Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Особенности растительных клеток
Перенос веществ между клетками
В предыдущем разделе мы говорили о том, что жесткая клеточная стенка порождает весьма специфические проблемы в связи с ростом и развитием растительных клеток. Кроме того, она сильно ограничивает возможности взаимодействия клеток, замурованных в ткани друг с другом и с окружающей их средой. Однако растительные клетки изобрели самые хитроумные способы преодоления этих ограничений. Непосредственная взаимосвязь клеток столь же важна для многоклеточных растений, как и для многоклеточных животных; поэтому возникли специальные канальцы, соединяющие цитоплазму растительной клетки с цитоплазмой соседних клеток (при этом обеспечивается контролируемый переход ионов и небольших молекул). Кроме того, у высших растений длинные тяжи цилиндрических клеток соединены друг с другом перфорациями, в результате чего образуются длинные трубки, обеспечивающие ток воды и питательных веществ.

Рис. 20-19. Специализированные слои небольших клеток у основания черешка, при участии которых осуществляется листопад. Два или три слоя небольших клеток секретируют ферменты, обусловливающие деградацию клеточной стенки. Черешок отваливается от стебля в том месте, где клеточные стенки тоньше. Несколько лигнифицированных клеток пересекают весь отделительный слой листа. Остающиеся в зоне отделения клетки откладывают суберин, в результате чего над раной образуется защитный слой.

Рис. 20-20. А. Цитоплазматические каналы, называемые плазмодесмами, пронизывают клеточные стенки и соединяют в единое целое все клетки растения. Б. Плазмодесма выстлана плазматической мембраной, без перерыва переходящей в мембраны обеих соседних клеток. Обычно в просвете плазмодесмы находится тонкая цилиндрическая структура, так называемая десмотубула, которая является производным эндоплазматического ретикулума
20.2.1. Растительные клетки соединены между собой специальными цитоплазматическими мостиками, так называемыми плазмодесмами [9]
За исключением очень немногих специализированных видов клеток, все живые клетки у высшего растения соединены с соседними при помощи тонких цитоплазматических каналов, называемых плазмодесмами, которые пронизывают разделяющие их клеточные стенки. Как показано на рис. 20-20, в области каждой плазмодесмы плазматическая мембрана одной клетки без перерыва переходит в мембрану соседней клетки. Сама плазмодесма представляет собой цилиндрический выстланный мембраной канал диаметром от 20 до 40 нм. По оси канала из одной клетки в другую тянется цилиндрическая структура меньшего диаметра - десмотубула, просвет которой, по данным электронной микроскопии, сообщается с полостями эндоплазматического ретикулума обеих смежных клеток (рис. 20-21). Пространство между наружной поверхностью десмотубулы и мембранной выстилкой плазмодесмы заполнено цитозолем (рис. 20-20). Очень часто у обоих концов канала отмечается сужение этого кольцевого слоя цитоплазмы. Возможно, что эти сужения играют очень важную роль, так как именно здесь каждая из клеток в принципе имеет возможность регулировать переход молекул из одной клетки в другую.
Обычно плазмодесмы образуются вокруг элементов эндоплазматического ретикулума, который в ходе цитокинеза оказывается погруженным в новую клеточную стенку, разделяющую материнскую клетку (см. рис. 13-71). Но плазмодесмы обнаруживаются также и в стенках клеток, которые не являются сестринскими. Кроме того, количество плазмодесм может увеличиваться при росте клеток, свидетельствуя о том, что они могут образовываться de novo.
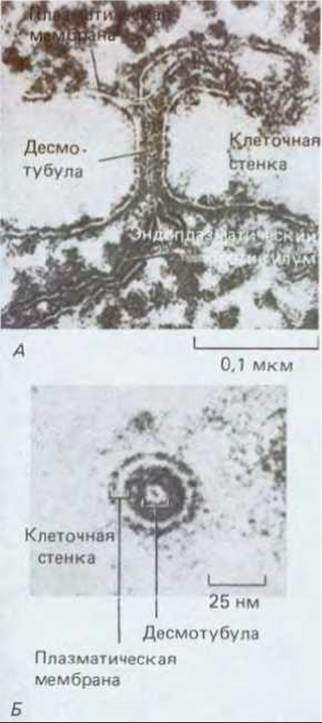
Рис. 20-21. Плазмодесмы на электронных микрофотографиях. А. Продольный срез плазмодесмы водного папоротника. Плазматическая мембрана выстилает пору и, не прерываясь, переходит из одной клетки а другую. Видны эндоплазматический ретикулум и связанная с ним центральная десмотубула. Б. Такая же плазмодесма на поперечном срезе. (С любезного разрешения R. Overall.)

