Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Поддержание нормальной организации тканей
Ткани с перманентными клетками
Не все популяции дифференцированных клеток организма подвержены обновлению. Клетки некоторых типов, образовавшиеся в нужном количестве у эмбриона, сохраняются в течение всей взрослой жизни; они никогда не делятся и в случае их утраты не могут быть заменены. В этом смысле перманентны почти все разновидности нервных клеток. Сюда можно отнести и некоторые другие клетки, в том числе у млекопитающих- клетки сердечной мышцы и хрусталика.
Все эти клетки живут чрезвычайно долго и, естественно, находятся в таких местах, где они в норме защищены от повреждающих воздействий, однако в остальном они очень сильно различаются между собой. Трудно указать какую-то одну причину того, что эти клетки - должны быть перманентными, тогда как множество других клеточных популяций обновляется. В случае сердечной мышцы вообще трудно представить себе смысл перманентности клеток. Что касается нейронов (которые будут подробно обсуждаться в гл. 19), то кажется понятным, почему интенсивное обновление этих клеток во взрослом организме нецелесообразно: было бы очень трудно в точности восстанавливать сложную систему нервных связей, созданную в период развития при совершенно иных условиях. Кроме того, следы памяти, записанные в виде небольших изменений структуры или связей определенных нейронов, вероятно, стиралась бы при замене прежних клеток новыми. С другой стороны, перманентность клеток хрусталика - это, по-видимому, простое и неизбежное следствие характера роста его ткани.
17.2.1. Клетки, расположенные у взрослого в центре хрусталика, образовались еще в эмбриональном периоде [6]
Очень немногое во взрослом организме состоит из тех самых молекул, которые были синтезированы у эмбриона. К тем редким структурам, в которых не происходит обновления клеток и даже их внутреннего содержимого, относится хрусталик глаза.
Хрусталик развивается из эктодермы в месте ее контакта с развивающимся глазным пузырем. Здесь эктодерма утолщается и образует впячивание, которое в конце концов отшнуровывается, становясь зачатком хрусталика (см. рис. 17-2). Таким образом, хрусталик закладывается в виде сферического пузырька из одного слоя эпителиальных клеток, окружающих центральную полость. Вскоре часть этого эпителия, расположенная сзади, т. е. обращенная к сетчатке, претерпевает резкое изменение. Ее клетки начинают синтезировать специфические белки хрусталика - кристаллины - и заполняются ими. При этом клетки необычайно удлиняются, дифференцируясь в волокна (см. рис. 17-4). В конце концов их ядра распадаются и синтез белков прекращается. Таким путем часть эпителия хрусталикового пузырька, обращенная к сетчатке, развивается в плотное преломляющее тело, которое состоит из множества высоких призматических клеток, лишенных признаков жизни и уложенных стопками (рис. 17-5). Центральная полость пузырька исчезает, в то время как передняя часть эпителия, обращенная к внешнему миру, сохраняется в виде тонкого слоя низких кубических клеток. Рост хрусталика зависит от пролиферации этих клеток в передней части, откуда они частично выталкиваются на края хрусталика и на его заднюю поверхность (см. рис. 17-4 и 17-5, А). Во время этого передвижения они перестают делиться, начинают синтезировать кристаллины и дифференцируются в волокна хрусталика. Таким путем на протяжении всей жизни в хрусталике появляются дополнительные волокна, хотя скорость их образования постепенно снижается.
