Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Нервная система
Рождение, рост и гибель нейронов
Проблема развития нервной системы уникальна. Каким образом аксоны и дендриты, отходящие от миллиардов нейронов, отыскивают надлежащих партнеров для связей так, чтобы создалась эффективно функционирующая сеть? Различного рода нейроны и сенсорные клетки, а также интернируемые мышцы чаще всего находятся у зародыша на значительных расстояниях друг от друга и первоначально не связаны между собой. Поэтому в первой фазе развития нервной системы различные ее части развиваются по собственным «локальным» программам в соответствии с принципами дифференцировки клеток, общими для всех тканей тела, о чем уже говорилось в гл. 16. В следующей фазе осуществляется тип морфогенеза, свойственный только нервной системе. На этом этапе создается хотя и предварительная, но уже упорядоченная схема связей между частями нервной системы с помощью аксонов и дендритов, растущих в нужных направлениях. Первоначально обособленные части могут теперь взаимодействовать друг с другом. В третьей, последней фазе, продолжающейся и в период взрослой жизни, возникшие ранее связи уточняются и совершенствуются в результате взаимодействия отдаленных компонентов с учетом электрических сигналов, передаваемых и получаемых этими компонентами.
19.7.1. Нейроны образуются в соответствии с определенными программами клеточного деления [48, 49]
Почти у всех животных от нематод до позвоночных образование нейронов, за редкими исключениями, подчиняется трем основным правилам: 1) зрелые нейроны не делятся; 2) после образования полного комплекта нейронов, свойственного взрослой особи, не остается никаких стволовых клеток, способных производить новые нейроны; 3) нейроны в каждой небольшой области развивающейся нервной системы формируются в соответствии с собственной программой деления, без влияния со стороны тех групп клеток, с которыми позднее образуются нервные связи.
У позвоночных нервная система развивается из двух клеточных комплексов эктодермального происхождения - нервной трубки (разд. 16.1.9) и нервного гребня (разд. 16.6,5). Из нервной трубки развивается центральная нервная система (головной и спинной мозг), тогда как нервный гребень служит источником большинства нейронов и поддерживающих клеток периферической нервной системы. Помимо этого из утолщений, или плакод, эктодермы головы развиваются некоторые сенсорные нейроны, в том числе иннервирующие ухо и нос (рис. 19-55).
Нервная трубка, о которой здесь в основном пойдет речь, вначале состоит из однослойного эпителия - в дальнейшем источника образования как нейронов, так и глиальных клеток центральной нервной системы (рис. 19-56). Позже этот эпителий утолщается и становится более сложным образованием со многими слоями клеток различного типа. Процесс пролиферации здесь был изучен с помощью 3Н-тимидина, который включается в клетки, находящиеся в S-фазе клеточного цикла. Ткань фиксировали либо сразу после введения метки, чтобы установить, какие клетки делятся, либо на более поздней стадии, чтобы узнать, какие зрелые клетки образуются из тех клеток-предшественников, которые делились в момент введения 3Н-тимидина. Такие исследования особенно успешно используются для определения «дат рождения» различных нейронов: так как нейроны, прошедшие дифференцировку, не делятся, каждый предшественник нейрона должен пройти свое последнее деление в определенный день перед началом своего созревания как нервной клетки. Оказалось, что как у позвоночных, так и у беспозвоночных «даты рождения» нейронов данного типа обычно приходятся на тот строго ограниченный период развития, после которого уже не образуется нейронов этого типа. Каждая область развивающейся нервной трубки имеет собственную программу деления клеток, и обычно нейроны, имеющие разные «даты рождения», предназначены для разных функций. Так как после завершения образования нервных клеток и начала формирования связей между НИМИ стволовые клетки прекращают свое существование, число нейронов в дальнейшем может меняться только в сторону уменьшения в результате отмирания клеток (см. ниже разд. 19.7.11).
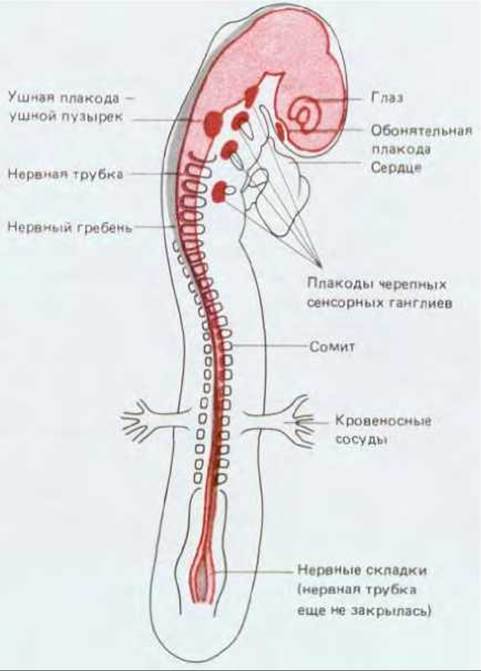
Рис. 19-55. Схематическое изображение раннего (2,5 дня) куриного зародыша: показано, из каких структур образуется нервная система. Нервная трубка (выделена розовым цветом) уже закрылась, за исключением хвостового участка, и располагается под эктодермой, частью которой она была вначале (см. рис. 16-13). Нервный гребень (показан серым цветом) расположен вверху между крышей нервной трубки и эктодермой. Из утолщений на поверхности эктодермы - плакод - развиваются определенные группы сенсорных клеток и нейронов. На этой стадии уже практически завершилась инвагинация статоакустической плакоды и образовался слуховой пузырек - зачаток внутреннего уха и источник нейронов связанного с ним ганглия; в результате инвагинации обонятельной плакоды образуется выстилка носа, включая обонятельные нейроны, ответственные за восприятие запахов. Другие черепные плакоды дадут начало клеткам черепных сенсорных ганглиев, которые обеспечивают большую часть сенсорной иннервации головы и шеи. В отличие от других сенсорных клеток сенсорные клетки глаза развиваются из нервной трубки.
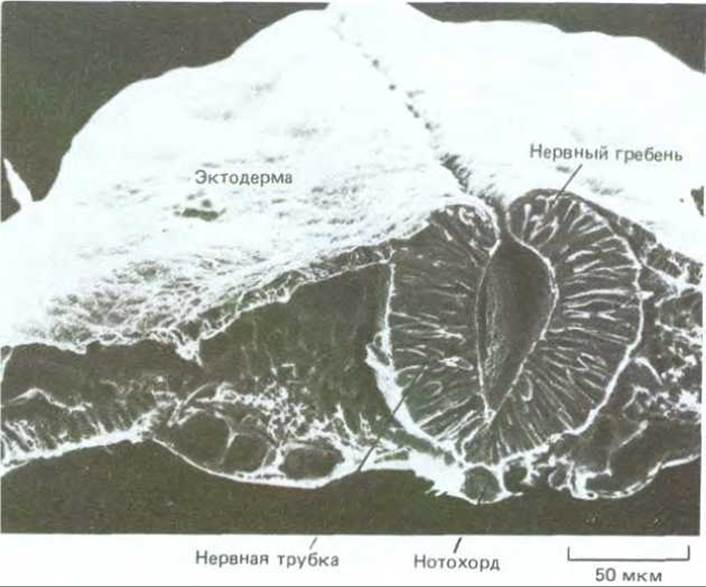
Рис. 19-56. Микрофотография поперечного среза двухдневного куриного зародыша, полученная с помощью сканирующего электронного микроскопа. Нервная трубка уже почти замкнулась и отделилась от эктодермы. На этом этапе эпителий нервной трубки состоит из одного клеточного слоя. (С любезного разрешения Jean-Paul Revel.)
