очень сходны с рецепторами стероидов.
12-10
12.2.1. Комплексы стероидных гормонов с рецепторами присоединяются к специфическим последовательностям ДНК и регулируют транскрипцию генов
Типичная клетка-мишень содержит около 10000 рецепторов для стероидных гормонов. Каждый рецептор может обратимо связать одну молекулу определенного гормона с высоким сродством (константа сродства Ка от 108 до 1010 л/моль). Поскольку рецепторы составляют менее 0,01% общей массы белка в клетке, очистить и охарактеризовать их очень трудно. Недавно, однако, последовательности ДНК, кодирующие некоторые рецепторы стероидных гормонов позвоночных (их кДНК), были клонированы и секвенированы. Оказалось, что рецепторные белки имеют очень сходную структуру. Их полипептидная цепь длиной около 800 аминокислотных остатков образует три отдельных домена: С-концевой домен, который связывает гормон, центральный домен, связывающийся с ДНК, и N-концевой, который активирует транскрипцию гена (рис. 12-9).

Рис. 12-9. Модель белка-рецептора для стероидного гормона. Как полагают, в неактивном состоянии он связан с ингибиторным белком, который блокирует ДНК-связывающий домен рецептора. Связывание гормона рецептором приводит к отделению белка-ингибитора, и в результате рецептор активируется. Прообразом для этой модели послужил рецептор кортизола (глюкокортикоида), но сходную структуру имеют также рецепторы для эстрогенов, тестостерона, прогестерона, альдостерона, тиреоидного гормона, ретиноевой кислоты и витамина D (см. рис. 10-25); вместе все эти белки образуют надсемейство рецепторов стероидных гормонов. В случае рецепторов кортизола и эстрогенов белком-ингибитором служит «белок теплового шока» hsp90 с мол. массой около 90000 (разд. 8.2.7).
Некоторые типы рецепторов стероидных гормонов изначально, в отсутствие гормона, находятся в цитозоле, а другие - в ядре. В обоих случаях присоединение гормона повышает сродство рецептора к ДНК, что позволяет рецептору прочно связываться с определенными нуклеотидными последовательностями в гене, который регулируется данным гормоном. Связывание гормон-рецепторного комплекса со специфическими участками гена активирует (или иногда подавляет) транскрипцию данного гена.
Получить прямое доказательство того, что активированные рецепторы стероидных гормонов связываются со специфическими генами, было очень трудно, и это удалось сделать лишь в 1983г., когда была разработана технология рекомбинантных ДНК. Она позволила клонировать гены, регулируемые стероидными гормонами, и получать в больших количествах специфические последовательности ДНК. Необходимо было еще очистить рецепторные белки, что само по себе является весьма трудоемкой и длительной процедурой. Как только удалось получить рецепторы в очищенном виде, связывающие их последовательности ДНК были картированы in vitro методом футпринтинга (разд. 4.6.6); оказалось, что присоединение рецептора защищает от мягкого расщепления нуклеазами или химическими реагентами группу специфических нуклеотидных последовательностей ДНК. Если эти короткие узнаваемые последовательности из гена удалить, то стероидный гормон уже не будет активировать его транскрипцию. Более того, если короткий фрагмент ДНК, который содержит узнаваемую последовательность, слить с другим геном (репортером) и затем перенести в клетку, содержащую рецепторный белок, то соответствующий стероидный гормон будет активировать транскрипцию гена-репортера. Эти эксперименты показывают, что последовательности ДНК, узнаваемые in vitro активированными рецепторами стероидных гормонов, действительно опосредуют действие рецептора в клетке. Гены, чувствительные к стероидным гормонам, как правило, содержат несколько групп узнаваемых последовательностей, обычно расположенных «выше» (а иногда и «ниже») кодирующей области где-нибудь внутри гена (рис. 12-10). Ввиду значительной структурной гомологии между разнообразными рецепторами для стероидных гормонов близкое сходство распознаваемых ими последовательностей не вызывает удивления.