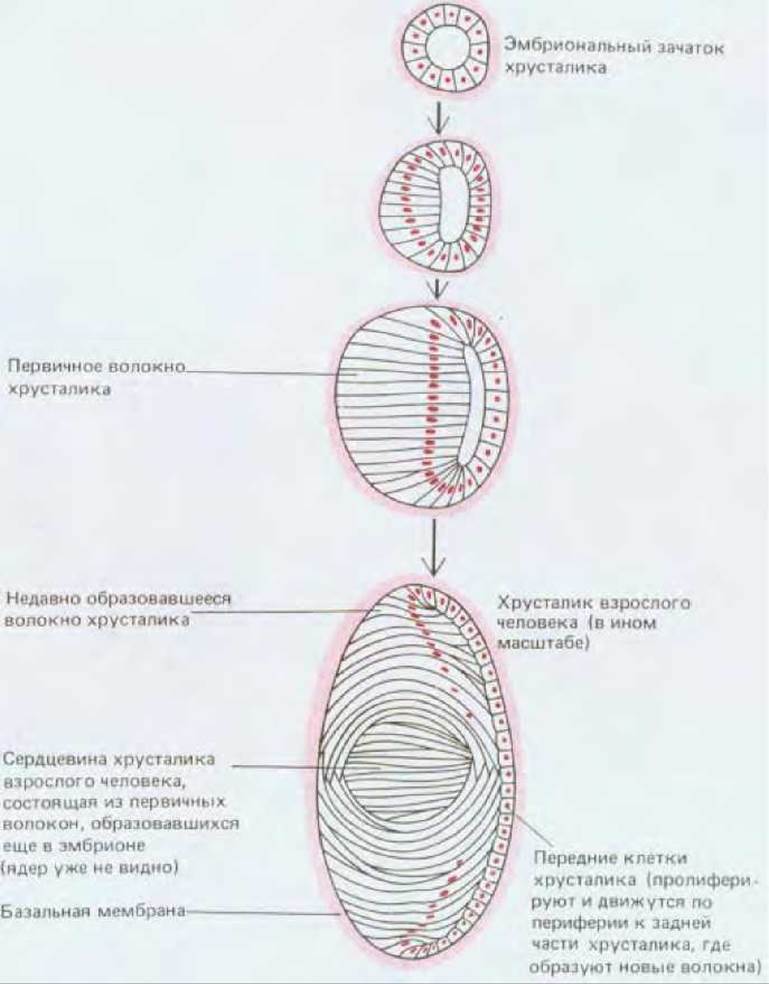
Рис. 17-4. Развитие хрусталика у человека (схематизировано).
Кристаллины в волокнах хрусталика, образовавшихся в ранний период, отличаются от кристаллинов более поздних волокон, подобно тому как гемоглобины в эритроцитах плода отличаются от гемоглобинов взрослого организма. Однако эритроциты заменяются новыми, а волокна хрусталика - нет. Поэтому в сердцевине хрусталика у взрослых находятся волокна, заложенные еще у эмбриона и содержащие кристаллины определенных типов, синтезированные в том раннем периоде. Различия в показателе преломления между ранними эмбриональными типами кристаллинов и более поздними типами помогают избавить хрусталик от оптических аберраций, свойственных простым линзам, сделанным из однородного материала, например из стекла.

Рис. 17-5. Строение хрусталика взрослого человека. А. Световая микрофотография среза части зрелого хрусталика. Показано соединение тонкого эпителия, покрывающего переднюю сторону хрусталика, с дифференцированными волокнами. Б. Микрофотография, полученная с помощью сканирующего электронного микроскопа. Волокна плотно упакованы и напоминают штабеля досок на лесоскладе. Каждое волокно - это одна омертвевшая удлиненная клетка. Длина отдельных волокон достигает 12 мм. (А - с любезного разрешения Peter Gould; Б - из R.G. Kessel, R. H. Kardon, Tissues and Organs: A Text-Atlas of Scanning Electron Microscopy, San Francisco: Freeman, 1979.)
17.2.2. Большинство перманентных клеток обновляет свои составные части. Пример: фоторецепторные клетки сетчатки [7]
Таких неизменяемых клеток, как волокна хрусталика, мало. Как правило, даже клетки, не делящиеся в течение всей жизни организма, обновляют свои компоненты. Клетки сердечной мышцы и нейроны, хотя они и не делятся, метаболически активны и обладают способностью не только синтезировать новые РНК и белки, но и изменять свои размеры и структуру в течение жизни. Например, клетки сердечной мышцы обновляют основную массу своих белковых молекул в течение 1-2 недель, и они могут установить такой баланс между синтезом и распадом белка, что сами клетки увеличатся в размерах, если возрастет нагрузка на сердце - скажем, при длительно повышенном кровяном давлении. Нервные клетки тоже непрерывно заменяют свои белковые молекулы; более того, многие нейроны способны регенерировать перерезанные аксоны и дендриты (см. гл. 19).