19.7.2. Радиальные глиальные клетки образуют временный «каркас», направляющий миграцию незрелых нейронов [50]
Незрелый нейрон еще не образовавший аксона и дендритов, обычно мигрирует из места своего «рождения» в какой-нибудь другой участок. Такие миграции нейронов можно прослеживать с помощью 3Н-тимидина: метка включается в клетки-предшественники, проходящие последнее деление, после чего их меченые потомки могут быть обнаружены в других местах. Например, мотонейроны, которые будут иннервировать конечности, проходят последний митоз около просвета нервной трубки, а затем перемещаются к ее периферии и попадают в вентральные рога будущего спинного мозга (рис. 19-57).
Миграцию нейронов направляют специализированные элементы нервной трубки - радиальные глиальные клетки (рис. 19-58, А). Это сохранившиеся клетки первоначального столбчатого эпителия нервной трубки, которые все больше вытягивались, по мере того как стенка трубки утолщалась: каждая из этих клеток простирается от внутренней поверхности трубки до наружной. В некоторых участках развивающегося головного мозга приматов это расстояние может достигать 2 см. Трехмерная реконструкция по электронным микрофотографиям серийных срезов показывает, что незрелые мигрирующие нейроны тесно примыкают к радиальным глиальным клеткам и, видимо, как бы ползут по ним (рис. 19-58, Б и В).

Рис. 19-57. Участки, из которых развиваются мотонейроны спинного мозга, но данным ридиоавтографии после введения небольшой дозы тимидина, меченного тритием, на раннем этапе развития. На схемах показаны поперечные разрезы нервной трубки раннего эмбриона (слева) и относительно зрелого спинного мозга, развивающегося из нее (справа). На поздней стадии наибольшее количество метки содержат те клетки, которые в момент введения 3Н-тимидина проходили свою последнюю фазу синтеза ДНК. Для простоты на схеме зрелого спинного мозга показаны только мотонейроны, хотя в сером веществе имеется также много других нервных клеток. Белое вещество (не закрашено) содержит главным образом пучки аксонов, идущих вдоль спинного мозга и связывающих разные области серого вещества. (У взрослых особей эти области выглядят белыми, так как содержат большое количество миелина.) О том, как образуются глиальные клетки в процессе развития организма, см. в гл. 16.

Рис. 19-58. А. Поперечный срез части развивающейся коры головного мозга обезьяны (упрощенная схема). На препарате, окрашенном по методу Гольджи, видны радиальные глиальные клетки, отходящие от просвета нервной трубки к ее наружной поверхности. Б. Более подробная схема области, выделенной розовым цветом на схеме А. Показаны незрелые нервные клетки, мигрирующие вдоль отростков радиальных глиальных клеток. В. Изображение одного из таких мигрирующих нейронов, основанное на реконструкциях по серийным электронно-микроскопическим срезам. (По P. Rakic, J. Сотр. Neurol., 145, 61-84, 1972.)
Радиальные глиальные клетки сохраняются в течение многих дней (у некоторых видов до нескольких месяцев) как популяция неделящихся клеток, ясно отличающихся от нейронов и их предшественников. Только к концу периода развития они в большинстве областей головного и спинного мозга исчезают; высказано предположение, что многие из них превращаются в астроциты, но это еще требует прямых доказательств. Таким образом, радиальные глиальные клетки можно рассматривать как вспомогательный аппарат развития - они необходимы в качестве «лесов» при построении сложных нервных структур, но в зрелой нервной системе почти нигде не сохраняются.
19.7.3. Тип нейрона и его будущие связи определяются временем его «рождения» [50, 51]
Существует определенная связь между «датой рождения» нейрона в центральной нервной системе млекопитающих и местом его окончательной локализации (возможно, это эволюционный «отголосок» жесткой связи между генеалогией клеток в развивающемся организме и конечной локализацией их у таких беспозвоночных, как нематоды - см. разд. 16.3). Например, в коре головного мозга нейроны располагаются слоями в соответствии с последовательностью их «рождения» благодаря такой миграции, при которой клетки, образовавшиеся позднее, мигрируют дальше клеток, образовавшихся раньше. По мере созревания клетки, расположенные в следующих друг за другом слоях коры, начинают различаться по форме, размерам и характеру связей с другими клетками. Так, малые пирамидные клетки, появляющиеся поздно, расположены в одном из наружных слоев и посылают свои аксоны в другие области коры, тогда как большие пирамидные нейроны и клетки неправильной формы, появившиеся ранее, находятся во внутренних слоях и посылают аксоны за пределы коры большого мозга.
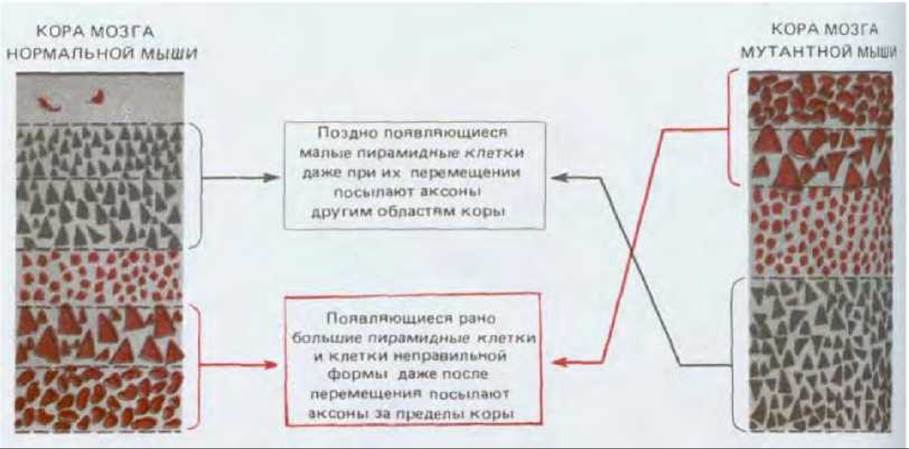
Рис. 19-59. Сравнение нейронных слоев в коре головного мозга нормальных мышей и мутантных мышей reeler. У мутантов нарушение миграции клеток приводит к инверсии нормальной взаимосвязи между «датой рождения» нейрона и его конечным положением. Тем не менее нейроны с измененным положением дифференцируются и устанавливают связи в соответствии с «датой рождения».
От чего зависят такие различия - от «даты рождения» или от места окончательной локализации? Ответить на этот вопрос помогают мыши мутантной линии reeler. У этих мутантов, названных так за нетвердую походку, нарушен механизм миграции нервных клеток: нейроны, образовавшиеся позднее, остаются во внутреннем слое, а ранние клетки переходят в наружный. Но, несмотря на инверсию в их расположении, дифференцировка кортикальных клеток соответствует времени их «рождения», т. е. клетки, образовавшиеся позже, становятся малыми пирамидными нейронами, а ранние клетки - большими пирамидными или же нейронами неправильной формы. Следовательно, в данном случае тип клетки определяется именно временем ее образования, а не окончательным местом (рис. 19-59). По-видимому, особенности нейронов и в самом деле обычно зависят главным образом от их происхождения и от места и времени образования.
В свою очередь присущие клетке особенности определяют характер ее будущих связей - это важное общее правило, которое будет подробнее рассмотрение позже (разд. 19.7.9). Поэтому у мышей reeler нейроны, изменившие местоположение, устанавливают связи (с незначительным числом ошибок) в соответствии с «датой рождения», а не с положением в коре мозга: аксоны малых пирамидных клеток направляются в другие участки коры, а аксоны больших пирамидных клеток и клеток неправильной формы - в области, лежащие за пределами коры. Для того чтобы понять, как устанавливаются такие избирательные связи, нужно будет сначала рассмотреть механизм образования аксонов и дендритов.