Однако под прямым влиянием стероидных гормонов находятся лишь немногие гены в любой клетке-мишени. Так, через 30 мин после добавления кортизола к культуре печеночных клеток крысы из тысячи белков, которые можно разделить методом двумерного гель-электрофореза, затронутыми оказались всего семь: количества шести из них увеличились, а количество одного уменьшилось. Эффект был обратимым: после удаления гормона скорости синтеза этих белков возвратились к норме. Как полагают, упомянутым методом можно выявить около 10% клеточных белков; таким образом, присоединение кортизола к 10000 молекул его рецептора в печеночной клетке, вероятно, влияет на транскрипцию лишь около полусотни генов-гораздо меньше, чем следовало ожидать, исходя из числа мест связывания на ДНК. Это означает, что многие активированные рецепторы связываются с ДНК в местах, где их присутствие не дает никакого эффекта.

Рис. 12-10. Локализация последовательностей ДНК в том гене вируса молочной железы, который узнают активированные рецепторы кортизола. Каждая цветная полоска соответствует одному месту связывания рецептора (длиной 8 нуклеотидов). [По К.. R. Yamamoto. In: Transfer and Expression of Eukaryotic Genes (H. Ginsbcig, H. J. Vogel, eds.), pp. 79-92. New York: Academic Press, 1984.]
Как связывание гормон-рецепторного комплекса с геном активирует его транскрипцию? Было показано, что узнаваемые гормоном участки ДНК могут стимулировать транскрипцию даже тогда, когда они удалены на тысячи оснований от промотора, где начинается синтез РНК. Механизм действия таких участков ДНК, называемых транскрипционными энхансерами, обсуждается в гл. 10 (разд. 10.2.11).
12-12
12.2.2. Стероидные гормоны часто вызывают и первичный, и вторичный ответ [7]
Во многих случаях реакция на стероидный гормон бывает двухстадийной. Прямая индукция транскрипции нескольких специфических генов называется первичным ответом. Затем продукты этих генов могут в свою очередь активировать другие гены и вызвать через некоторое время вторичный ответ. Таким образом, простой гормональный пусковой сигнал способен производить весьма сложные изменения в картине экспрессии генов.
Поразительный пример мы находим у плодовой мушки дрозофилы. Уже спустя 5-10 мин после инъекции стероидного гормона линьки-экдизона — в гигантских политенных хромосомах слюнных желез появляются шесть новых участков интенсивного синтеза РНК (они имеют вид пуфов; см. разд. 9.2.6). Через определенное время некоторые из белков, синтезированных во время первичного ответа, индуцируют синтез РНК еще примерно в сотне новых участков, что приводит к появлению большой группы белков, характерных для вторичного ответа. Один или несколько первичных белков управляют всем ответом, выключая по принципу обратной связи транскрипцию всех генов, участвовавших в первичном ответе (рис. 12-11). Вполне вероятно, что аналогичные механизмы осуществляют усиление и регуляцию многих реакций на гормоны и в клетках млекопитающих.
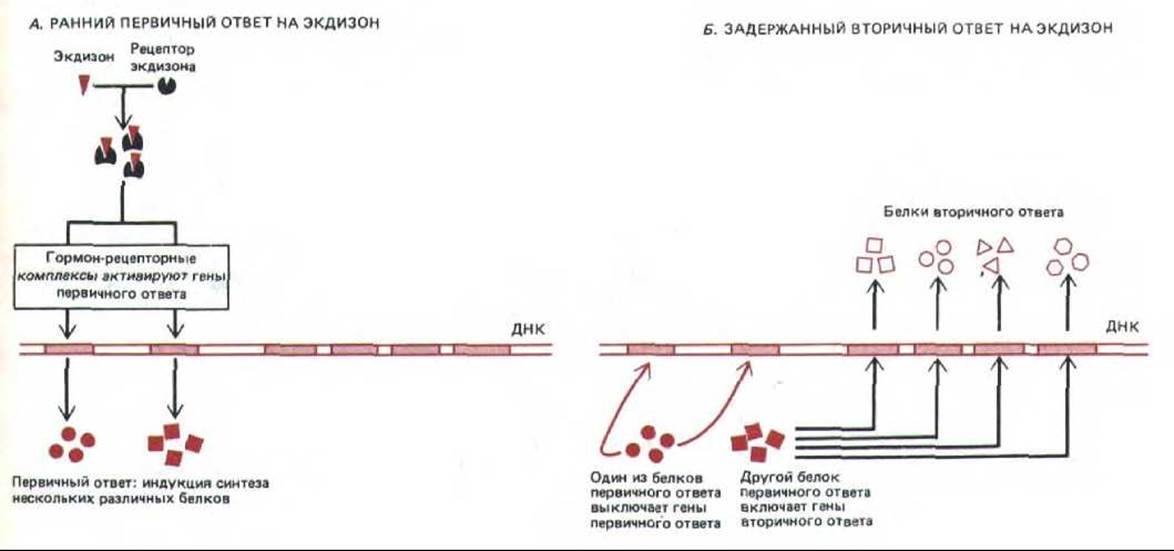
Рис. 12-11. Схема раннего первичного (А) и задержанного вторичного (Б) ответа клеток дрозофилы на экдизон. Некоторые из белков, синтезируемых при первичном ответе, включают гены, ответственные за вторичный ответ, а другие выключают гены, участвовавшие в первичном ответе. На самом деле в обоих ответах участвует больше генов, чем показано на схеме.