Рис. 20-22. Использование различных по размеру, флуоресцирующих пептидов для определения функционального размера каналов в плазмодесмах. А. Флуоресцирующий пептид с мол. массой более 850 дальтон. Б. Флуоресцирующий пептид с мол. массой менее 850 дальтон. В обоих случаях пептид вводили в одну клетку эпидермиса рдеста маленького. Обратите внимание, что пептид большого размера почти не продвинулся, тогда как меньший пептид проник во многие соседние клетки. (С любезного разрешения Р. В. Goodwin.)
20.2.2. Плазмодесмы позволяют молекулам непосредственно переходить из одной клетки в другую [9, 10]
Что же еще, помимо характерной структуры, может служить свидетельством в пользу предполагаемой роли плазмодесм в межклеточной связи? На эту роль косвенно указывает тот факт, то плазмодесмы особенно многочисленны в стенках клеток, группирующихся в зонах интенсивной секреции, например в нектарниках цветков. У таких клеток на 1 мкм3 поверхности клеточной стенки может приходиться 15 и более плазмодесм, тогда как у других клеток это число нередко бывает меньше единицы.
Наиболее прямые указания на межклеточный транспорт через плазмодесмы были получены в экспериментах с введением красителей и с пропусканием электрического тока. Так, например, флуоресцентные красители лишь с трудом проходят через плазматическую мембрану, однако после введения их с помощью микрокапилляра в одну из клеток листа элодеи они довольно быстро появляются в соседних клетках. Точно так же при подаче электрических импульсов внутрь одной из клеток эти импульсы регистрируются (хотя и в ослабленном виде) электродами в соседних клетках. Степень ослабления электрического сигнала зависит от плотности расположения плазмодесм и от числа клеток, находящихся между электродами. Кроме того, электрод, приложенный к наружной поверхности плазматической мембраны, не улавливает сигналов, поданных внутрь клетки; значит, они распространяются какими-то внутриклеточными путями.
Таким образом, представляется вероятным, что именно плазмодесмы обеспечивают транспорт растворенных веществ между соседними растительными клетками, подобно тому, как щелевые контакты обеспечивают межклеточный транспорт у животных (см. разд. 14.1.5). Данные, полученные при микроинъекции флуоресцентных красителей, соединенных с пептидами различных размеров, свидетельствуют о том, что по плазмодесмам могут перемещаться молекулы с массой, не превышающей 800 дальтон (рис. 20-22), что примерно соответствует ограничениям по размеру для щелевых контактов. Ряд данных свидетельствует о том, что транспорт через плазмодесмы регулируется сложным образом. Так, эксперименты по введению красителей показали, что, несмотря на наличие внешне нормальных плазмодесм, между некоторыми системами тканей перенос даже низкомолекулярных веществ затруднен. Например, передвижение краски не обнаруживается между клетками корневого чехлика и клетками кончика корня, или между клетками эпидермиса и коры как в корнях, так и в побегах. Механизмы, ограничивающие в этих случаях сообщение между клетками, остаются пока неизвестными, хотя имеются некоторые указания на то, что в них могут быть задействованы ионы Са2+ и фосфорилирование белка. С другой стороны, показано, что определенные вирусы растений способны вызывать расширение плазмодесм, обеспечивая себе возможность передвижения из клетки в клетку (рис. 20-23). Например, установлено, что у вируса табачной мозаики транспорт через плазмодесмы зависит от одного-единственного вирусного белка (Р30) с молекулярной массой 30000: дефектный вирус, неспособный перемещаться из клетки в клетку в обычном растении, вполне успешно делает это в трансгенном растении табака, содержащим соответствующий ген.

