Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Иммунная система
Как создается разнообразие антител
По имеющимся оценкам, даже в отсутствие антигенной стимуляции у мыши может вырабатываться много миллионов разных молекул антител, - это ее преиммунный репертуар. Поскольку антиген-связывающие участки многих антител могут перекрестно реагировать со множеством близких, но различающихся антигенных детерминант, преиммунный репертуар, видимо, достаточно велик для того, чтобы почти для каждой потенциальной антигенной детерминанты нашелся подходящий антиген-связывающий участок.
Антитела представляют собой белки, а белки кодируются генами. Поэтому разнообразие антител ставит сложную генетическую проблему: каким образом число видов вырабатываемых в организме антител может превышать число генов в его геноме? (Полагают, например, что у человека геном содержит менее 105 генов.) Однако проблема не столь грандиозна, как кажется на первый взгляд. Поскольку в антиген-связывающий участок вносят вклад как L-, так и Н-цепи, у животного с 1000 генов, кодирующих L-цепи, и с 1000 генов, кодирующих Н-цепи, продукты этих генов могли бы образовать 1000 х 1000 различных комбинаций, т.е. 106 разных антиген-связывающих участков (если предположить, что при этом любая L-цепь может сочетаться с любой Н-цепью). Тем не менее иммунная система выработала уникальные генетические механизмы, позволяющие ей генерировать почти неограниченное число различных L- и Н-цепей. Эти механизмы могут варьировать в зависимости от вида животных; например, они весьма различны у кур и у млекопитающих. Ниже мы сосредоточим внимание на механизмах, используемых у млекопитающих.
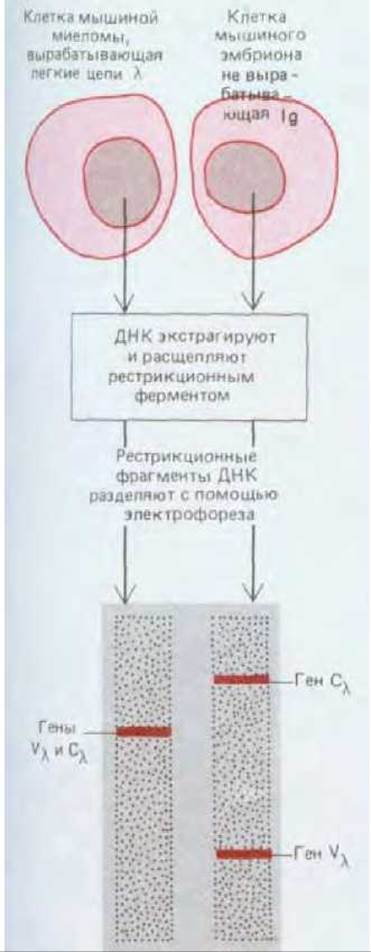
Рис. 18-30. Эксперимент, который прямо показал, что в ходе развития В-клеток ДНК подвергается перестройке. Была экстрагирована ДНК из мышиной опухоли плазматических клеток (миеломы). синтезирующей специфическую легкую цепь Ig, и из 13-дневного мышиного эмбриона, у которого антитела еще не вырабатываются. Ту и другую ДНК расщепляли рестрикционной зндонуклеазой и полученные фрагменты подвергали электрофорезу в агаровом геле. Затем выявляли фрагменты, в которых имелись последовательности, кодирующие С-область L-цепи, и фрагменты с кодом для определенной V-области L-цепи (для этого использовали гибридизацию по Саузерну с двумя радиоактивными ДНК-зондами: один из них был комплементарен кодирующей последовательности для V- области, а другой - для С-области мРНК для специфической L-цепи миеломы) (разд. 18.3). В ДНК клеток миеломы последовательности, кодирующие С - и V-области, были обнаружены в составе одних и тех же фрагментов ДНК, тогда как в ДНК из эмбриона они оказались в разных фрагментах (так же как и в ДНК из другой миеломной опухоли, вырабатывавшей другую L-цепь; на схеме не показано).
18.4.1. В процессе развития В-клеток происходит сборка генов антител из отдельных генных сегментов [20]
Изучение аминокислотных последовательностей миеломных белков привело к предположению о том, что вариабельная (V) и константная (С) области каждой из цепей Ig могут кодироваться двумя отдельными генными сегментами, которые каким-то образом соединяются в ДНК перед их экспрессией. Первые прямые данные о перестройке ДНК в процессе развития В- клеток были получены в 1976 г. при сравнении ДНК из ранних мышиных эмбрионов, неспособных к выработке антител, с ДНК из клеток мышиной миеломной клеточной линии, вырабатывающих антитела. Как показали эксперименты, специфические последовательности, кодирующие V- и С-области и используемые клетками миеломы, находились в этих клетках в одном и том же рестрикционном фрагменте, а у эмбрионов - в двух разных рестрикционных фрагментах. Следовательно, на каком-то этапе дифференцировки В-клеток происходит перестройка последовательностей ДНК, кодирующих молекулы антител (рис. 18-30).