12.2.3. В разных клетках-мишенях стероидные гормоны регулируют активность разных генов [8]
Реакция на стероидные гормоны, как и на гормоны вообще, определяется не только природой гормона, но в не меньшей степени и природой клетки-мишени (см. рис. 12-5). В принципе этому может быть два объяснения: либо разные типы клеток имеют разные рецепторы для одного и того же гормона, либо рецепторы одинаковы, но они активируют разные гены. Имеющиеся данные говорят в пользу второго объяснения.
Такие данные были получены в молекулярногенетических экспериментах, показавших, что рецепторные белки для эстрадиола, кортизола и прогестерона кодируются каждый своим собственным единичным геном, и при изучении мутантов млекопитающих с дефектным рецептором мужского полового гормона тестостерона. Все млекопитающие, не подвергшиеся в эмбриональном периоде воздействию тестостерона, развиваются по женскому пути. Мутантные генетические самцы имеют нормальные семенники, вырабатывающие тестостерон, но ткани этих самцов не реагируют на гормон из-за дефектности соответствующих рецепторов. Поэтому у таких самцов развиваются все вторичные половые признаки самок, и семенники их не опускаются в мошонку, а остаются в брюшной полости. Этот синдром тестикулярной феминизации встречается у мышей, крыс, крупного рогатого скота, а также у человека. Хотя изменен только ген, кодирующий рецептор тестостерона, затронутыми оказываются все разнообразные типы клеток, в норме реагирующих на этот гормон (рис. 12-12).
Почему в клетках разного типа один и тот же стероидный гормон активирует разные группы генов? Как описано в гл. 10, для активации эукариотического гена нужно, чтобы с ним, как правило, связалось несколько регуляторних белков (разд. 10.1.5). Поэтому рецептор стероидного гормона может активировать ген только в присутствии необходимой комбинации регуляторных белков, причем некоторые из этих белков тканеспецифичны.

Рис. 12-12. Различные клетки-мишени по-разному реагируют на тестостерон, хотя содержат одинаковые рецепторные белки. При синдроме тестикулярною феминизации мутация в одном-единственном гене, изменяющая рецептор тестостерона, приводит к тому, что все клетки-мишени теряют способность реагировать на этот гормон.
Таким образом, каждый стероидный гормон вызывает характерный комплекс ответов, так как 1) рецепторы для данного гормона имеются только в клетках определенных типов и 2) клетки каждого из этих типов содержат различные комбинации других тканеспецифичных регуляторных белков, которые совместно с активированным стероидным рецептором влияют на транскрипцию специфических групп генов.
Заключение
Стероидные гормоны - это небольшие гидрофобные молекулы, производные холестерола. В крови они находятся в форме водорастворимого комплекса со специальным белком-переносчиком. Освободившись от переносчика, они диффундируют через плазматическую мембрану клетки-мишени и обратимо связываются в цитоплазме или в ядре со специфическими белками-рецепторами. Присоединив к себе гормон, рецептор приобретает повышенное сродство к определенным последовательностям ДНК, которые начинают действовать как энхансеры, т. е. стимулируют транскрипцию нескольких соседних генов. Продукты некоторых из этих генов могут в свою очередь активировать другие гены и вызывать более поздний вторичный ответ, усиливая таким образом действие гормона. Каждый стероидный гормон узнается своим особым рецептором-представителем группы гомологичных белков. Один и тот же рецептор в разных клетках-мишенях регулирует разные наборы генов, вероятно потому, что для транскрипции специфических генов нужны также и другие связывающиеся с ДНК белки, которые в разных клетках различны.