Рис. 20-23. На этой электронной микрофотографии видно, как мелкие сферические вирусные частицы переходят по плазме из одной клетки растения в другую. Диаметр этих растительных вирусов составляет 25 нм, что значительно превышает размер пептидов, не способных проходить через эти отверстия. (С любезного разрешения К. Plaskitt.)

Схема 20-2. Особенности раннего развития цветковых растений.

Рис. 20-24. Сильно схематизированное изображение группы растительных клеток, соединенных плазмодесмами. Плазматическая мембрана, которая выстилает плазмодесмы, разделяет весь объем растения на два компартмента: внеклеточный и внутриклеточный (симпласт). Для большей ясности клеточные органеллы на схеме не показаны.
20.2.3. У высших растений биологические жидкости разделены на два больших компартмента - внутриклеточный и внеклеточный
Плазмодесмы соединяют плазматическую мембрану и цитоплазму соседних клеток, превращая их в сложное сообщество живых протопластов. Поэтому все тело растения можно рассматривать как систему из двух компартментов: 1) внутриклеточного - так называемого симпласта, состоящего из объединенного множества протопластов (включая и протопласты ситовидных трубок флоэмы) и ограниченного объединенной плазматической мембраной всех живых клеток и 2) внеклеточного, или апопласта, включающего все клеточные стенки и мертвые пустые проводящие ксилемы, а также находящуюся в тех и в других воду (рис. 20-24). Оба компартмента имеют свои собственные транспортные системы, однако в определенных точках они могут сообщаться между собой, а также подвергаться локальной модификации для обеспечения контроля протекающих между ними обменных процессов.
20.2.4. Фотосинтезирующие и поглощающие клетки связаны друг с другом сосудистыми тканями, в состав которых входит ксилема и флоэма [11]
Многоклеточная организация у растений, как и у животных, делает возможным разделение функций, при котором различные типы клеток дополняют друг друга благодаря специализации, приобретаемой ими в процессе дифференцировки. Две важнейшие специфические для растений функции осуществляются фотосинтезирующими клетками, которые содержат хлоропласта и служат для всего организма источником органических веществ, в частности сахарозы, и всасывающими клетками, которые поглощают из окружающей среды воду и растворенные минеральные вещества. У большинства высших растений эти две функции не могут выполняться одними и теми же клетками, поскольку для первой из них необходим свет, а вторая осуществляется в толще почвы и темноте. Для каждого из этих процессов требуется и ряд других условий. Фотосинтез, например, должен протекать в особой микросреде. где строго регулируется относительная влажность и содержание двуокиси углерода. Достигается это с помощью устьиц - особых отверстий в покрытом кутикулой эпидермисе листа, которые способны открываться и закрываться в зависимости от тургора замыкающих клеток (см. рис. 20-11). С другой стороны, для эффективного поглощения веществ из почвы нужна очень большая всасывающая поверхность, которую обеспечивают корни; необходимы также мембранные транспортные системы, к которым часто добавляются транспортные системы симбиотических микроорганизмов. Таким образом, фотосинтезирующие и всасывающие клетки питают друг друга, а вместе - снабжают все остальные части растения минеральными и органическими веществами, необходимыми для процессов биосинтеза. Чтобы обеспечить дальний транспорт этих веществ, фотосинтезирующие и всасывающие ткани связаны с ксилемой и флоэмой, которые образуют разветвленную сеть проводящих элементов. Каждый из них состоит из цепей цилиндрических клеток, соединенных своими концами и образующих трубочки (см. схему 20-1 и рис. 20-16).
20.2.5. Вода и растворенные в ней соли передвигаются по ксилеме [12]
Ксилема - сложный компонент системы проводящих тканей. Зрелые элементы сосудов ксилемы представляют собой погибшие клетки, лишенные цитоплазмы. Боковые стенки их сильно лигнифицированы, и на их внутренней стороне имеются вторичные утолщения. Эти трубочки обеспечивают перенос воды и растворенных в ней неорганических ионов от корней к остальным частям растения (рис. 20-25). Ксилема также осуществляет опорную функцию, особенно у древесных растений. Ток жидкости в ксилеме направлен в одну сторону - к местам испарения влаги. Вода насасывается в трубочки, похожие на капилляры, благодаря испарению. Лигнин откладывается вокруг сосудов ксилемы таким образом, что получаются очень устойчивые к сжатию структуры, что весьма важно для трубочек, несущих жидкость, находящуюся под отрицательным давлением. Без подобного укрепления длинные трубочки попросту бы слиплись, как тонкая соломка для коктейлей.