Сейчас известно, что для каждого типа цепей Ig-легких цепей х, легких цепей X и тяжелых цепей - существует отдельный «пул» (обширная группа) генных сегментов, которые могут быть использованы при синтезе отдельной полипептидной цепи. Эти пулы находятся в разных хромосомах, и каждый из них содержит обычно большое число генных сегментов, кодирующих V-область цепи Ig, и несколько меньшее число сегментов, кодирующих С- область. В процессе развития В-клетки происходит - путем сайт-специфической рекомбинации (разд. 5.4.7) - сборка полной кодирующей последовательности для каждой из двух цепей Ig, которые будут синтезироваться; при этом последовательности, кодирующие V- и С-области, соединяются друг с другом. Помимо соединения отдельных сегментов гена, эти перестройки также активируют транскрипцию с генного промотора - возможно, путем изменений относительного положения энхансеров и сайленсеров, воздействующих на промотор (разд. 10.2.7). Поэтому цепь Ig может синтезироваться только после того, как произойдет перестройка ДНК. Процесс объединения генных сегментов увеличивает разнообразие антиген-связывающих участков; здесь играют роль несколько факторов, которые мы рассмотрим ниже.
18.4.2. Каждая V-область кодируется более чем одним генным сегментом [21]
Когда изучили последовательности ДНК, кодирующие V- и С-области, оказалось, что С-область той или иной цепи Ig кодируется лишь одним генным сегментом (С-сегментом), тогда как каждая V-область- более чем одним сегментом. V-область каждой L-цепи кодируется последовательностью ДНК, собираемой из двух генных сегментов - длинного V-сегмента и короткого соединительного J-сегмента (не путать с белковой J-цепью, которая кодируется в другой части генома, - см. разд. 18.2.5). На рис. 18-31 показаны генетические механизмы, участвующие в образовании L-цепей.

Рис. 18-31. Процесс соединения V-J, с которым связано образование æ-легкой цепи у мыши. В ДНК первичных половых клеток (где гены иммуноглобулинов не экспрессируются и поэтому не перестраиваются) группа из четырех J-генных сегментов отделена от С-генного сегмента коротким интроном, а от примерно 300 V-генных сегментов - тысячами нуклеотидных пар. J-сегмент кодирует около 15 С-концевых аминокислот V-области; участок соединения V- и J-сегментов совпадает с третьей гипервариабельной областью. V-сегменты на хромосоме часто бывают собраны в группы гомологичных семейств, которые, по-видимому, распределены по области длиной более 50000 нуклеотидных пар. В ходе развития В-клеток определенный V-сегмент (в представленном здесь случаеV3) перебрасывается точно в положение рядом с одним из J-сегментов (в данном случае J3). «Лишний» J-ген (74) и интрон транскрибируются (вместе с соединенными V3-, J3- и С-сегментами), а затем вырезаются путем РНК-сплайсинга. В результате образуются молекулы мРНК, в которых последовательности V3, J3 и С расположены рядом. Эти мРНК затем
![]()
V-область каждой Н-цепи кодируется последовательностью ДНК, собираемой из трех генных сегментов: V-сегмента, J-сегмента и D-сегмента (diversity gene segment). Организация генных сегментов, участвующих в синтезе Н-цепей, показана на рис. 18-32.
Большое число наследуемых V-, J- и D-сегментов, кодирующих цепи Ig, само по себе вносит существенный вклад в разнообразие антител. Однако этот вклад еще больше увеличивается путем комбинирования разных сегментов (так называемое комбинационное разнообразие).
Например, у мыши каждый из примерно 300 V-сегментов пула для легкой цепи ϰ может соединиться с любым из четырех J-сегментов (см. рис. 18-31), и в результате этот пул может кодировать по меньшей мере 1200 (300 ∙ 4) разных V-областей ϰ -цепи. (Пул для легкой цепи X у мыши содержит лишь два V-сегмента; у человека - значительно больше.) Аналогично, любой из 1000 (или около того) V-сегментов пула для Н-цепи мыши может соединиться с любым из четырех J-сегментов и из по крайней мере 12 D-сегментов, так что могут быть закодированы минимум 48000 (1000-4-12) разных V-областей тяжелой цепи. Эти оценки весьма приблизительны, поскольку точное число V-генных сегментов в этих пулах неизвестно.
Наиболее важный механизм, приводящий к разнообразию антиген-связывающих участков антител, - это комбинационное разнообразие, возникающее при сборке унаследованных V-, J- и D-генных сегментов в различных сочетаниях. Как мы уже видели, у мыши один лишь этот механизм дает возможность вырабатывать по меньшей мере 1000 разных VL- областей и порядка 50000 разных VH-областей. Те и другие затем могли бы образовать 5∙107 комбинаций, т.е. разных антиген-связывающих участков. В дополнение к этому сам по себе механизм соединения, как мы увидим, намного увеличивает число комбинаций - вероятно, более чем в 1000 раз. В результате это число будет значительно больше общего числа В-лимфоцитов у мыши (~108).

