Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Молекулярная биология. Предмет и основные разделы курса
Функционирование клеток
Вообще говоря, клетка - это просто некий объем водной среды, отделенный от окружающей среды клеточной мембраной, которая называется плазматическая мембрана, и которая не дает свободно перемещаться молекулам внутрь клетки и наружу из клетки. Кроме того, клетки эукариот имеют развитую систему внутренних мембран, которые отделяют различные внутриклеточные органеллы. Мембраны состоят из двойного слоя молекул фосфолипидов, которые имеют гидрофильную полярную головку и гидрофобный углеводородный хвост (рисунок 7).
Взаимодействие с водным окружением приводит к ориентации молекул фосфолипидов в бислое полярной головкой наружу, а неполярными хвостами внутрь бислоя.
В биомембраны входят также некоторые другие липиды, как, например, холестерол, и множество белков, которые могут как свободно перемещаться в плоскости мембраны (плавать в жидкой мембране), так и быть жестко прикрепленными к внутри- или внеклеточному матриксу.
Цитозоль клетки и внеклеточное пространство существенно отличаются по солевому, ионному и аминокислотному составам. Свой собственный "микроклимат" имеют и клеточные органеллы, что позволяет реализовать "разделение труда" между ними.
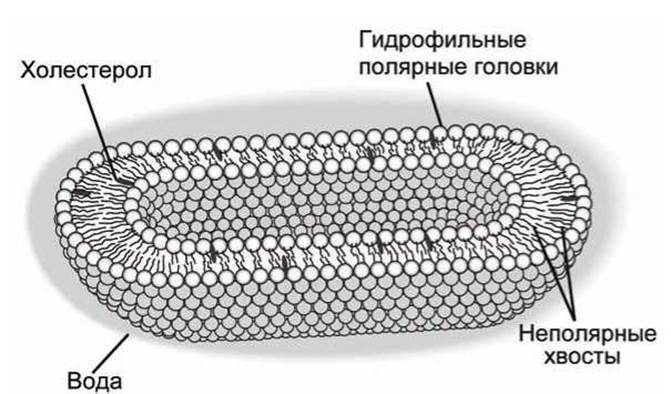
Рисунок 7 - Схема строения плазматической мембраны клетки
Можно образно сравнить клетку с фабрикой, а клеточные органеллы - с различными цехами этой фабрики. Все процессы синтеза на этой фабрике реализуются за счет энергии, предварительно извлеченной из сахаров, жиров или солнечного света и запасенной в АТФ. В клетках животных и растений большинство АТФ синтезируется в митохондриях и хлоропластах из АДФ и неорганического фосфата Рi (индекс "і" означает "inorganic"). Энергия ∆Е, выделяющаяся при гидролизе АТФ,
АТФ → АДФ + Рi + ∆Е
используется во многих энергозависимых клеточных процессах.
Вообще говоря, прямо или косвенно все питательные вещества в биосфере были созданы клетками растений с использованием солнечного света для синтеза сложных макромолекул в ходе фотосинтеза.
Для выполнения специфических функций клетке нужны специфические белки. Синтез белков происходит на рибосомах либо в цитозоле, либо в цистернах эндоплазматического ретикулума. Последнее характерно для белков, секретируемых клеткой, и для большинства мембранных белков. Дальнейшее "созревание" и модификация белков происходит в аппарате Гольджи.
Белки, которые перемещаются из эндоплазматического ретикулума в аппарат Гольджи, имеют специальные короткие последовательности аминокислот или присоединенные цепочки сахаров (олигосахариды), которые срабатывают как почтовый адрес, направляя белки к их месту назначения. Эти "адреса" распознаются и связываются другими белками, которые осуществляют сортировку и доставку транспортируемых белков в необходимый компартмент клетки.
Внутренний цитоскелет и внешние клеточные контакты придают клеткам специфическую форму. Цитоскелет животных клеток формируется из трех типов белков и представляет собой микрофиламенты или белковые нити, промежуточные волокна ж микротрубочки (рисунок 8).

Рисунок 8 - Микрофотографии элементов цитоскелета клетки: а - промежуточные волокна, б - микротрубочки, в - микрофиламенты
Цитоскелет участвует в движении клетки как целого, и во внутриклеточном транспорте везикул, хромосом и макромолекул. Цитоскелет через поверхность клетки может присоединяться к внеклеточному матриксу или цитоскелету других клеток, формируя тем самым специфическую ткань.
Все виды цитоскелета представляют собой длинные полимеры белковых субъединиц, и разборка и сборка этих полимеров в нужном месте клетки позволяет ей изменять форму, выпячивать и втягивать участки мембраны, тем самым перемещаться в нужном направлении со скоростью до 20 мкм/с. Движение отдельных клеток многоклеточного организма происходит:
1) во время развития эмбриона и формирования тканей,
2) при защите от инфекции,
3) при транспорте питательных веществ,
4) при залечивании ран.