Процесс обновления клеточных компонентов особенно ярко можно проиллюстрировать на примере высокоспециализированных нервных клеток, образующих фоторецепторы сетчатки. Нервная часть сетчатки (см. рис. 17-2) состоит из нескольких клеточных слоев, расположенных, казалось бы, весьма странным образом: нейроны, передающие зрительные сигналы в мозг (ганглиозные клетки сетчатки), лежат ближе всего к внешнему миру, так что свет, фокусируемый хрусталиком, должен пройти через них по пути к фоторецепторным клеткам. Последние расположены так, что концы их, воспринимающие свет, - наружные сегменты - частично погружены в пигментный эпителий (рис. 17-6). В соответствии со своей формой фоторецепторы делятся на палочки и колбочки. Они содержат различные светочувствительные комплексы белка со зрительным пигментом. Палочки особенно чувствительны при малой освещенности, тогда как колбочки, представленные тремя разновидностями (каждая для своего участка спектра), служат для восприятия цвета и тонких деталей. Наружный сегмент фоторецептора каждого типа - это, по-видимому, видоизмененная ресничка: в нем мы находим характерное для ресничек расположение микротрубочек в участке, связывающем наружный сегмент с остальной клеткой (рис. 17-7). Главная же часть наружного сегмента почти целиком заполнена плотно уложенными мембранами, в которые погружены светочувствительные белки, связанные со зрительным пигментом. Противоположные концы фото-рецепторных клеток образуют синаптические контакты со вставочными нейронами сетчатки.
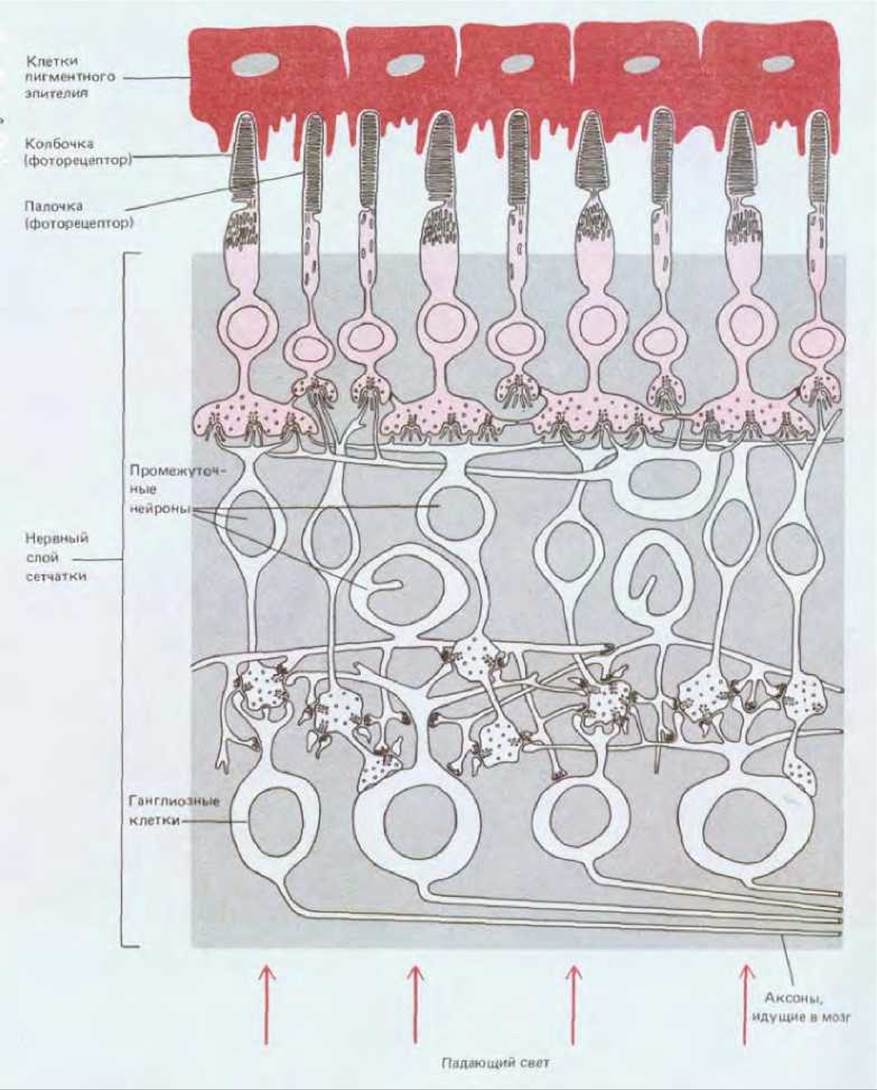
Рис. 17-6. Схема строения сетчатки. При стимуляции фоторецепторов нервные сигналы передаются через промежуточные нейроны ганглиозным клеткам, а те в свою очередь передают их в мозг. Пространство между нейронами и фоторецепторами в нервном слое сетчатки (на рисунке светлосерое) заполнено специализированными опорными клетками, которые на схеме не показаны. (По J. Е. Dowling, В. В. Boycott, Proc. R. Soc. Lond. (Biol.), 166, 80-111, 1966.)