19.7.4. Аксоны и дендриты удлиняются благодаря конусу роста на их кончиках [52]
Как правило, аксон, а затем и дендриты начинают расти от тела нервной клетки вскоре после того, как нейроны попадают на свои окончательные места. Всю последовательность событий впервые наблюдали в интактной эмбриональной ткани, применяя окрашивание по Гольджи (рис. 19-60). Эта методика и другие методы, разработанные позднее, позволили выявить на конце растущего отростка нервной клетки своеобразное утолщение неправильной формы. Эта структура, называемая конусом роста, видимо, и прокладывает путь через окружающую ткань. Конус роста служит одновременно и «локомотивом», и приспособлением, направляющим отросток по надлежащему пути.
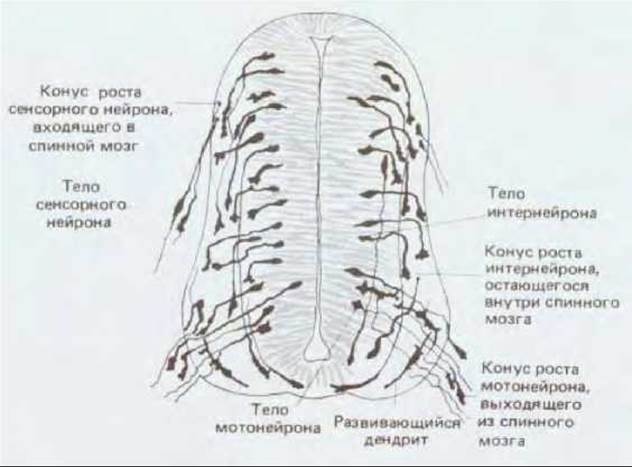
Рис. 19-60. Конусы роста в развивающемся спинном мозгу трехдневного куриного эмбриона, видимые на поперечном срезе, окрашенном по Гольджи. У большинства нейронов, по-видимому, пока только один длинный отросток - будущий аксон. Конусы роста вставочных нейронов остаются внутри спинного мозга, конусы роста мотонейронов выходят из него (чтобы направиться к мышцам), а конусы роста сенсорных нейронов прорастают в спинной мозг извне (оттуда, где расположены их тела). Многие клетки центральных областей спинного мозга у эмбриона еще делятся и пока не дифференцировались в нейроны и глиальные клетки. (S. Ramón у Cajal, Histologie du Systeme Nerveux de 1'Homme et des Vertebres, Paris: Maloine, 1909 1911; reprinted, Madrid: C.S.I.C., 1972.)
Большая часть имеющихся ныне сведений о свойствах конусов роста получена при изучении нервной ткани в культуре. Эмбриональные нервные клетки in vitro выпускают отростки, которые трудно идентифицировать как аксоны или дендриты и которые получили поэтому нейтральное название нейритов. Конус роста на конце каждого нейрита продвигается со скоростью около 1 мм в сутки. Это широкая утолщенная часть нейрита, похожая на ладонь со множеством длинных тонких микрошипиков, или филоподий, напоминающих пальцы (рис. 19-61). Филоподии находятся в непрестанном движении: в то время как одни втягиваются обратно в конус роста, другие, наоборот, удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему. «Перепонки» между филоподиями покрыты складчатой, как бы «гофрированной» мембраной (разд. 11.2.11). Как показывает электронная микроскопия, микротрубочки и микрофиламенты, имеющиеся в нейрите, в конусе роста оканчиваются, и широкая «ладонь» этого конуса заполнена уплощенными мембранными пузырьками, а также содержит митохондрии (рис. 19-62). В волнистых краях конуса роста и в филоподиях находится густая сеть актиновых филаментов. Все эти данные, полученные с помощью микроскопа, позволяют предполагать, что конус роста продвигается вперед способом, напоминающим передвижение ведущего края таких клеток, как нейтрофилы и фибробласты (разд. 11.6.4).

Рис. 19-61. Конусы роста: микрофотографии, полученные с помощью сканирующего электронного микроскопа. А. Конусы роста на конце нейрита куриного симптического нейрона в культуре. Бывший ранее единственным, конус роста недавно разделился на два. Обратите внимание на многочисленные филоподии и на то, что нейрит имеет вид натянутой струны в результате продвижения конусов роста, которые часто бывают единственными точками прочного сцепления с субстратом. Б. Конус роста сенсорного нейрона, «ползущий» по внутренней поверхности эпидермиса головастика Xenopus in vivo. (A - из D. Bray, Cell Behaviour [R. Bellairs, A. Curtis, G. Dunn, eds.] Cambridge, U.K.: Cambridge University Press, 1982; Б - из A. Roberts. Brain Res., 118, 526-530, 1976.)
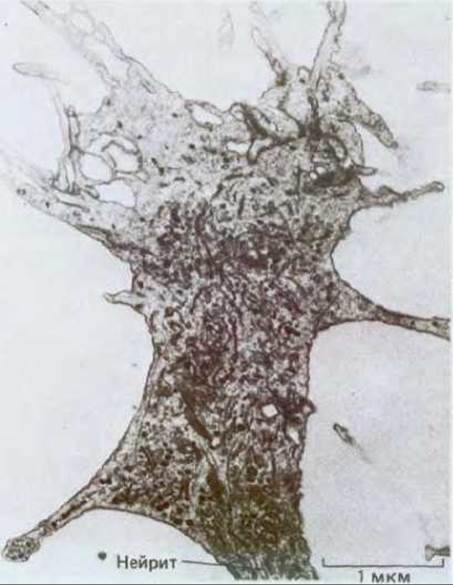
Рис. 19-62. Электронная микрофотография среза конуса роста. Видны многочисленные пузырьки неправильной формы, ограниченные мембраной. По-видимому, это связано с высокой интенсивностью экзоцитоза и эндоцитоза. (С любезного разрешения Gerald Shaw.)
19.7.5. В конусе роста скапливаются и используются материалы, необходимые для роста [53]
Конус роста служит не только «локомотивом» для удлинения нейрита, но и участком, где встраиваются новые компоненты растущей клетки (рис. 19-63). Поскольку в нейроне рибосомы сосредоточены главным образом в теле клетки, именно оно и должно быть местом синтеза белков, используемых для роста нейрита. В теле клетки синтезируются и новые мембраны, которые затем переносятся в форме мелких пузырьков к конусу роста с помощью быстрого аксонного транспорта (разд. 19.1.4). По мере поступления этих пузырьков в конус роста они включаются в плазматическую мембрану путем экзоцитоза. Хотя часть мембранного материала вновь поглощается при эндоцитозе и используется повторно, в общем итоге его количество при росте нейрита возрастает. Данные о таком способе роста были получены при наблюдении за передвижением мелких частиц пыли, прилипших к наружной поверхности растущего нейрона: частицы на поверхности самого конуса роста быстро продвигаются, тогда как частицы, находящиеся ближе к основанию нейрита, остаются неподвижными относительно тела клетки даже при удлинении отростка. «Рельсами» для быстрого аксонного транспорта служат микротрубочки (разд. 11.4.8); очевидно, мембранные пузырьки, передвигающиеся вдоль микротрубочек, доставляются к тем местам, где эти пути оканчиваются. Различные эксперименты позволяют предположить, что от микротрубочек зависит, где может образоваться конус роста, так как они способны регулировать доставку мембран.
В то же время и сами микротрубочки должны расти, так же как и остальной цитоскелет. С потоком медленного аксонного транспорта (разд. 19.1.4) из тела клетки переносится тубулин, но еще не ясно, где происходит сборка субъединиц в микротрубочки. Известно, однако, что обычно микротрубочки ориентированы таким образом, что их плюс-концы (быстро растущие концы, см. разд. 11.4.2) направлены к конусу роста. Известно также, что конусы роста необычайно чувствительны к локальному воздействию препаратов, препятствующих сборке микротрубочек. Это позволяет думать, что микротрубочки удлиняются путем добавления субъединиц в конусе роста.
