Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Нервная система
Роль ионных каналов в совместной переработке информации нейронами
В центральной нервной системе нейроны обычно принимают сигналы от множества пресинаптических клеток - их число может достигать тысячи или даже нескольких тысяч. Например, на типичном мотонейроне спинного мозга синапсы образуют тысячи нервных окончаний от сотен и, возможно, тысяч различных нейронов; тело нейрона и дендриты почти полностью покрыты синапсами (рис. 19-30). Некоторые из этих синапсов передают сигналы от головного мозга, другие доставляют сенсорную информацию от мышц и кожи, третьи сообщают результаты «вычислений», производимых вставочными нейронами спинного мозга. Мотонейрон должен интегрировать информацию, получаемую из этих многочисленных источников, и либо реагировать, посылая сигналы по аксону, либо оставаться в покое.
Мотонейрон служит типичным примером того, как отдельные нейроны участвуют в фундаментальной задаче вычисления правильного выходного сигнала в ответ на сложную совокупность входных сигналов. Из множества синапсов на мотонейроне одни будут стремиться возбудить его, а другие - затормозить. Хотя все окончания аксона данного мотонейрона выделяют один и тот же медиатор, мотонейрон имеет много различных рецепторных белков, сосредоточенных на разных постсинаптических участках его поверхности. В каждом из таких участков под действием пресинаптических импульсов открывается или закрывается определенная группа каналов, в результате чего в мотонейроне происходит характерное изменение потенциала - возникает постсинаптический потенциал (ПСП). Деполяризация соответствует возбудительному ПСП (возникающему, например, при открытии каналов для ионов натрия), а гиперполяризация - тормозному ПСП (появляющемуся, например, при открытии хлоридных каналов). ПСП, генерируемые разными синапсами на одном и том же нейроне, очень сильно варьируют по величине и по продолжительности. В одном синапсе на мотонейроне пришедший импульс может вызвать деполяризацию менее 0,1 мВ, а в другом - деполяризацию в 5 мВ. Но, как мы увидим, природа системы такова, что даже небольшие ПСП, суммируясь, могут давать большой эффект.
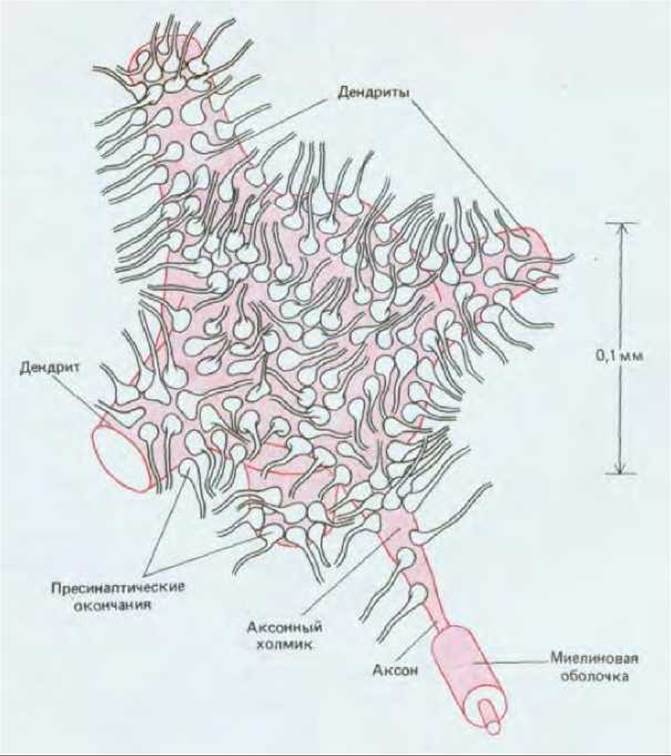
Рис. 19-30. Тело спинномозгового мотонейрона. Показана лишь небольшая часть из многих тысяч нервных окончаний, образующих на клетке синапсы и приносящих сигналы от других частей организма для регулирования ее импульсного разряда. Участки плазматической мембраны мотонейрона, не занятые синаптическими окончаниями, покрыты глиальными клетками (на рисунке не показаны).