Рис. 18-32. У мыши пул генных сегментов для Н-цепи содержит, по-видимому, около 1000 V-сегментов, по меньшей мере 12 D- сегментов, 4 J-сегмента и упорядоченное скопление С-сегментов, каждый из которых кодирует Н-цепь своего класса. D-сегмент кодирует от 1 до 15 аминокислот в третьем гипервариабельном участке V-области. Единый масштаб на рисунке не выдержан, а многие детали опущены. Например:
1) имеются четыре Су-сегмента (Сy1, Сy2a, Су2b) и Сy3);
2) каждый С-сегмент составлен из многих экзонов (см. рис. 18-27);
3) JH1- и Са-сегменты отделены друг от друга примерно 200 000 пар нуклеотидов; так же как и в случае Va-сегментов, VH-сегменты собраны на хромосоме в группы гомологичных семейств. В образовании Н-цепи участвуют те же генетические механизмы, что и в образовании L- цепей (как показано на рис. 18-31), за исключением того, что вместо одной перестройки ДНК необходимы две: сначала D-сегмент соединяется с J-сегментом, а затем к ним присоединяется V-сегмент.
18.4.3. Неточное соединение генных сегментов увеличивает разнообразие V-областей [21]
Генные сегменты, которые могут быть удалены друг от друга на сотни тысяч пар нуклеотидов, соединяются, образуя функциональные последовательности, кодирующие VL- или VH-области. Детали этого механизма неизвестны. К каждому генному сегменту примыкают консервативные последовательности ДНК, которые, вероятно, служат участками узнавания для ферментов сайт-специфической рекомбинации (разд. 5.4.7); благодаря этому рекомбинируются только подходящие друг к другу сегменты. Например, V-сегмент всегда соединяется только с J- или D-сегментом, но не с другим V-сегментом (рис. 18-33).
В большинстве случаев сайт-специфической рекомбинации соединение ДНК бывает точным, но это не относится к соединению генных сегментов для антител. На концах рекомбинирующих сегментов часто теряется различное число нуклеотидов; в случае Н-цепей может также произойти вставка одного или нескольких случайно выбранных нуклеотидов. Такие случайные выпадения или вставки нуклеотидов в местах соединения сегментов в огромной степени увеличивают разнообразие кодирующих V-область последовательностей, образующихся путем рекомбинации, особенно в третьей гипервариабельной области. В данном случае увеличение разнообразия обходится дорого, так как во многих случаях оно приводит к сдвигу рамки считывания и, как следствие, к возникновению нефункционального гена. В процессе развития В-клеток такое «непродуктивное» соединение происходит очень часто.

Судьба промежуточного участка ДНК при соединении двух генных сегментов Ig зависит от ориентации сегментов. Если транскрипционная ориентация этих двух сегментов одинакова, то промежуточная ДНК, как показано на рисунке, освобождается в виде кольца и отбрасывается (делеционное соединение); если же сегменты ориентированы противоположно, эта ДНК сохраняется в перевернутом положении (инверсионное соединение). В биохимическом отношении процессы в обоих случаях одинаковы.
Рис. 18-33. Два типа сигналов для узнавания, используемых при соединении генных сегментов V-области в пулах генных сегментов L- и Н-цепей. А. Локализация сигналов. Для простоты показаны только по два генных сегмента каждого типа. Сигналы обоих типов состоят из консервативных последовательностей (приведенных внизу) длиной в 7 и 9 пар нуклеотидов (п. н.), разделенных неконсервативным спейсером; в одном случае ![]() длина спейсера 21-23 п. н. (примерно два витка двойной спирали ДНК), в другом
длина спейсера 21-23 п. н. (примерно два витка двойной спирали ДНК), в другом ![]()
11-12 п.н. (примерно один виток). Соединяются только подходящие друг к другу генные сегменты: это происходит лишь при сочетании «одновиткового» спейсера с «двухвитковым» (так называемое правило 12/23). Б. Как полагают, сигналы узнавания в свою очередь узнаются ферментом сайт-специфической рекомбинации (рекомбиназой), катализирующим соединение.