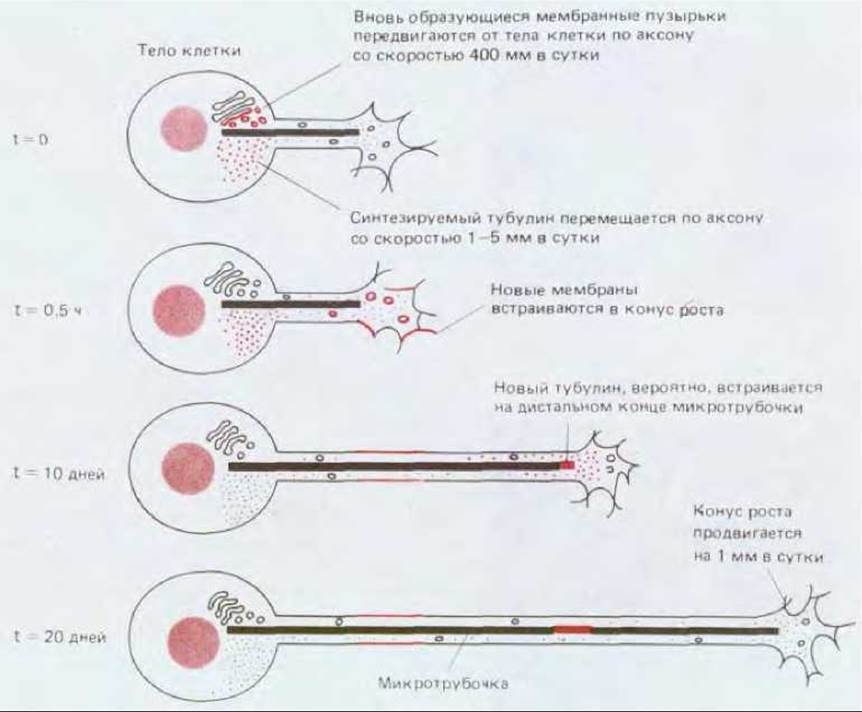
Рис. 19-63. На этих схемах показано, как транспортируется материал, необходимый для роста нейрита, из тела клетки к конусу роста, где происходит его включение. Для простоты показана только одна микротрубочка. Микротрубочки служат путями для быстрого аксонного транспорта мембранного материала. Тубулин переносится от тела клетки с помощью медленного аксонного транспорта. Место добавления субъединиц для удлинения микротрубочек еще точно не установлено.
19.7.6. Движение конуса роста in vitro может направляться избирательной адгезией, хемотаксисом и электрическими полями [52, 54]
«Упрощенные» условия клеточной культуры дают возможность исследовать механизмы, которые могли бы направлять движение конусов роста у интактного животного. Конусы роста, так же как нейтрофилы и фибробласты, при выборе субстрата предпочитают поверхности, к которым они прилипают наиболее прочно (рис. 19-64). По мере продвижения вперед конусы роста непрерывно вытягивают микрошипики к участкам, лежащим впереди и сбоку. Некоторые из этих выступов могут прикоснуться к менее адгезивному субстрату и в этом случае относительно быстро втягиваются обратно; другие встречают более адгезивную поверхность и сохраняются дольше. По-видимому, микрошипики, словно щупальца, исследуют близлежащие поверхности и направляют конус роста по пути с наиболее сильными адгезивными свойствами.
Однако на продвижение конусов роста влияет не только адгезивность субстрата. Важна также форма поверхности - например, конусы роста, «прицепившиеся» к волокнам, будут склонны двигаться вдоль них (феномен, называемый "contact guidance"). Определенное влияние, видимо, оказывают и вещества, растворенные во внеклеточной жидкости.
Если эмбриональный ганглий с сенсорными нейронами, которые будут иннервировать, например, челюсть, поместить в культуре на расстоянии около миллиметра от зачатка челюсти, нейриты будут расти преимущественно в сторону этого зачатка; это позволяет предположить, что ткань-мишень секретирует молекулы, вызывающие хемотаксический эффект. Электрическое поле тоже оказывает сильное направляющее воздействие, заставляя конусы роста в культуре нервной ткани продвигаться к катоду; электрическое поле в 7 мВ/мм уже эффективно.
Хотя подобные эксперименты in vitro показывают, какие факторы способны направлять движение конусов роста, они ничего не говорят о том, какой именно вид воздействия играет главную роль в развивающемся организме.
Ограничено ли продвижение конусов роста специфическими путями или эти пути выбираются путем проб и ошибок? Некоторые ответы удалось получить, исследуя поведение конусов роста в естественных условиях.
19.7.7. In vivo конус роста направляет движение нейрита по строго определенному пути ("pathway guidance") [55]
Обычно в живом организме конусы роста продвигаются к своим мишеням по строго определенным путям. Изучение действующих при этом механизмов у большей части позвоночных затруднено; несколько проще проводить исследования на некоторых беспозвоночных, например на прямокрылых насекомых, у которых можно детально проследить иннервацию развивающихся конечностей (рис. 19-65).
Сенсорные нейроны, иннервирующие у этих насекомых конечности, образуются у эмбриона из особых клеток в эпителии зачатков ног; тела нервных клеток остаются на периферии, а аксоны прорастают в центральную нервную систему по точно определенным зигзагообразным путям. Эти пути прокладываются в каждой конечности одним или двумя аксонами-пионерами, которые можно селективно окрасить с помощью антител или путем инъекции в тело клетки флуоресцентного красителя люциферина желтого. В результате можно увидеть, что на каждом повороте пути конусы роста «первопроходцев» вступают в контакт со специфическими клетками, которые служат чем-то вроде «дорожных указателей» (рис. 19-66). Конусы роста образуют на этих клетках временные щелевые контакты: если ввести в аксон краситель, то эти вспомогательные клетки тоже ярко окрасятся. Микрошипики, выпускаемые первыми конусами роста, достигают длины 50 или даже 100 мкм; этого достаточно, чтобы дотянуться до следующей клетки-указателя на пути аксона. Микрошипики, вступившие в контакт с такой клеткой, стабилизируются, а остальные втягиваются обратно. Таким образом конус роста продвигается шаг за шагом к центральной нервной системе. Если клетку-указатель разрушить лазерным лучом раньше, чем конус роста достигнет ее, то конус в этой точке «заблудится» (рис. 19-66). На тех участках пути, где в норме нет клеток-указателей, конус роста будет продвигаться в соответствии с адгезивностью базальной мембраны. лежащей под эпителием конечности (разд. 14.2.15). На всем пути конус роста продвигается благодаря специфическим молекулам на его поверхности, которые позволяют ему прилипать к соответствующему субстрату. Некоторые из этих адгезионных молекул уже идентифицированы.
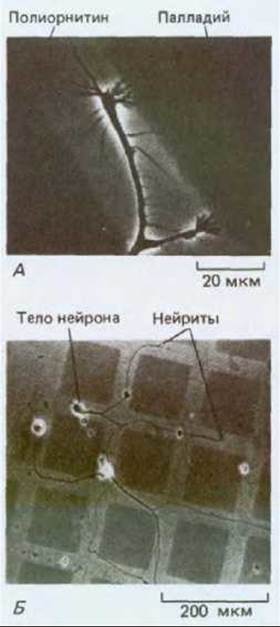
Рис. 19-64. Можно проследить за выбором субстрата конусом роста, культивируя клетки в чашке, поверхность которой сначала покрыта полиорнитином, а затем сверху нанесены квадраты из палладия. Так как поверхность клеток чаще всего заряжена отрицательно, клетки сильно прилипают к полиорнитину, который несет положительный заряд. Конусы роста продвигаются по «дорожкам» из полиорнитина и избегают палладия; но если клетке предоставлен выбор между палладием и еще менее адгезивным субстратом, они будут перемещаться по палладию. А. Фазовоконтрастная микрофотография при сильном увеличении, показывающая конусы роста на границе между двумя субстратами. Б. То же при меньшем увеличении; пути, выбираемые конусами роста, обозначены нейритами, которые тянутся за ними и остаются прикрепленными к полиорнитину. (P. Letoumeau, Dev. Biol., 44, 92-101, 1975.)