19.4.1. Сдвиг мембранного потенциала в теле постсинаптической клетки - это результат пространственной и временной суммации множества постсинаптических потенциалов [25, 26]
Хотя мембрана дендритов и тела большинства нейронов богата рецепторными белками, она содержит очень мало потенциал-зависимых натриевых каналов и поэтому относительно невозбудима. Одиночные ПСП, как правило, не приводят к возникновению потенциала действия. Каждый пришедший сигнал точно отображается величиной градуального ПСП, которая уменьшается по мере удаления от входного синапса. Если сигналы одновременно приходят к синапсам, находящимся на одном и том же участке дендритного дерева, то общий ПСП будет близок к сумме индивидуальных ПСП, причем тормозные ПСП будут учитываться с отрицательным знаком. В то же время суммарное электрическое возмущение, возникшее в одном постсинаптическом участке, будет распространяться на другие участки благодаря пассивным кабельным свойствам мембраны дендрита.
Тело нейрона, где сходятся все эффекты от ПСП, обычно невелико (менее 100 мкм в диаметре) по сравнению с дендритным деревом, длина ветвей которого может измеряться миллиметрами. Поэтому мембранный потенциал тела клетки и ближайших к нему частей отростков будет примерно одинаков - это будет совокупный результат эффектов, производимых всеми входными сигналами с учетом удаленности того или иного синапса от тела нейрона. Таким образом, можно сказать, что суммарный постсинаптический потенциал тела клетки - это результат пространственной суммации всех полученных стимулов. Если преобладают возбуждающие входные сигналы, то тело клетки деполяризуется, если тормозные - обычно гиперполяризуется.
В то время как пространственная суммация интегрирует эффекты сигналов, принятых разными участками мембраны, временная суммация объединяет сигналы, поступившие в разное время. Нейромедиатор, высвобождающийся после прибытия потенциала действия к синапсу, создает на постсинаптической мембране ПСП, который быстро достигает пика (благодаря кратковременному открытию лиганд-зависимых ионных каналов) и затем экспоненциально (что определяется емкостью мембраны) снижается до исходного уровня. Если второй импульс придет до того, как первый ПСП полностью затухнет, то этот второй ПСП суммируется с оставшимся «хвостом» первого. Когда после некоторого периода покоя приходит длинный залп быстро повторяющихся импульсов, каждый последующий ПСП будет накладываться на предыдущий, давая в итоге большой ПСП, величина которого отражает частоту разряда пресинаптического нейрона (рис. 19-31). Таким образом, суть временной суммации состоит в том, что частота получаемых сигналов преобразуется в величину суммарного ПСП.
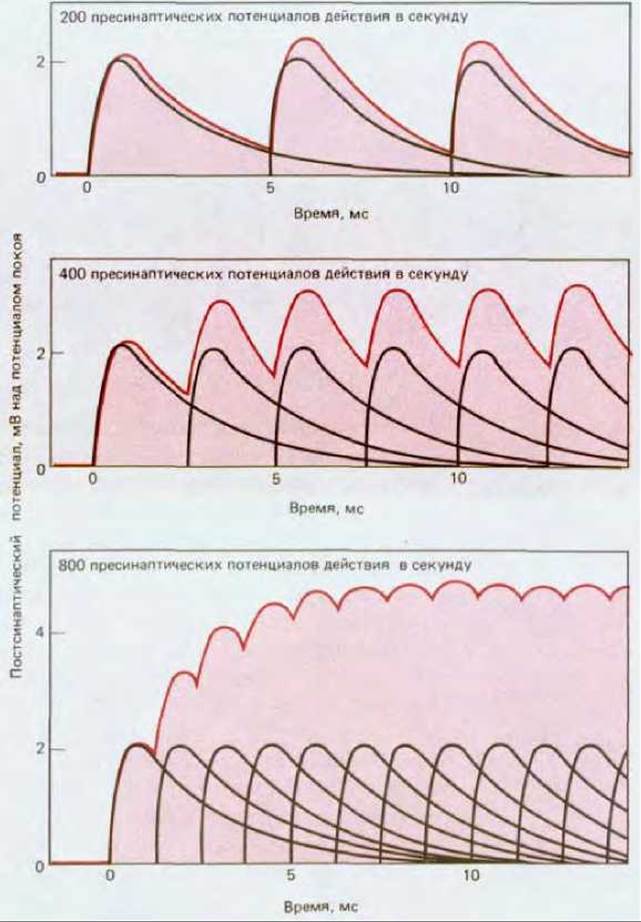
Рис. 19-31. Временная суммация. Перекрывающиеся черные кривые накрашенной области показывают индивидуальный вклад каждого из последовательных пресинаптических импульсов в суммарный постсинаптический потенциал.