По сосудам ксилемы транспортируются главным образом минеральные соли и азотсодержащие соединения, которые, вероятно, поступают сюда в результате активной секреции их клетками корневой паренхимы.

Рис. 20-25. Две главные проводящие системы - ксилема и флоэма, с их помощью вода и растворенные в ней вещества транспортируются по всему растению. Это сильно упрощенная схема, в частности на ней не отражен интенсивный водный обмен, происходящий между ксилемой и флоэмой через боковые стенки.
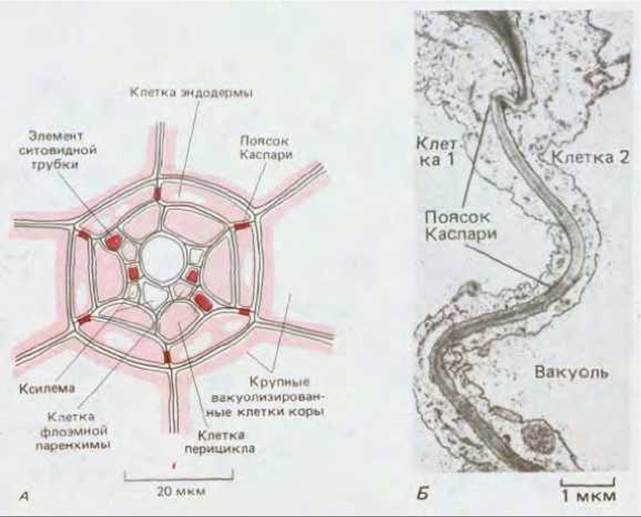
Рис. 20-26. А. Проводящая сеть низкоорганизованного растения, водного папоротника Azolla (схематический поперечный разрез осевой части корня). У этого растения картина расположения проводящих структур чрезвычайно проста: имеются четыре сосуда ксилемы и четыре элемента ситовидных трубок флоэмы. У большинства высших растений распределение клеток проводящих тканей гораздо сложнее. Обратите внимание на то, что ксилема и флоэма окружены клетками эндодермы и что пояски Каспари, занимающие в эндодерме стратегические позиции, препятствуют утечке воды из сосудистой системы через апопласт. Б. Поясок Каспари между двумя эндодермальными клетками корня, аналогичными представленным на рис. А. В отличие от обычной первичной клеточной стенки, поясок Каспари имеет гладкую текстуру из-за высокого содержания кутина; гидрофобные свойства кутина делают эти участки клеточной стенки непроницаемыми для воды, (Б - по В. Gunning, M. Stееу, Ultrastucture and Biology of Plant Cells. London: Arnold 1975.)
Утечка растворенных веществ в обратном направлении через апопласт на входных участках ксилемы блокируется так называемыми поясками Каспари, которые по своей функции аналогичны плотным контактам между соседними эпителиальными клетками животных (рис. 20-26). На выходе проводящих путей ксилемы особые паренхимные клетки, снабженные специфическими транспортными белками, локализованными в мембране, перекачивают растворенные вещества в фотосинтезирующие ткани. Большая часть растворителя (воды), проходящего по сосудам ксилемы, в конце концов испаряется, главным образом с поверхности фотосинтезирующих тканей листа.
Так как накачивание растворенных веществ в ксилему корней происходит непрерывно, оно может вызвать осмотический поток, обеспечивающий доставку растворенных веществ к листьям и побегам даже в тех случаях, когда погодные условия затрудняют испарение. Капельки воды, которые можно увидеть на кончиках листьев луговых трав ранним утром, выделяются именно благодаря этому феномену корневого давления, обусловленного осмосом. (Описание некоторых типов клеток, участвующих в этом процессе, см. на схеме 20-1.)