18.4.4. Направляемое антигеном соматическое гипермутирование осуществляет тонкую подстройку образования антител [22]
После иммунизации с течением времени обычно происходит постепенное увеличение сродства антител к тому антигену, которым проводилась иммунизация. Этот феномен называют созреванием сродства, и он обусловлен накоплением соматических мутаций в последовательностях, кодирующих V-области, после антигенной стимуляции В-лимфоцитов. Это было наиболее убедительно показано при изучении выработки олигоклональных антител (разд. 18.1.6) у инбредных мышей, у которых V-области L-цепей или Н-цепей кодируются в основном какой-то одной комбинацией генных сегментов. Поскольку изначально последовательности, кодирующие V-область, одинаковы, происходящие в них со временем мутации можно легко обнаружить. В такого рода исследованиях активированные В-клетки, взятые от индивидуальных мышей в разное время после иммунизации, гибридизуют с клетками миеломы, не секретирующими антитела; в результате получают клетки-гибридомы, каждая из которых продуцирует один-единственный вид антител (разд. 4.5.4). Эти бессмертные клетки представляют собой неограниченный источник РНК и ДНК, кодирующих V-области антител. Нуклеиновые кислоты можно затем секвенировать и выявлять изменения в исходных последовательностях, кодирующих V-область. Было показано, что после повторной иммунизации в таких последовательностях со временем быстро накапливаются точечные мутации. Частоту соматических мутаций в этих последовательностях оценивают в 10-3 на одну пару нуклеотидов за одну клеточную генерацию, что примерно в миллион раз больше частоты спонтанного мутирования в других генах. Поэтому такой процесс получил название соматического гипермутирования. Полагают, что этот процесс происходит во время активации В-клеток антигеном с превращением их в клетки памяти, а не при активации с образованием клеток, секретирующих антитела. В-клетки памяти образуются главным образом в лимфоидных фолликулах вторичных лимфоидных органов (см. рис. 18-8).
Поскольку пролиферация В-клеток стимулируется связыванием антигена, любая происходящая в процессе иммунного ответа мутация, увеличивающая сродство мембраносвязанной молекулы антитела, будет вызывать предпочтительную пролиферацию В-клетки, синтезирующей такое антитело, особенно тогда, когда с течением времени после иммунизации концентрация антигена будет снижаться. Таким образом, созревание сродства - это результат повторяющихся циклов соматического гипермутирования, за которыми следует направляемая антигеном селекция в процессе гуморального ответа.
18.4.5. Соединение генных сегментов для антител регулируется таким образом, что обеспечивает моноспецифичность В-клеток [23]
Многочисленные экспериментальные данные показали, что, как и предсказывает теория клональной селекции, каждая отдельная В-клетка вырабатывает антитела с антиген- связывающим участком только одного типа, т.е. В-клетки моноспецифичны.
Моноспецифичность В-клеток может быть важна по меньшей мере по двум причинам. Во-первых, если бы каждая клетка могла вырабатывать более одного вида антиген-связывающих участков, то некоторые из них могли бы продуцировать как полезные антитела, так и антитела к «своим» антигенам (аутоантитела); это затруднило бы отбор на способность реагировать с чужеродными антигенами при одновременном поддержании толерантности к собственным антигенам. Во-вторых, моноспецифичность гарантирует, что все антитела, вырабатываемые одной клеткой, составлены из двух идентичных половинок и поэтому содержат два идентичных антитен-связывающих участка; это позволяет секретируемым антителам образовывать обширные сети из сшитых антигенов, что способствует элиминации антигена (см. рис. 18-13).
Требование моноспецифичности означает, что должен существовать какой-то механизм, благодаря которому при активации генов Ig в процессе развития каждая В-клетка образует только один тип VL-областей и один тип VH-областей. Поскольку В-клетки (как все соматические клетки) диплоидны, каждая из них имеет шесть пулов генных сегментов, кодирующих антитела, два пула для Н-цепей, по одному от каждого из родителей, и четыре
пула для L-цепей, по одному ϰ- и одному λ, от каждого из родителей. Таким образом, перестройки ДНК могли бы в принципе происходить независимо в каждом пуле для Н-цепей и для L-цепей; в результате сборки разных последовательностей, кодирующих V-область в разных пулах, в одной клетке могло бы вырабатываться до восьми различных видов антител, каждое со своим антиген-связывающим участком. В действительности, однако, каждая В-клетка использует лишь два из шести пулов: один из четырех генных пулов для L-цепей и один из двух для Н-цепей (рис. 18-34). Таким образом, каждая В-клетка должна делать выбор
не только между ϰ и λ-пулам и для L-цепей, но также между материнскими и отцовскими генными пулами. Экспрессия только материнского или только отцовского аллеля какого- либо гена в данной клетке называется аллельным исключением; этот феномен, по-видимому, свойствен только генам, кодирующим антитела (и еще генам, кодирующим близко родственные белки Т-клеточных рецепторов, см. разд. 18.6.1). Материнские и отцовские аутосомные гены для других белков экспрессируются в клетке приблизительно одинаково.
Механизмы аллельного исключения и выбора ϰ- или λ-L-цепей в ходе развития В-клеток неясны. Имеющиеся данные позволяют предполагать, что это происходит путем регуляции перестройки ДНК по принципу обратной связи: видимо, функциональная перестройка в одном пуле генных сегментов подавляет перестройки в остальных пулах, кодирующих полипептидную цепь того же типа. Некоторые из таких данных получены в экспериментах, в которых уже собранные ранее клонированные гены инъецировали в ядро оплодотворенной яйцеклетки мыши. В результате получаются трансгенные мыши (разд. 5.6.10), у которых во всех лимфоцитах содержится перестроенный ген. Например, в клонах В-клеток, выделенных из трансгенных мышей с перестроенным геном L-цепи, перестройка эндогенных генов L-цепи обычно оказывается подавленной. Аналогично, введение перестроенного гена μ обычно подавляет перестройку эндогенных генов Н-цепи.