19.4.2. Для передачи информации на большие расстояния суммарный ПСП снова преобразуется в частоту нервных импульсов [27]
Благодаря временной и пространственной суммации мембранный потенциал тела одного постсинаптического нейрона регулируется частотой разрядов множества пресинаптических нейронов. В результате интеграции всех входных сигналов постсинаптическая клетка формирует определенный ответ, обычно в виде импульсов для передачи сигналов другим клеткам, нередко находящимся в отдаленных частях организма. Этот ответный сигнал отражает величину суммарного ПСП в теле клетки. Однако, хотя суммарный ПСП все время плавно изменяется, потенциалы действия имеют постоянную амплитуду и подчиняются закону «всё или ничего». Единственной переменной величиной при передаче сигналов с помощью импульсов остается временной интервал между последовательными импульсами. Поэтому для передачи информации на большие расстояния величина суммарного ПСП должна быть преобразована, или перекодирована, в частоту импульсного разряда (рис. 19-32) Такое кодирование достигается с помощью специальной группы потен циал-зависимых ионных каналов, сосредоточенных у основания аксона в области, называемой аксонным холмиком (см. рис. 19-30).
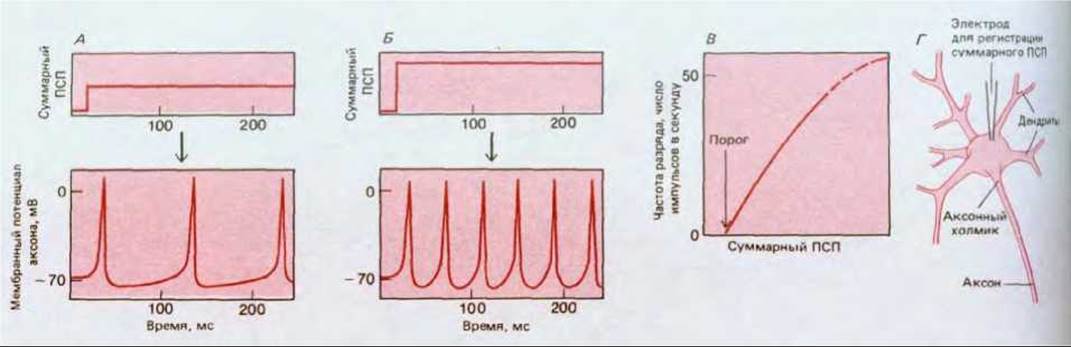
Рис. 19-32. Перекодирование суммарного ПСП в частоту импульсного разряда в аксоне. Из графиков А и Б видно, как частота импульсов в аксоне возрастает с увеличением суммарного ПСП; на графике В представлена общая зависимость. Г-способ измерения суммарного ПСП. А и Б: на верхних графиках - суммарная интенсивность синаптических стимулов, получаемых телом клетки; на нижних графиках соответствующие разряды, посылаемые по аксону. На верхних графиках показано, как выглядели бы ПСП, если бы импульсный разряд был каким-то образом блокирован.
Прежде чем объяснить, как действуют такие каналы, необходимо сделать некоторые уточнения. Само по себе возникновение импульса приводит к резким изменениям мембранного потенциала всего тела клетки, который уже не будет прямо отражать суммарную синаптическую стимуляцию, получаемую клеткой. Поэтому очень трудно провести точный анализ кодирующего механизма. В последующем чисто качественном описании мы будем употреблять выражения «сила синаптической стимуляции» или «суммарный ПСП, имея в виду тот суммарный ПСП, который создавался бы при условии, что генерирование импульсов каким-то образом подавляется; и мы будем предполагать что именно этот основной суммарный ПСП и является причиной возникновения нервных импульсов.