Клетки растений не движутся поодиночке. Новые клетки появляются в результате деления уже существующих и развитие растения происходит за счет роста клеток, а не их перемещения с места на место.
Клетки постоянно получают информационные сигналы извне и реагируют на них соответствующим образом. Внешние сигналы могут быть в виде небольших химических молекул, белков, фотонов (света) и механического движения. Сигналом может быть как увеличение так и уменьшение интенсивности внешнего воздействия.
Способность клеток получать внешние сигналы и реагировать на них является критически важной как для дифференцировки клеток, так и для развития вообще. Причем ответный сигнал (реакция) клетки может варьироваться в зависимости от комбинации полученных сигналов.
Существует большое количество сенсорных клеточных систем. Условно их можно разделить на простые, когда реакция рецептора на внешний сигнал сразу передается активируемому белку, и сложные, когда внешний сигнал активирует экспрессию генов, что в свою очередь запускает белковый синтез (рисунок 9).
Связывание гормона или другой сигнальной молекулы с специфическим мембранным рецепторным белком может непосредственно запустить внутриклеточный процесс, который повышает или снижает активность ферментов (рисунок 9(a)).
Например, пища богатая углеводами приводит к повышению концентрации сахара в крови, что стимулирует выделение инсулина клетками поджелудочной железы в кровоток. Связывание инсулина с рецептором на мембране клеток печени или мышц приводит к активации фермента гликоген синтаза, который синтезирует гликоген из глюкозы, тем самым снижая уровень сахара в крови и запасая гликоген в клетках.
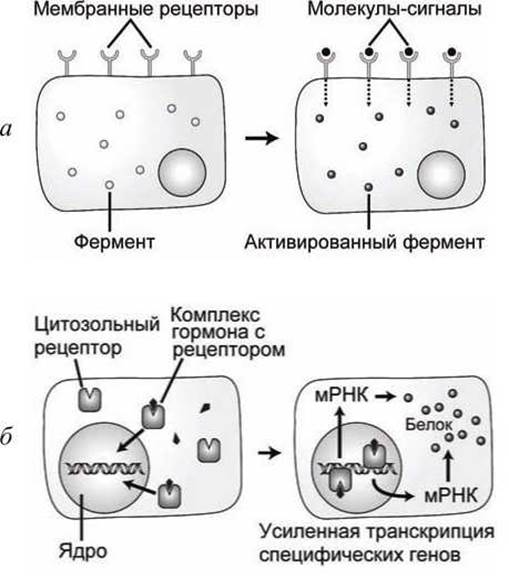
Рисунок 9 - Схемы сенсорных систем: а - простая; б - сложная
Рецепторы для стероидных гормонов расположены в цитозоле клетки, а не на внешней мембране (рисунок 9(б)). Комплекс рецептора с гормоном активирует транскрипцию специфических генов, что приводит к усилению синтеза специфических белков. Многие мембранные рецепторы также запускают сложные системы отклика клетки на внешнее событие, изменяя экспрессию генов.
Самым впечатляющим свойством клеток и целых организмов является, конечно же, их способность к воспроизводству. Биологическая репродуктивность в комбинации с эволюционной селекцией самых функционально эффективных качеств приводят к удивительной стабильности биологических видов.
Например, камчатский краб сегодня выглядит точно так же как и 300 миллионов лет назад, геологическое время, за которое в среднем возникает, растет, разрушается и исчезает с лица Земли горная система. Так, например, растущие Скалистые горы возникли только 10 миллионов лет назад, за это время сменилось 500 000 поколений крабов (продолжительность жизни краба порядка 19 лет). Нам кажется, что биологические структуры изменчивы, а геологические постоянны только потому, что продолжительность нашей собственной жизни коротка. Именно способность к воспроизводству обеспечивает то "бессмертие" биологическим структурам, которого нет у геологических объектов.
Простейший тип размножения представляет собой деление клетки на две дочерние, которое происходит в ходе клеточного цикла - последовательности событий, в ходе которых клетка готовится к делению, и собственно деления, которое называется митоз.
В клетках эукариот выделяют четыре фазы (этапа) деления (рисунок 10).
В ходе фазы S (synthesis) происходит репликация ДНК и набор хромосом удваивается. Реплицированные хромосомы разделяются в ходе фазы М (mitotic), во время которой каждая дочерняя клетка получает копию каждой хромосомы родительской клетки в ходе деления. Фазы М и S разделены паузами - этапами G1 и G2 (gap - щель, промежуток, "окно"), в ходе которых происходит активный синтез мРНК и белков.