19.7.8. Конусы роста используют специфические адгезионные молекулы для сцепления с поверхностью клеток и внеклеточным матриксом [56]
Как только первые нейриты проложили путь, другие следуют за ними по принципу контакта: конусы роста прилипают к уже существующим нейритам и продвигаются вдоль них. Это универсальная тенденция, наблюдаемая как у позвоночных, так и у беспозвоночных. Поскольку существует сильная адгезия между нейритами и между нейритом и конусом роста, нервные волокна у взрослого животного оказываются сгруппированными в плотные параллельные пучки (фасцикулы). Таким же путем образуются крупные периферические нервы позвоночных, видимые невооруженным глазом (хотя впоследствии отдельные аксоны изолируются друг от друга оболочками из шванновских клеток). У позвоночных найдены специфические интегральные мембранные гликопротеины, обеспечивающие адгезию между нейритами. Примерами могут служить два хорошо изученных гликопротеина - так называемая молекула адгезивности нервных клеток, или N-CAM (разд. 14.3.6), и гликопротеин L1, известный также как молекула адгезивности нейрон-глия, или Ng-CAM. Было показано, что антитела к этим белкам, относящиеся к суперсемейству иммуноглобулинов (разд. 14.3.6), подавляют тенденцию развивающихся нейритов к образованию пучков и нарушают нормальный рост аксона (в разной степени в разных частях нервной системы). N-CAM имеется не только у нейронов, но и у глиальных клеток, а во время развития и регенерации и на многих других клетках, включая мышечные. В последнем случае эти молекулы, возможно, помогают «притягивать» конусы роста мотонейронов в те области, где должны сформироваться синапсы.
Движение конусов роста направляется их прилипанием не только к поверхности других клеток, но и к различным компонентам внеклеточного матрикса. Важным примером может служить регенерация нерва.

Рис. 19-65. Микрофотография зародыша прямокрылого (вид снизу), полученная с помощью сканирующего электронного микроскопа. Видны зачатки конечностей (показано стрелкой). (D. Bentley, H. Keshishian, Science, 218, 1082-1088, 1982. Copyright 1982 by the AAAS.)
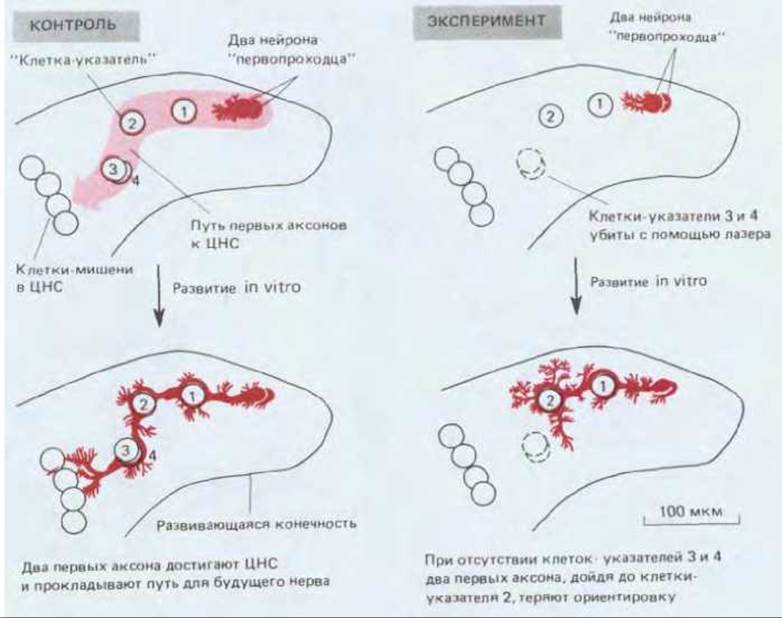
Рис. 19-66. В конечности у зародыша прямокрылого первые аксоны выбирают направление роста с помощью специальных клеток-«указателей». Если эти клетки разрушены, аксоны не способны выбрать верный путь. Позднее из клеток-указателей развиваются нейроны, аксоны которых направляются к ЦНС по путям, проложенным аксонами-первопроходцами. (По D. Bentley, M. Caudy, Nature, 304, 62-65, 1983.)
Если перерезать периферический нерв, то аксоны обычно регенерируют с помощью конусов роста, образующихся на разрезанных концах. Эти конусы движутся по тоннелям из базальной пластинки, которую ранее создали шванновские клетки, одевавшие аксоны дистальной, дегенерировавшей части нерва. Имеются данные в пользу того, что ключевую роль в этом процессе играют ламинин (разд. 14.2.15) или комплекс ламинина с протеогликаном, содержащим гепарансульфат: они связываются с рецепторами, которые находятся в мембране конуса роста и относятся к семейству интегринов (разд. 14.2.17). Ламинин и упомянутый комплекс ускоряют рост нейритов in vitro, а антитела к комплексу подавляют регенерацию нервов in vivo.
Хотя белки, подобные N-CAM, Ng-CAM и ламинину, играют, по-видимому, важную роль в клеточной адгезии и в управлении миграцией конусов роста, свойства этих белков не дают ясного ответа на главный вопрос: почему одни конусы роста выбирают один путь, тогда как другие предпочитают другой?
19.7.9. Организация нервных связей определяется различиями в свойствах нейронов: теория нейронной специфичности [57]
В развивающейся конечности куриного зародыша, как и в других подобных случаях, конусы роста аксонов продвигаются по строго определенным путям (рис. 19-67). Эти пути на своем протяжении ветвятся, и разные ветви идут к разным мишеням. Поэтому в каждой из точек ветвления перед конусом роста встает проблема выбора одного из возможных направлений. Этот ряд последовательных выборов осуществляется в соответствии с точными правилами, и в результате создается высокоупорядоченная система соединений между нейронами и их клетками-мишенями. Такие соединения можно проследить при помощи метода с использованием пероксидазы хрена (разд. 19.1.5). Оказалось, что тела мотонейронов, иннервирующих одну и ту же мышцу, образуют компактную группу, расположенную у всех особей в одном и том же участке спинного мозга, а тела клеток, иннервирующих разные мышцы, находятся в разных участках (рис. 19-68).

Рис. 19-67. Зачатки крыльев восьмидневного куриного зародыша (световая микрофотография, препарат окрашен серебром, чтобы выявить расположение нервов). Сравните правое крыло с левым: нервы располагаются на обеих сторонах тела почти в точности симметрично, что указывает на существование эффективной системы, направляющей рост нервов. Отдельные конусы роста должны делать выбор в каждой точке разветвления пути, и этот выбор делается в соответствии со строгими правилами.
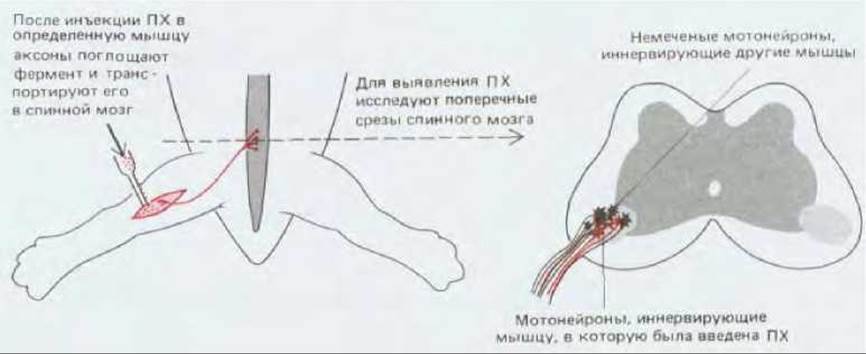
Рис. 19-68. Здесь показано, как можно использовать ретроградный перенос пероксидазы хрена (ПХ) для выявления спинномозговых мотонейронов, иннервирующих определенную мышцу. Для ясности размеры нейронов, изображенных на поперечном разрезе, преувеличены и показано только три нейрона. На самом деле к каждой мышце подходит нерв, содержащий отростки множества нервных клеток (обычно нескольких сотен), тела которых в спинном мозгу расположены тесной группой.