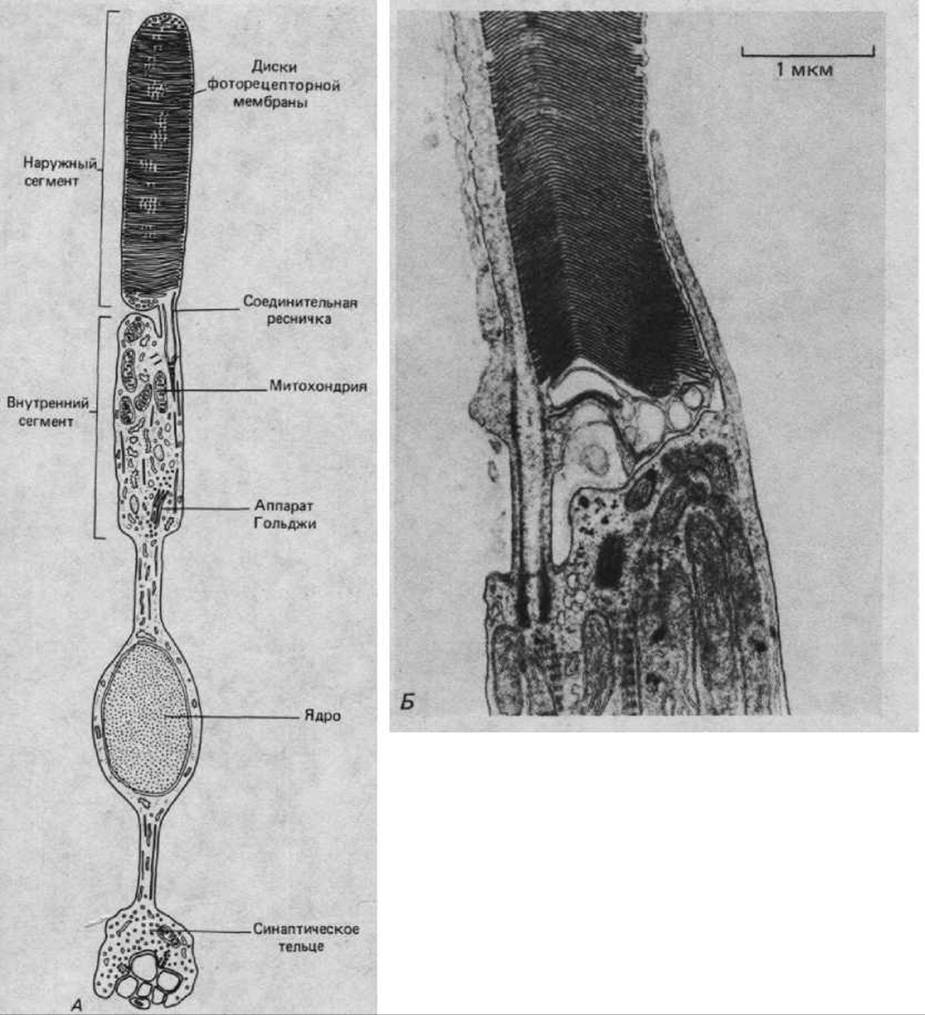
Рис. 17-7. Строение палочки. А. В действительности число фоторецепторных дисков в наружном сегменте достигает примерно тысячи. Б. Электронная микрофотография участка палочки. Можно видеть основание наружного сегмента и видоизмененную ресничку, которая связывает наружный сегмент с внутренним. (A-T.L. Lentz, Cell Fine Structure. Philadelphia: Saunders, 1971; Б-M.J. Hogan, J. A. Alvarado, J. E. Weddell, Histology of the Human Eye: An Atlas and Textbook. Philadelphia: Saunders, 1971.)