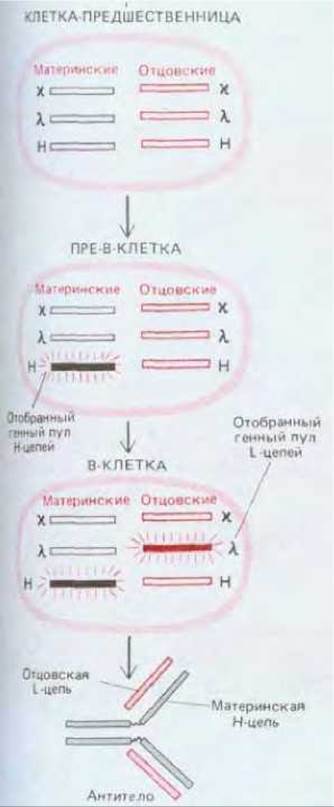
Рис. 18-34. Последовательный выбор активируемых генов Ig, который должен происходить в развивающихся В-клетках, для того чтобы они выработали антитела только с одним типом антиген-связывающего участка.
Клетка должна отобрать один из четырех пулов генных сегментов для L-цепей и один из двух пулов генных сегментов для Н-цепей. В ходе развития в клетке-предшественнице сначала активируется один генный пул Н-цепей, и она становится пре-В-клеткой, вырабатывающей только свободные тяжелые цепи класса μ. После периода интенсивной пролиферации в пре-В-клетке активируется один пул легких цепей г или X, и она становится В-клеткой, вырабатывающей уникальную молекулу IgM.
Сборка последовательностей, кодирующих V-область, в развивающейся В-клетке происходит, по-видимому, в строгом порядке, начиная с пула для Н-цепи и с шагом в один сегмент. Сначала на обеих родительских хромосомах D-сегменты объединяются с JH- сегментами. Затем в одной из этих хромосом происходит соединение VH с DJH. Если такая перестройка дает функциональный ген, последующий синтез полной р-цепи (которая всегда образуется первой из Н-цепей) выключает дальнейшие перестройки генных сегментов, кодирующих Vн-область, и подает сигнал для включения перестроек VL. Сначала происходит соединение VL с JL в ϰ-пуле генных сегментов. Если оно неудачно, перестраивается другой ϰ-пул. Если и эта перестройка оказывается неудачной, происходит объединение в одном, а затем и в другом λ-пуле. Наконец, если все-таки на каком-либо этапе происходит удачное соединение и образуются L-цепи, то они объединяются с предшествующими μ-цепями молекулы антител IgM, которые выключают дальнейшую сборку последовательностей, кодирующих область Vl.
Для того чтобы выключить дальнейшую сборку последовательностей, кодирующих VH, достаточно, видимо, образования интактной μ-цепи. Однако для того, чтобы остановить сборку последовательностей, кодирующих VL, требуется, вероятно, построение полной молекулы антитела. Если развивающейся В-клетке не удается собрать функциональные последовательности, кодирующие как VH-, так и VL-области, она оказывается неспособной вырабатывать молекулы антител и, вероятно, погибает.
Хотя никаких биологических различий между легкими цепями и I не было обнаружено, наличие двух отдельных пулов генных сегментов, кодирующих L-цепи, явно имеет смысл: оно увеличивает вероятность того, что пре-В-клетка, успешно собравшая последовательность, кодирующую VH, успешно соберет также и последовательность для VL и превратится в В-клетку.
18.4.6. Переключение синтеза с мембраносвязанной на секретируемую форму одного и того же антитела происходит путем изменения РНК-транскриптов для Н-цепи [24]
Рассмотрев генетические механизмы, от которых зависит структура антиген-связывающего участка, мы перейдем теперь к механизмам, определяющим биологические свойства антитела, т.е. тип константной области тяжелой цепи. Если сделанный однажды выбор определенных генных сегментов для кодирования антиген-связывающего участка является окончательным и для самой В-клетки, и для ее потомков, то тип синтезируемой СH-области изменяется в процессе развития В-клеток. Изменения могут быть двух типов: переход от мембраносвязанной формы к секретируемой форме одной и той же СH-области и изменение класса этой области.