Как же происходит выбор пути? Не попадают ли конусы роста в разные места назначения просто в зависимости от исходной позиции, подобно автомашинам на скоростном шоссе, где запрещено менять полосу движения? Чтобы проверить эту гипотезу, у раннего зародыша вырезали кусочек нервной трубки и перевертывали его на 180° еще до того, как начинался рост аксонов. Тогда мотонейроны, первоначально предназначенные для иннервации мышцы А, оказывались на месте нейронов для мышцы В, и наоборот. Как выяснилось, если перемещение не слишком велико, то конусы роста перемещенных нейронов все же приходят к мышцам, соответствующим первоначальному положению нейрона в нервной трубке, хотя для этого они вынуждены двигаться по измененным путям (рис. 19-69). Это означает, что нейроны, предназначенные для разных мышц, не эквивалентны (разд. 16.4.6): подобно нейронам головного мозга у мышей reeler, они различаются не только своим расположением, но и какими-то внутренними химическими особенностями. Такую неэквивалентность нейронов обычно называют нейронной специфичностью. Как уже говорилось в гл. 16, клетки соединительной ткани в различных областях зачатка конечности тоже неэквивалентны, так что различия между ними могли бы определять выбор того или иного пути тем или иным конусом роста.
В центральной нервной системе как у позвоночных, так и у беспозвоночных тоже, судя по имеющимся данным, определенные группы нейронов или глиальных клеток несут специфические метки, которые узнаются другими нейронами и тем самым помогают установить избирательные нервные связи. Но мы еще почти ничего не знаем о молекулах, участвующих в таких процессах как в центральной, так и в периферической нервной системе.

Рис. 19-69. Схематически представленный здесь эксперимент на курином зародыше показывает, что мотонейроны даже после их перемещения посылают свои аксоны к тем мышцам, которые соответствуют первоначальному положению этих нейронов в спинном мозгу зародыша. Обратите внимание, что аксоны мотонейронов, расположенных на разных уровнях спинного мозга, сближаются, образуя сплетение у основания конечности, а затем расходятся для иннервации различных мышц. Конус роста, проходящий через сплетение, должен выбрать один из многих путей.
19.7.10. Ткани-мишени выделяют нейротропные факторы, регулирующие рост и выживание нервных клеток [58]
В начальном участке пути движение конуса роста направляется тканями, в которых он перемещается; приблизившись к месту назначения, конус роста попадает под влияние мишени - часто еще до прямого соприкосновения с нею - благодаря действию нейротропных факторов, выделяемых клетками-мишенями. Как мы уже видели на примере ганглия, иннервирующего зачаток челюсти, такие факторы могут служить хемотаксическими аттрактантами для конусов роста. Однако еще важнее то, что от них зависит выживание конусов роста, ветвей аксона и целых нейронов.
Первым из нейротропных факторов был идентифицирован фактор роста нервов (ФРН), и в настоящее время он лучше всего изучен. Этот фактор был открыт случайно в ходе экспериментов с трансплантацией тканей и опухолей куриным эмбрионам. Трансплантаты одного вида опухолей необычайно обильно иннервировались и вызывали значительное разрастание определенных групп периферических нейронов в близлежащих областях. Такому влиянию подвергались нейроны только двух категорий: сенсорные и симпатические (подкласс периферических вегетативных нейронов, регулирующих сокращение гладкой мускулатуры и функцию экзокринных желез). Экстракты из опухоли стимулировали также рост нейритов в культуре этих нейронов. Дальнейшие исследования показали, что в культуре другой ткани-слюнной железы самца мыши - такой же стимулирующий фактор образуется в огромных количествах. Эта «игра природы» пока еще не разгадана, так как образование ФРН клетками слюнной железы не имеет видимой связи с главной функцией этого фактора, но так или иначе открылась возможность получать чистый ФРН в количествах, достаточных для выяснения его химической природы и изучения его функций. Оказалось, что активность связана с белком- димером, содержащим две идентичные полипептидные цепи из 118 аминокислотных остатков каждая. После того как ФРН был выделен в чистом виде, появилась возможность получать антитела, блокирующие его действие. Если антитела к ФРН ввести мыши, у которой развитие нервной системы еще не закончено, то большая часть симпатических нейронов и некоторые сенсорные нейроны погибнут.
Точно так же и в культуре ткани без ФРН симпатические нейроны и часть сенсорных нейронов погибают; если же ФРН присутствует, они выживают и образуют нейриты (рис. 19-70). Влияние на выживание нейронов и на образование нейритов - два разных эффекта ФРН. Это можно наглядно показать, если поместить клетки в средний отсек культуральной чашки, отделенный от двух боковых отсеков перегородкой, которая препятствует смешиванию сред, но допускает прорастание нейритов (рис. 19-71). Когда ФРН имеется во всех трех отсеках, нейриты прорастают во все три. Если в одном из боковых отсеков ФРН отсутствует, нейриты в него не прорастают. И наконец, если ФРН удалить из бокового отсека, в котором уже появились нейриты, то они уменьшаются и втягиваются до перегородки между отсеками. Клетки в центральном отсеке не смогут выжить и выпустить отростки, если ФРН не будет присутствовать здесь с самого начала; но если вначале во все отсеки добавить ФРН, а затем, после того как нейриты прорастут в боковые отсеки, убрать ФРН из центрального, то клетки выживают и рост нейритов в боковых отсеках продолжается.
Таким образом, ФРН не только оказывает локальное воздействие на отдаленные части клетки, поддерживая и стимулируя рост нейритов и конусов роста, но служит также фактором выживания для всей клетки в целом. При локальном воздействии на конус роста наблюдается прямой, быстрый эффект, не зависящий от связи с телом клетки; если среду, содержащую ФРН, заменить средой без ФРН, то конусы роста уже через 1-2 мин прекратят движение. Помимо непосредственной реакции на ФРН конусы роста клеток, чувствительных к ФРН, поглощают его путем эндоцитоза, а затем ФРН в пузырьках переносится с помощью ретроградного аксонного транспорта к телу нейрона, где этот фактор (или какой-то внутриклеточный посредник), по-видимому, предотвращает гибель клетки.