19.4.3. Кодирование требует совместного действия различных ионных каналов [28]
Проведение нервных импульсов зависит главным образом, а во многих аксонах позвоночных почти полностью, от потенциал-зависимых натриевых каналов. Первоначально импульсы генерируются мембраной аксонного холмика, где таких каналов очень много. Но для осуществления особой функции кодирования мембрана аксонного холмика должна содержать еще по меньшей мере четыре класса ионных каналов - три избирательно проницаемых для ионов калия и один проницаемый для Са2+. Три разновидности калиевых каналов обладают различными свойствами - мы будем называть их медленными, быстрыми и Са2+ - зависимыми калиевыми каналами. Кодирующие функции этих канала наиболее изучены на гигантских нейронах моллюсков, но те же принципы используются, по-видимому, и в других нейронах.
Чтобы понять, для чего нужны каналы нескольких типов, посмотрим, как будет вести себя мембрана нервной клетки, содержащая только один вид потенциал-зависимых каналов - натриевые каналы. При слабой синаптической стимуляции, не доводящей деполяризацию мембраны аксонного холмика до порогового уровня, потенциал действия не буди возникать. При постепенном усилении стимуляции порог будет достигнут, натриевые каналы откроются и возникнет потенциал действия.
В результате последующей инактивации натриевых каналов появившийся потенциал действия исчезнет. Прежде чем сможет возникнуть другой потенциал действия, натриевые каналы должны будут выйти из инактивированного состояния. Но для этого необходимо, чтобы величина мембранного потенциала вновь достигла большого отрицательного значения, а этого не произойдет, пока поддерживается сильный деполяризующий стимул (от ПСП). Поэтому для того, чтобы реполяризовать мембрану после импульса и подготовить клетку для проведения следующего импульса, нужен еще один вид каналов. Эту функцию выполняют медленные калиевые каналы, которые уже упоминались, когда речь шла о распространении потенциала действия (разд. 19.2.2). Эти каналы зависимы от потенциала и открываются при деполяризации мембраны так же, как и натриевые, но происходит это с некоторым запаздыванием. Открываясь во время спада потенциала действия, калиевые каналы пропускают ионы К+ из клетки наружу, в результате чего устраняется эффект даже длительного деполяризующего стимула и мембранный потенциал возвращается к уровню равновесного калиевого потенциала. Этот потенциал настолько отрицателен, что натриевые каналы выводятся из состояния инактивации. Кроме того, прекращается и выход калия из клетки: реполяризация мембраны приводит к тому, что медленные калиевые каналы опять закрываются (так и не успев инактивироваться). Как только произошла реполяризация, деполяризующие стимулы от синаптических входов могут вновь поднять мембранный потенциал до порогового уровня и вызвать очередной потенциал действия. Благодаря этому непрерывная стимуляция дендритов и тела клетки приводит к многократно повторяющемуся возбуждению аксона.
Однако недостаточно только непрерывности разряда - нужно еще, чтобы его частота отражала интенсивность стимуляции. Детальные расчеты показывают, что простая система натриевых и медленных калиевых каналов не отвечает этому требованию. Если сила постоянного стимула ниже определенного порогового уровня, потенциалов действия не будет вовсе; если же сила стимуляции превысит порог, то сразу же начнется частая импульсация. Проблему решают быстрые калиевые каналы (называемые также А-каналами). Эти каналы тоже потенциалзависимы и открываются при деполяризации мембраны, но специфическая зависимость их от потенциала и кинетика инактивации такова, что они снижают частоту разряда при уровнях стимуляции, которые лишь ненамного выше порога. Таким образом, быстрые калиевые каналы помогают устранить разрыв непрерывности в соотношении между интенсивностью стимула и частотой разряда; в результате частота импульсов пропорциональна силе деполяризующего стимула в очень широком диапазоне (см. рис. 19-32).