Фоторецепторы - это перманентные клетки, не способные делиться. Но молекулы светочувствительного белка не перманентны. Они все время обновляются, и это можно обнаружить по непрерывному включению в них радиоактивных аминокислот. В палочках (любопытно, что этого нет в колбочках) такое обновление идет как на конвейере. В опытах с кратковременным внесением аминокислот можно проследить, как через всю клетку продвигается эшелон меченых белковых молекул (рис. 17-8). После обычных этапов включения аминокислот в белок и упаковки продукта в аппарате Гольджи, происходящих во внутреннем сегменте клетки, радиоактивный материал появляется сначала у основания стопки мембран в наружном сегменте. Отсюда он постепенно перемещается к кончику сегмента, в то время как в основание стопки поступает новый материал. Наконец, после того как меченые белки вместе со слоями мембраны, в которую они погружены, дойдут до вершины стопки (у крысы приблизительно через 10 дней), они фагоцитируются и перевариваются клетками пигментного эпителия.
Дальнейшие сведения о фоторецепторах и их функции в нервной системе читатель найдет в гл. 19.
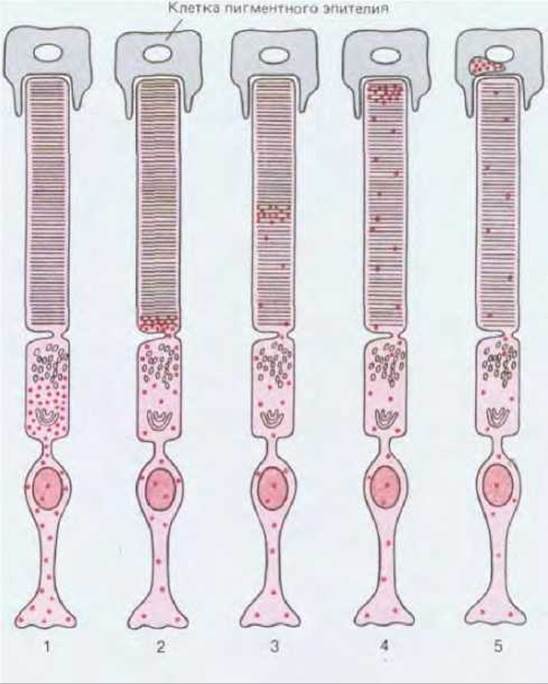
Рис. 17-8. Обновление мембранного белка в палочке сетчатки. После кратковременного введения 3Н-лейцина можно с помощью радиоавтографии следить за перемещением его в клетке. Красные точки - места, где есть радиоактивность. Метод выявляет только лейцин, включившийся в белки; невключившаяся метка отмывается во время приготовления препарата. Включенный лейцин сначала концентрируется по соседству с аппаратом Гольджи (1); отсюда он переходит в основание наружного сегмента и попадает в только что синтезированный диск фоторецепторной мембраны (2). Здесь образуется около 3-4 новых дисков в час (у млекопитающих), и они оттесняют более старые диски в сторону пигментного эпителия (3-5).
Заключение
Нейроны, клетки сердечной мышцы и волокна хрусталика в течение всей жизни организма не делятся и не заменяются новыми. В зрелых волокнах хрусталика клеточные ядра уже дегенерировали и белковый синтез прекратился, так что во внутренней центральной области хрусталика находятся белки, синтезированные еще в раннем эмбриогенезе. Но в большинстве других перманентных клеток метаболическая активность продолжается и идет непрерывное обновление клеточных компонентов. Это четко показано на палочках сетчатки, где новые слои фоточувствительной мембраны синтезируются около ядра, непрерывно перемещаются к верхушке клетки и затем постепенно поглощаются и перевариваются клетками пигментного эпителия.