20.2.6. Сахара переносятся под действием давления, возникающего во флоэме [13]
Флоэма представляет собой сложный комплекс клеток проводящей ткани, обеспечивающий транспорт растворенных органических веществ, главным образом сахарозы, из фотосинтезирующих клеток листа к остальным частям растения (см. рис. 20-25). Основной проводящий компонент флоэмы - это ситовидная трубка ~ длинная колонка из живых цилиндрических клеток, которые сообщаются друг с другом через отверстия в торцевых участках их клеточных стенок (ситовидные пластинки). Сахароза поступает в клетки, образующие верхнюю часть ситовидной трубки и в виде концентрированного раствора (обычно 10-25%) перемещается по трубке вниз, переходя из одной ее клетки в другую. Клетки с толстой стенкой образуют трубку, пригодную для транспорта жидкости под высоким давлением (до 30 атм). Хотя зрелые ситовидные элементы представляют собой живые клетки с функционирующей плазматической мембраной (и, следовательно, являются частью симпласта), ядра и часть цитоплазмы они потеряли. Их существование обеспечивают связанные с ними клетки-спутники, которые переносят растворенные питательные вещества и другие молекулы в обоих направлениях: внутрь ситовидных трубок и из них через собранные в группы плазмодесмы, расположенные на общих боковых стенках (см. схему 20-1).

Рис. 20-27. Специализированные передаточные клетки в небольшой жилке листа. Элемент ситовидной трубки флоэмы (СТ) окружен тремя передаточными клетками (ПК). Впячивания клеточной стенки, выстланные плазматической мембраной, увеличивают поверхность этих клеток в двадцать раз. Передаточные клетки обнаруживаются в тех областях растения, в которых скорость переноса растворенных веществ через плазматическую мембрану особенно высока, например, там, где неорганические ионы перекачиваются из ксилемы в ткани или, как в данном случае, где сахароза накачивается во флоэму.
Транспорт по флоэме - это гораздо более сложный процесс, чем передвижение веществ по ксилеме, поскольку он не ограничивается одним направлением: растворенные органические вещества, главным образом сахароза, переносятся от мест их синтеза к местам потребления и хранения, независимо от того, где эти места расположены. Сахароза активно переносится внутрь ситовидных трубок и из них специализированными передаточными клетками, расположенными в источниках и потребителях соответственно) (рис. 20-27). Повышение концентрации сахара в источниках приводит к тому, что во флоэму в этих местах поступает больше жидкости, при этом создается давление, необходимое для сильного тока жидкости через ситовидные трубки к потребителям метаболитов. Здесь сахар в основном задерживается, а вода удаляется осмотическим путем (главным образом в ксилему). Жидкость движется по флоэме со скоростью около 1 м/ч, это значительно превышает скорость диффузии.
Следует заметить, что транспорт жидкости у растений имеет по меньшей мере две характерные особенности, отличающие его от аналогичного процесса у животных. Во-первых, животные имеют только одну транспортную систему - кровеносную, растения же обладают двумя различными системами - флоэмой и ксилемой. Во-вторых, у растений жидкости не циркулируют по замкнутому пути, подобно крови у животных; вместо этого здесь поддерживается постоянный ток воды от корней к листьям по двум «открытым трубопроводам» (см. рис. 20-25).
Заключение
Наличие жесткой относительно непроницаемой клеточной стенки в значительной степени определяет специфику взаимодействия растительных клеток друг с другом, а также с окружающей средой. Все живые клетки растения соединены между собой плазмодесмами - миниатюрными цитоплазматическими каналами, выстланными плазматической мембраной, которые пронизывают клеточные стенки и обеспечивают переход многих растворенных веществ из клетки в клетку. Таким образом, все живые протопласты растительного организма составляют единую систему - так называемый симпласт. Остальное пространство, занятое клеточными стенками и отмершими «пустыми» клетками, по которым в растении транспортируется большая часть воды, называется апопластом. Фотосинтезирующие клетки растений производят сахара, которые переходят во все остальные органы и ткани растения через живые клетки флоэмы, составляющие часть симпласта. Клетки корней поглощают из почвы воду и растворенные в ней минеральные вещества, транспортируемые затем к листьям по сосудам ксилемы, образованным отмершими клетками, которые входят в состав апопласта. И снова физические силы, в данном случае испарение, обеспечивают непрерывное продвижение столба воды от корня к листьям.