Антитела всех классов могут синтезироваться как в мембраносвязанной, так и в растворимой, секретируемой форме. Мембраносвязанная форма служит рецептором для антигена на поверхности В-клеток. Растворимая форма вырабатывается только после того, как клетка будет стимулирована антигеном и должна будет секретировать антитела. Две формы антител различаются только С-концевым участком Н-цепи: например, у мембраносвязанных молекул IgM Н-цепь оканчивается гидрофобным участком, закрепляющим ее в липидном бислое плазматической мембраны В-клетки, тогда как у Н-цепей секретируемых молекул IgM имеется вместо этого гидрофильный С-конец, позволяющий молекулам выходить из клетки. Способность В-клетки производить μ-цепи с константными областями двух различных типов сначала казалась парадоксальной, так как В-клетка содержит лишь одну копию генного сегмента Сμ на гаплоидный геном и использует при выработке антител только один из имеющихся двух генных пулов для Н-цепей. Парадокс был разрешен, когда выяснилось, что активация В-клеток антигеном приводит к изменению способа процессинга РНК-транскриптов для μ-цепи в ядре, как показано на рис. 18-35 (см. также разд. 10.4.5). В переключении с мембраносвязанной на секретируемую форму антител других классов участвует сходный механизм.
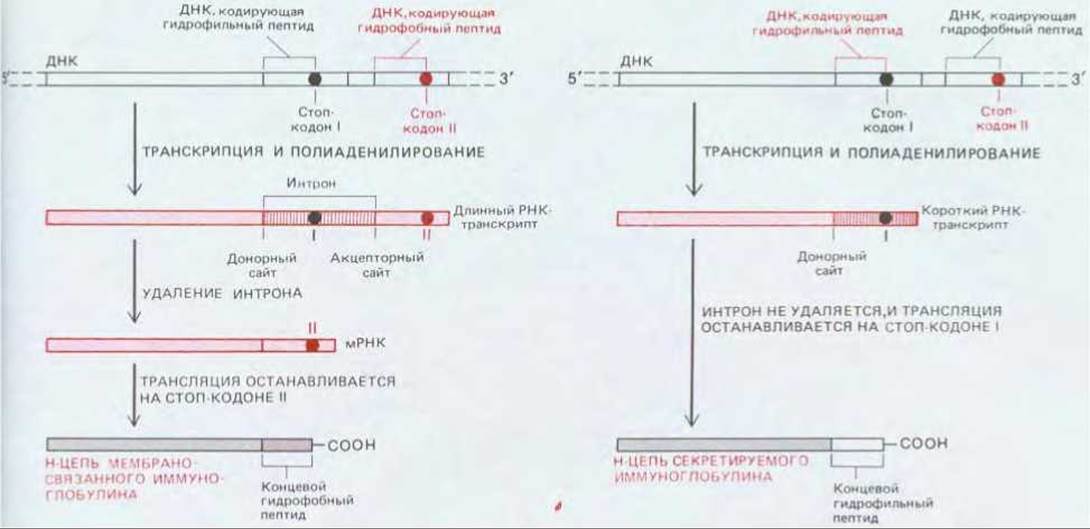
Рис. 18-35. При активации антигеном В-клетка переключается с синтеза антител, связанных с плазматической мембраной, на выработку секретируемой формы тех же антител путем изменения образующихся в клетке мРНК для Н-цепи. Предполагают, что это обусловлено изменением способа расщепления первичных РНК-транскриптов для Н-цепи и их 3'-концевого полиаденилирования. Две формы Н-цепи различаются только С-концевой областью: мембраносвязанная форма имеет гидрофобный «хвост», удерживающий ее в мембране, тогда как у секретируемой формы этот хвост гидрофильный, что позволяет ей выйти из клетки. Длинный полиаденилированный РНК-транскрипт, определяющий мембраносвязанную форму Н-цепи, имеет донорный и акцепторный сайты сплайсинга, что дает возможность удалить последовательность РНК, кодирующую гидрофильный хвост секретируемой формы. В отличие от этого короткий полиаденилированный РНК-транскрипт, определяющий секретируемую форму, имеет только донорный сайт сплайсинга; поэтому последовательность РНК, которая образуется в результате перекрестного разрыва-воссоединения, не может быть удалена.
18.4.7. В-клетки могут переключаться с выработки одного класса антител на выработку другого [25]
В ходе своего развития многие В-клетки переключаются с выработки антител одного класса на выработку антител других классов - процесс, называемый переключением класса. Все В-клетки начинают свою деятельность по синтезу антител с выработки молекул IgM, которые встраиваются в плазматическую мембрану и служат рецепторами для антигена. Затем, еще до взаимодействия с антигеном, большая часть В-клеток переходит к одновременному синтезу молекул IgM и IgD, используемых как мембраносвязанные антигенные рецепторы. При стимуляции антигеном некоторые из этих клеток активируются и начинают выделять антитела IgM, преобладающие в первичном гуморальном ответе (разд. 18.2.5). Другие стимулированные антигеном клетки переключаются на выработку антител классов IgG, IgE или IgA; клетки памяти несут эти антитела на своей поверхности (часто одновременно с IgM), а активные В-клетки их секретируют. Молекулы IgG, IgE и IgA в совокупности называют антителами вторичных классов, так как они, по-видимому, образуются только после антигенной стимуляции и преобладают во вторичных гуморальных ответах.