19.4.4. Адаптация уменьшает реакцию на постоянный стимул [29]
Обычно процесс кодирования видоизменяют еще два типа каналов, имеющихся в аксонном холмике. О них уже упоминалось - это потенциал-зависимые кальциевые каналы и Са2+-зависимые калиевые каналы. Первые подобны кальциевым каналам, участвующим в высвобождении медиатора из окончаний аксона: в области аксонного холмика эти каналы открываются при возникновении потенциала действия и пропускают Са2+ внутрь аксона. Са2+-зависимые калиевые каналы отличаются от всех других каналов, описанных ранее. Они открываются при повышении концентрации кальция у внутренней поверхности мембраны нервной клетки.
Предположим, что сильный и продолжительный деполяризующий стимул приводит к длительной импульсации. В результате каждого импульса в клетку через потенциал-зависимые кальциевые каналы переходит небольшое количество ионов Са2+, так что их внутриклеточная концентрация постепенно поднимается до высокого уровня. Это ведет к открытию Са2+-зависимых калиевых каналов, и проницаемость мембраны для калия повышается, что затрудняет деполяризацию и увеличивает интервалы между последовательными импульсами. Таким образом, при длительном воздействии постоянного стимула сила ответа нейрона постепенно снижается. Это явление, в основе которого могут лежать и другие механизмы, называют адаптацией (рис. 19-33). Благодаря адаптации нейрон, так же как и нервная система в целом, способен с высокой чувствительностью реагировать на изменение стимула, даже если оно происходит на фоне сильной постоянной стимуляции (разд. 19.6.8). Это одно из общих приспособлений, благодаря которым мы, например, не замечаем постоянного давления одежды на наше тело, но в то же время быстро реагируем на внезапное прикосновение.
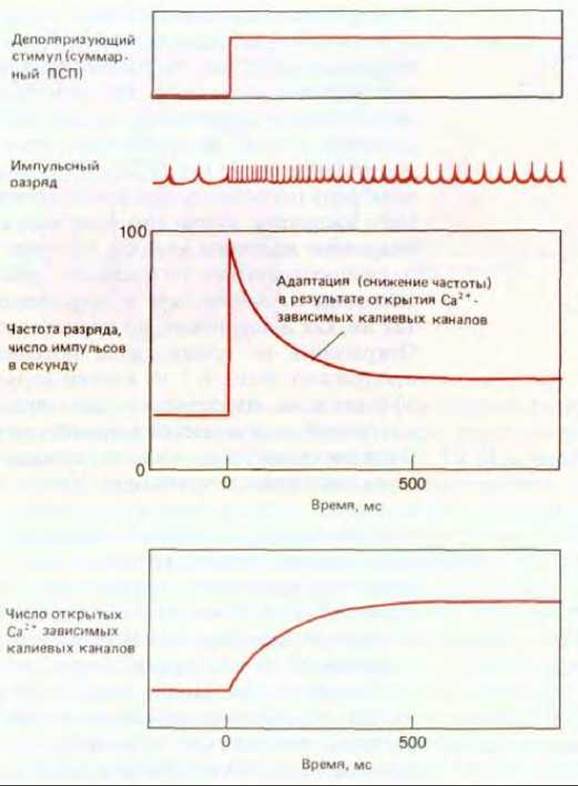
Рис. 19-33. Адаптация. При длительной стимуляции постоянной силы реакция клетки на стимул постепенно ослабевает, что выражается в уменьшении частоты импульсного разряда.
19.4.5. Сигналы могут передаваться не только по аксонам [30]
В типичном нейроне, о котором говорилось выше, дендриты и аксон резко различаются между собой по строению и функции. Однако некоторые нейроны не соответствуют такой схеме, хотя молекулярные основы их функционирования те же. Например, у большинства беспозвоночных нейроны чаще всего имеют униполярную организацию: тело клетки связано одним-единственным «стебельком» с системой ветвящихся клеточных отростков, среди которых не всегда можно отличить дендриты от аксона (рис. 19-34). Функциональные отличия тоже мот быть «смазаны», что встречается и у позвоночных, и у беспозвоночных: отростки, которые по их строению можно отнести к дендритам, часто образуют как пре-, так и постсинаптические структуры и способны как принимать сигналы, так и передавать их другим клеткам. И наоборот, входные синаптические сигналы иногда воспринимаются «стратегическими» участками аксона - например, вблизи окончания, где эти сигналы могут усиливать или тормозить высвобождение нейромедиатора из данного окончания, не влияя на передачу в окончаниях других ветвей того же аксона (рис. 19-35). Пример этого важного механизма пресинаптического торможения и пресинаптического облегчения будет рассмотрен позже (разд. 19.5.4).