В среднем клеточный цикл занимает 10-20 часов. В ходе интерфазы, которая включает S, G1 и G2 фазы, масса клетки практически удваивается. В митотической М-фазе хромосомы и содержимое клетки делятся поровну между дочерними клетками. В особых случаях таких, как голод или если ткань достигнет необходимого размера, клеточное циклирование прекращается и клетки остаются в "ждущем" режиме, называемом G0.
Большинство клеток при соответствующих условиях могут из G0-режима снова перейти в клеточный цикл.
В одноклеточных организмах обе дочерние клетки обычно (но не всегда) такие же, как родительская клетка. В многоклеточных организмах в результате деления стволовых клеток образуются две разные клетки, одна из которых подобна родительской (стволовой) а другая - нет.
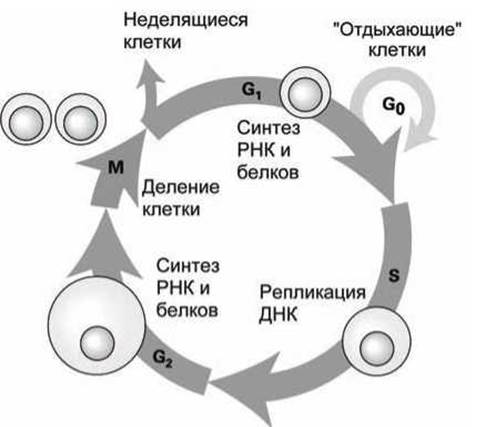
Рисунок 10 - Схема клеточного (митотического) цикла
Такой тип асимметричного деления позволяет дифференцироваться тканям.
Митоз представляет собой неполовой способ размножения, поскольку дочерняя клетка наследует точно тот набор генов, который был в родительской клетке.
При половом размножении происходит слияние двух клеток и образование третьей клетки, которая содержит генетическую информацию обеих родительских клеток.
Поскольку такое слияние удваивает число хромосом, то половой цикл размножения включает особый тип клеточного деления, мейоз, в ходе которого уменьшается число хромосом при подготовке к слиянию клеток (рисунок 11). Клетки с полным набором хромосом называются диплоидными.
В ходе мейоза диплоидные клетки один раз удваивают набор хромосом как при обычном митозе, но затем дважды осуществляют деление, не реплицируя ДНК между делениями.
В результате образуются четыре дочерние клетки, каждая из которых несет половинный набор хромосом, их называют гаплоидными.
Гаплоидные клетки животных - яйцеклетки и сперматозоиды — называют гаметами.

Рисунок 11 - Схема половой дифференциации при мейозе
Клетка человека, из которой образуются гаметы, имеет 46 хромосом: 44 неполовых хромосомы, которые называются аутосомы (autosomes) и 2 половые хромосомы X и Y.
В ходе мейоза образуются разные мужские гаметы с 22 аутосомами и одной (X или Y) половой хромосомой. Обе женские гаметы (яйцеклетки) одинаковы - они содержат 22 аутосомы и Х-хромосому. Слияние яйцеклетки и сперматозоида (оплодотворение) образует оплодотворенную яйцеклетку, зиготу, с 46 хромосомами, по 22 аутосомы от отца и матери плюс две Х-хромосомы в женской зиготе, либо одна X и одна Y в мужской зиготе.
Ошибки в мейозе приводят к патологиям вследствие ненормального числа хромосом в зиготе. Например, синдром Дауна вызван лишней хромосомой номер 21, а синдром Кляйнфельтера - лишней Х-хромосомой.
Поврежденная или инфицированная клетка умирает. Смерть клетки в результате такого "травматического" события сопровождается выделением из клетки токсичных или агрессивных веществ, что приводит к повреждению окружающих клеток.
Иначе развиваются события, когда клетка отмирает если она не получает сигналов о продолжении функционирования или, наоборот, получает сигнал об отмирании. Такая программируемая смерть клетки называется апоптоз. При апоптозе клетка сама синтезирует белки, необходимые для самоуничтожения, при этом не происходит выделения потенциально токсических веществ.
Программируемая клеточная смерть необходима для функционального развития организмов. Так, например, при внутриутробном развитии человеческого плода между пальцами сначала существуют перепонки. Клетки перепонок затем отмирают и у новорожденного пальцы свободны. Отмирают и нервные клетки, если они по каким-либо причинам утрачивают способность осуществлять электрические соединения с другими клетками.
Некоторые лимфоциты - клетки иммунной системы, предназначенные для распознавания чужеродных белков и полисахаридов, - могут также реагировать и с собственными тканями организма. Такие опасные для самого организма лимфоциты программируются для отмирания ещё до их созревания. Если же эти клетки не будут умерщвлены до окончательного созревания, они могут вызвать аутоиммунные заболевания, при которых иммунная система разрушает те ткани, которые она должна защищать.