Класс антител определяется константной областью их Н-цепи (разд. 18.3.3). Поэтому способность В-клеток изменять класс вырабатываемых антител без изменения антиген-связывающего участка означает, что одна и та же собранная последовательность, кодирующая VH-область, может в разное время ассоциироваться с разными генными сегментами для СH. Эта способность В-клеток имеет важный функциональный смысл: благодаря ей определенный антиген-связывающий участок, отобранный антигенами окружающей среды, может у данной особи быть распределен между всеми классами иммуноглобулинов и может, таким образом, приобрести все биологические свойства, характерные для каждого из классов.
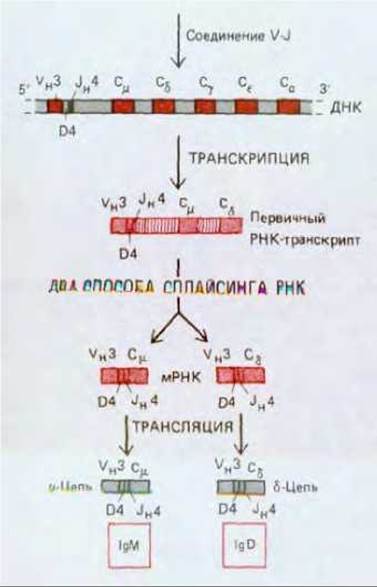
Рис. 18-36. В В-клетках, вырабатывающих одновременно мембраносвячанные антитела IgM и IgD с одними и теми же антигенсвязывающими участками, синтезируются длинные РНК-транскрипты, содержащие как Сμ, так и 05- последовательности. Такие транскрипты подвергаются сплайсингу двумя способами, приводящими к образованию молекул мРНК, у которых одна и та же VH-последовательность соединена либо с Сμ, либо с Сδ-последовательностью. Возможно, что РНК-транскрипты, синтезируемые в таких клетках, еще длиннее и содержат все различные СН-последовательности.
Переключение класса происходит с помощью двух различных молекулярных механизмов. Когда виргильная В-клетка переходит от выработки одного лишь мембраносвязанного IgM к одновременному синтезу мембраносвязанных IgM и IgD, переключение происходит, вероятно, благодаря изменению процессинга РНК. Клетки продуцируют длинные первичные РНК-транскрипты, содержащие наряду с собранной последовательностью VH-области как Сμ-, так и Сδ-последовательности. Затем благодаря альтернативному сплайсингу этих транскриптов образуются молекулы IgM и IgD (рис. 18-36). По-видимому, тот же механизм действует при переключении на другие классы мембраносвязанных lg, когда виргильные В-клетки стимулируются антигеном и созревают в клетки памяти, несущие на своей поверхности IgG, IgE или IgA в качестве рецепторов для антигена.
По-иному происходит терминальное созревание активной В-клетки, секретирующей антитела одного из вторичных классов: оно сопровождается необратимым изменением на уровне ДНК - процессом, который называют рекомбинацией переключения. При этом происходит выпадение всех СH-сегментов «сверху» (т. е. с 5'-стороны по кодирующей цепи) от определенного сегмента, предназначенного для экспрессии в клетке (рис. 18-37). Доказательство того, что этот этап переключения класса связан с делецией ДНК, было получено в опытах с клетками миеломы: оказалось, что клетки, секретирующие IgG, не содержат ДНК, кодирующей Сμ- и Cδ-области, а клетки, секретирующие IgA, не содержат ДНК для С-области Н-цепей всех остальных классов.

Рис. 18-37. Пример перестройки ДНК, происходящей при «переключении» СH-областей. Когда В-клетка, вырабатывающая антитела IgM с участием собранной последовательности VDJ, стимулируется антигеном и созревает в клетку, секретирующую антитела IgA, происходит удаление участка ДНК между последовательностью VDJ и генным сегментом Са. Специфицические участки ДНК (переключающие последовательности, показаны цветными кружками), расположенные «выше» каждого СH-сегмента (за исключением Cδ), рекомбинируют друг с другом, и в результате промежуточный участок ДНК удаляется.
18.4.8. Идиотопы молекул антител служат основой иммунологической сети [26]
Антитела не только защищают организм от инфекций, но и играют важную роль в регуляции самих иммунных ответов. Окончание гуморального ответа на антиген бывает отчасти обусловлено связыванием секретируемых антител с антигеном, который в результате не может присоединяться к рецепторам В-клеток, поэтому стимуляция В-клеток прекращается. Наряду с таким простым ингибированием по типу обратной связи антитела могут участвовать и в более тонком механизме регуляции иммунитета, выступая как часть сложной иммунологической сети.