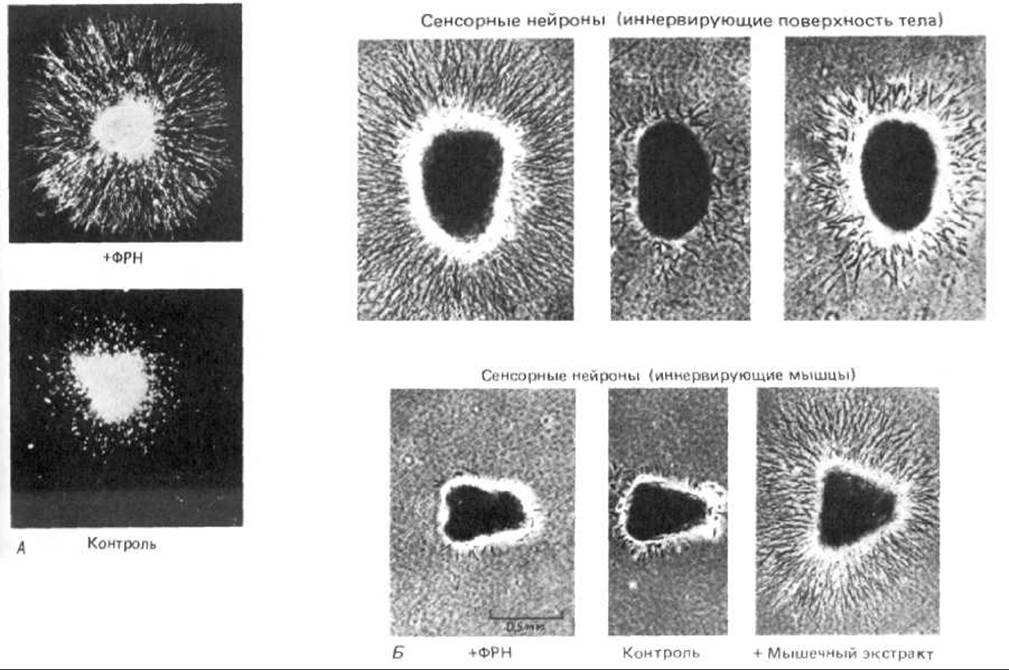
Рис. 19-70. А. Симпатический ганглий, культивируемый в течение 48 ч с ФРН (вверху) и без него (внизу) (микрофотография в темном поле). Нейриты прорастают из симпатических нейронов только в присутствии ФРН. Каждая культура содержит также шванновские клетки, мигрировавшие из ганглия, но ФРН на эти клетки не влияет. Б. Поведение сенсорных нейронов, которые росли 24 ч в среде с ФРН (слева), в контрольной среде без ФРН (в середине) и в среде, содержащей экстракт скелетной мышцы (справа) (фазовоконтрастные микрофотографии). Сенсорные нейроны, представленные в верхнем ряду, иннервируют главным образом поверхность тела (но, кроме того, посылают небольшое число отростков к скелетным мышцам и другим мишеням). Большая часть этих клеток реагирует на ФРН так же, как и симпатические нейроны, показанные на фото А. Сенсорные нейроны на фотографиях нижнего ряда в норме иннервируют скелетные мышцы (обеспечивая сенсорную обратную связь). Эти нейроны нечувствительны к действию ФРН, но сильно реагируют на добавление экстракта, приготовленного из скелетной мышцы. (А -с любезного разрешения Naomi Kleitman; Б из A.M. Davies. Dev. Biol., 115; 56-67, 1986.)
19.7.11. В результате гибели клеток число выживших нейронов регулируется в соответствии с количеством ткани- мишени [59]
У позвоночных спинномозговые сенсорные ганглии образуются по сегментарной схеме, соответствующей ряду позвонков в позвоночнике. Каждый ганглий состоит из группы сенсорных нейронов, происшедших из нервного гребня и посылающих один нейрит наружу, к периферии тела, а другой - внутрь, к спинному мозгу. Вначале зачатки всех ганглиев одинаковы по размерам, но у взрослого животного ганглии тех сегментов тела, где прикреплены конечности, содержат гораздо больше нейронов, чем ганглии других сегментов (рис. 19-72). Такое различие обусловлено главным образом гибелью клеток: значительная часть нейронов во многих ганглиях погибает. Если на ранней стадии развития удалить зачаток конечности, то ближайшие к нему ганглии уменьшатся до размеров остальных; и напротив, если в грудную область зародыша пересадить еще один, добавочный зачаток конечности, то он будет иннервироваться и на этом уровне в ганглии сохранится необычно большое число нейронов. Полагают, что выживание нейронов ганглия в соответствии с количеством ткани-мишени в значительной степени опосредуется ФРН, выделяемым этой тканью. Если на определенной стадии развития в эмбрион ввести дополнительное количество ФРН, то многие из тех нейронов, которые в норме погибли бы, сохранятся, так же как и нейроны в области ампутированного зачатка конечности.
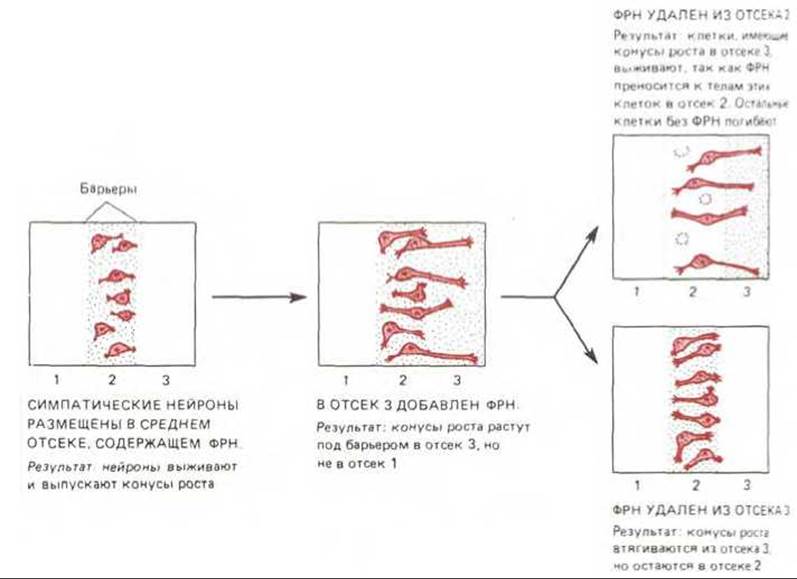
Рис. 19-71. Схема эксперимента на культуре ткани, доказывающего, что симпатические нейроны не только нуждаются для своего выживания в ФРН, но посылают конусы роста лишь туда и поддерживают нейриты лишь в тех областях, где имеется ФРН. Обратите внимание, что до тех пор, пока клетка имеет нейриты, доходящие до участков с ФРН, для выживания клетки не требуется присутствия ФРН в области ее тела.
Может показаться, что образовывать избыточные нейроны, а затем частично уничтожать их с целью регулирования их числа слишком расточительно. И все же эта стратегия широко используется у позвоночных в процессах развития как сенсорных, так и моторных нейронов, как в центральной нервной системе, так и в периферической. Например, около 50% всех мотонейронов, иннервирующих скелетные мышцы, погибают в процессе развития зародыша в течение нескольких дней после установления контакта с мышцами-мишенями. По-видимому, выживание нейронов в этих системах регулируют разнообразные трофические факторы, вроде ФРН, выделяемые клетками-мишенями. Такая стратегия обладает рядом важных преимуществ. Во-первых, этот механизм позволяет автоматически корректировать отклонения в относительных размерах различных частей тела. Во-вторых, эти процессы облегчают эволюцию: если в результате мутаций изменится величина какой-то части тела, то число иннервирующих ее нейронов будет автоматически приспосабливаться к этому и не понадобится дополнительных мутаций, изменяющих программу образования нейронов. И наконец, небольшое число нейротропных факторов, таких как ФРН, могут регулировать количественное соответствие большого числа мишеней и источников их иннервации, даже если система связей очень сложна. Благодаря аксонному транспорту фактор, выделяемый данной мишенью, селективно переносится в тела нейронов, иннервирующих эту мишень, а не каких-либо других нейронов, которые могут быть расположены там же и имеют сходные рецепторы, но посылают аксоны в другие места. Таким образом, гибель клеток, регулируемая нейротропными факторами роста, может помогать установлению точного соответствия между числом клеток в различных частях нервной системы.