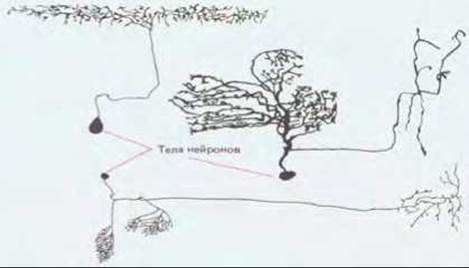
Рис. 19-34. Нейроны мухи. Строе-кие их типично для большинства нейронов беспозвоночных животных; тело клетки соединено с системой отростков при помощи стебелька, так что нет дендритов, отходящих прямо от тела. Сходной организацией обладают и сенсорные нейроны из спинномозговых ганглиев позвоночных. (N. Strausfield, Atlas of an Insect Brain, New York, Springer, 1976.)
Синапсы, через которые дендриты передают стимулы другой клетке, играют важную роль в коммуникации между нейронами, расположенными друг от друга на расстоянии нескольких миллиметров или еще ближе. На такие расстояния электрические сигналы могут передаваться по дендриту пассивно от постсинаптического участка, где они были восприняты, до пресинаптического участка, где они регулируют высвобождение медиатора. Встречаются даже нейроны, совсем не имеющие аксона, не проводящие потенциалов действия и передающие все сигналы через дендриты. Более того, если дендритное дерево велико, то отдельные части его могут более или менее независимо использоваться для связи и обработки информации. Диапазон возможностей некоторых нейронов расширяется еще больше благодаря наличию в мембране дендритов потенциал-зависимых каналов, что позволяет дендритам проводить потенциалы действия. Таким образом, даже отдельный нейрон способен функционировать как очень сложное вычислительное устройство.
Заключение
Дендриты и тело типичного нейрона принимают множество различных возбуждающих и тормозных синаптических сигналов, которые подвергаются пространственной и временной суммации и создают суммарный постсинаптический потенциал тела клетки. Для передачи сигналов на большие расстояния величина этого потенциала преобразуется в частоту импульсного разряда при помощи системы ионных каналов в мембране аксонного холмика. Механизму такого кодирования часто свойственна способность к адаптации, и тогда клетка слабо реагирует на постоянный стимул, но чувствительна ко всякому изменению его силы. Существует много вариантов описанной общей схемы; например, не все нейроны дают ответ в виде потенциалов действия, дендриты могут быть не только постсинаптическими, но и пресинаптическими, а аксон - не только пресинаптическим, но и постсинаптическим.
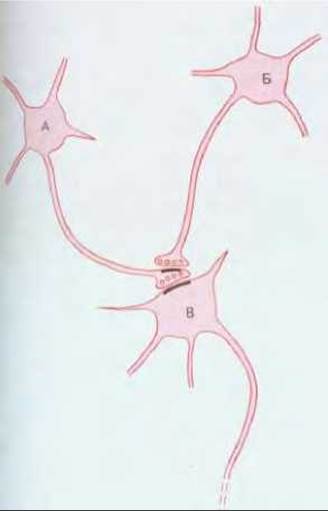
Рис. 19-35. Аксоаксонный синапс. Нейромедиатор, выделяемый окончанием аксона клетки В, воздействует на каналы в окончании аксона клетки А, изменяя тем самым число квантов нейромедиатора, поступающих на клетку В при возбуждении Б. Если возбуждение Б ослабляет силу стимуляции В клеткой А, то говорят, что Б осуществляет пресинаптическое торможение. Противоположный эффект называют пресинаптическим облегчением.