Иммуноглобулины сами могут действовать как антигены, и можно получить антитела, которые будут узнавать антигенные детерминанты как константных, так и вариабельных участков цепей Ig. Антигенные детерминанты (эпитопы) вариабельных областей L- и Н-цепей, расположенные на антиген-связывающем участке антитела, называются идиотопами (рис. 18-38). Каждый специфический антиген-связывающий участок имеет свой характерный набор идиотопов, поэтому у животного, обладающего миллионами различных антиген-связывающих участков, будут также миллионы различных идиотопов. Поскольку в организме каждый отдельный идиотоп присутствует в очень малом количестве, животное не толерантно к своим собственным идиотопам, и если его надлежащим образом иммунизировать каким-либо из его собственных антител, организм животного будет давать и Т-, и В-клеточные иммунные ответы.
Можно ожидать, что животное, иммунизированное антигеном А, будет сначала вырабатывать большие количества антител к А, затем антитела к идиотопам этих антител к А, а затем в свою очередь антитела к антиидиотопическим антителам и т.д. Действительно, такой тип реакций, дающий «сеть» антител, был продемонстрирован при некоторых иммунных ответах, однако его роль в иммунорегуляции остается неясной.
Как мы увидим позже, рецепторы Т-клеток имеют много общего с мембраносвязанными антителами: они существуют в миллионах форм, каждая из которых имеет свой собственный антиген-связывающий участок и набор идиотопов. По-видимому, в организме животного содержатся как антитела, так и Т-клеточные рецепторы, узнающие большую часть собственных идиотопов организма. При этом в среднем, вероятно, антиген- связывающий участок узнаёт по меньшей мере один идиотоп собственной иммунной системы организма. Таким образом, антиген-связывающие участки иммунной системы потенциально объединены в сложную сеть взаимодействий антиидиотопов (рис. 18-39). Подобно нейронам в нервной системе, многие лимфоциты, возможно, в большей степени взаимодействуют друг с другом, чем с окружающим миром, и тогда иммунный ответ можно было бы рассматривать не как ответ независимых реагирующих с антигеном лимфоцитов, а как реверберирующее возмущение иммунологической сети.
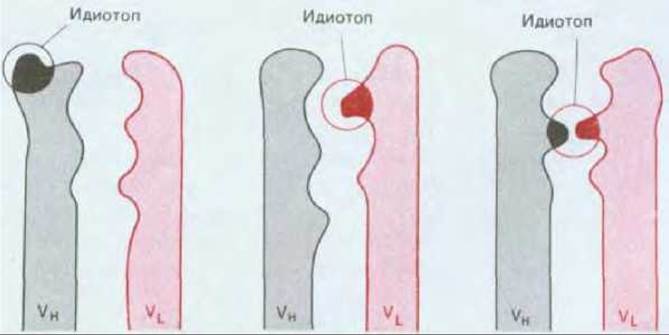
Рис. 18-38. Идиотоп представляет собой антигенную детерминанту молекулы антитела, находящуюся в антиген-связывающем участке или около него. Он может быть образован областью VH, областью VL или обеими областями. Каждый из различных антиген-связывающих участков имеет свой собственный уникальный набор идиотопов, который составляет его идиотип. Идиотопы ассоциированы также и с антиген-связывающими участками Т-клеточных рецепторов.
Заключение
Антитела синтезируются при участии трех пулов генных сегментов, кодирующих соответственно ***-, λ- и Н-цепи. В каждом пуле отдельные генные сегменты, кодирующие разные части вариабельных областей L и и Н-цепей, соединяются путем сайт-специфической рекомбинации в процессе дифференцировки В-клеток. Пулы для L-цепей содержат один или несколько константных (С) генных сегментов и наборы вариабельных (V) и соединительных (J) генных сегментов. Пул для Н-цепей содержит набор С-сегментов и наборы V-, D- и J-сегментов. Для того чтобы синтезировалась молекула антитела, нужно, чтобы VL-сегмент присоединился к JL-сегменту с образованием последовательности ДНК, кодирующей V-область легкой цепи, а VH-сегмент соединился с D- и JH-сегментами с образованием отрезка ДНК, кодирующего V-областъ тяжелой цепи. Каждый из собранных генных сегментов котранскрибируется затем вместе с соответствующей последовательностью С-области, что дает молекулу мРНК, кодирующую всю полипептидную цепь. Комбинируя различным образом унаследованные генные сегменты, кодирующие VL и VH-области, позвоночные могут вырабатывать тысячи различных L-цепей и тысячи различных Н-цепей, которые могут объединяться с образованием миллионов разных антиген-связывающих участков. Это число еще больше увеличивается в результате выпадения и вставки нуклеотидов в процессе соединения генных сегментов и в результат соматических мутаций, происходящих в этих сегментах с очень высокой частотой вслед за антигенной стимуляцией.
Все В-клетки вначале вырабатывают антитела IgM. Некоторые затем образуют антитела других классов, имеющие, однако, тот же антиген-связывающий участок, что и исходные антитела IgM. Такое переключение класса позволяет одним и тем же антиген-связывающий участкам распределяться между антителами с различными биологическими свойствами.