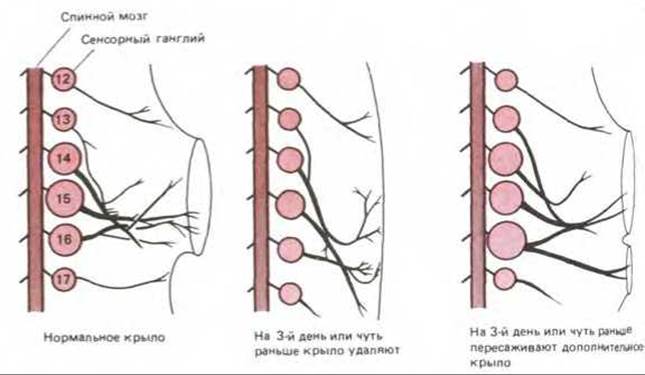
Рис. 19-72. Регуляция выживания нервных клеток в сенсорных спинномозговых ганглиях куриного эмбриона. Изображены ганглии и нервы 8-9дневного зародыша. Размеры каждого ганглия соответствуют числу выживших нейронов, которое в свою очередь зависит от количества ткани, иннервируемой данным ганглием. (По V. Hamburger, J. Exp. Zool. 68: 449 -494, 1934 а. J. Exp. Zool., 80; 347-389, 1939.)
19.7.12. Нервные связи создаются и разрушаются на протяжении всей жизни [60]
Есть данные, что даже в нормальной, неповрежденной нервной ткани дендриты и окончания аксонов непрерывно втягиваются и отрастают вновь. Например, можно наблюдать, как определенные нейроны зрелого вегетативного ганглия мыши втягивают одни дендритные ветви и выпускают другие в течение месяца (рис. 19-73). В нормальных условиях такие изменения происходят медленно и в ограниченной степени, но если часть клеток-мишеней лишается иннервации, этот механизм включается на полную мощность. Это можно показать на примере скелетной мышцы, перерезав часть иннервирующих ее аксонов; тогда денервированные мышечные волокна, очевидно, выделяют способный к диффузии фактор, стимулирующий образование новых конусов роста из нервных окончаний, сохранившихся на соседних мышечных клетках (рис. 19-74). Этот фактор скелетных мышц пока не идентифицирован, но для гладких мышц показано, что аналогичную роль там играет ФРН. Денервация приводит к увеличению количества ФРН в гладкой мышце (по крайней мере отчасти из-за того, что теперь его поглощает и отводит меньшее число нервных окончаний); избыток ФРН стимулирует рост аксонов по направлению к мышце, и иннервация восстанавливается.
Очевидно, в интактном организме ФРН действует так же, как и в культуре in vitro, т. е. как фактор выживания, определяющий, будут ли клетки жить или погибнут, и как локальный стимулятор активности конусов роста, регулирующий ветвление концевых участков аксона. Первая функция имеет особое значение в период развития, а вторая важна на протяжении всей жизни; однако обе они приводят к одному результату: с их помощью иннервация приспосабливается к потребностям мишени. Сейчас появляется все больше данных о существовании других нейротропных факторов роста, выполняющих такие же функции по отношению к другим видам нервных клеток (см. рис. 19-70). В следующем разделе мы увидим, что такие факторы, вероятно, играют важную роль в регулирующем влиянии электрической активности на развитие систем нервных связей.
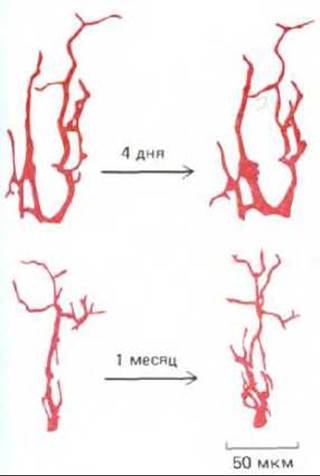
Рис. 19-73. Перестройка дендритов у нейронов верхнего шейного (вегетативного) ганглия мыши. У наркотизированного животного осторожно обнажали ганглий и для выявления дендритов вводили путем микроинъекции флуоресцентный краситель в тело одной из нервных клеток. Рану зашивали, а через несколько дней или недель операцию повторяли и снова вводили краситель в тот же самый нейрон. На верхних и нижних рисунках показаны два нейрона, повторно окрашенные через разные промежутки времени. Чем длиннее интервал между первой и второй инъекциями, тем больше изменений в картине ветвления дендритов. (Перепечатано с разрешения D. Purves, R. D. Hadley, Nature, 315, 404-406, 1985. Copyright 1985 Macmillan Fournals Limited.)
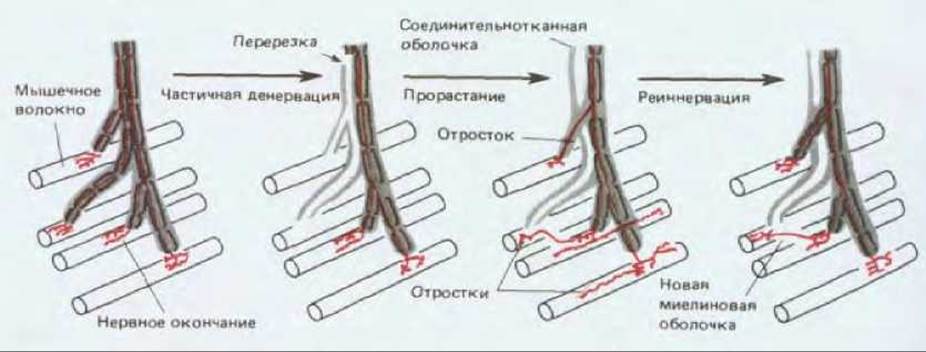
Рис. 19-74. Если перерезать часть аксонов, иннервирующих скелетную мышцу, то какая-то доля волокон этой мышцы лишится иннервации, тогда как у соседних волокон иннервация сохранится. Перерезанные аксоны дегенерируют, а оставшиеся аксоны, хотя они не были повреждены, начинают активно ветвиться в тех местах, где они лежат вблизи денервированного волокна. Через один или два месяца те из новых веточек, которые нашли путь к освободившимся участкам на денервированном мышечном волокне, образуют на нем стабильные синапсы и восстанавливают иннервацию, а все остальные веточки отмирают. Этот феномен позволяет предполагать, что денервированные мышечные волокна выделяют какой-то «фактор, способствующий прорастанию». (Воспроизведено с разрешения М.С. Brown, R. L. Holland, W. G. Hopkins, Ann. Res. Neurosci., 4, 17-42, 1981. Copyright 1981 by Annual Reviews Inc.)
Заключение
Развитие нервной системы удобно разделить на три этапа, которые частично перекрываются. На первом этапе нейроны образуются в соответствии с собственной программой клеточной пролиферации и вновь образующиеся клетки мигрируют из мест своего «рождения», чтобы упорядоченным образом расположиться в других участках. На втором этапе от клеток отрастают аксоны и дендриты, кончики которых продвигаются с помощью конусов роста. Конусы роста перемещаются по строго определенным путям, направляемые главным образом контактными взаимодействиями с поверхностью других клеток или с компонентами внеклеточного матрикса. Нейроны, предназначенные для связи с разными мишенями, ведут себя так, как если бы они обладали разными, только им присущими особенностями (нейронная специфичность), что может выражаться в различных свойствах клеточной поверхности, позволяющих конусам роста выбирать разные пути. В конце своего пути конус роста встречается с клеткой, с которой он должен образовать синапс, и оказывается под влиянием нейротропных факторов, выделяемых этой клеткой. Эти факторы регулируют ветвление аксона и передвижение конусов роста вблизи ткани-мишени и, кроме того, контролируют выживание нейронов, которым принадлежат конусы роста. С помощью этих двух эффектов нейротропные факторы, такие как фактор роста нервов (ФРН), регулируют плотность иннервации тканей-мишеней. На третьем этапе развития нервной системы, который будет рассмотрен в следующем разделе, образуются синапсы, а затем схема связей уточняется с помощью механизмов, зависящих от электрической активности.