Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Контроль генной экспрессии
Регуляция транскрипции прокариот
Синтез мРНК и соответственно синтез белка должны строго регулироваться, поскольку у клетки недостаточно ресурсов для одновременной транскрипции и трансляции всех структурных генов.
И прокариоты, и эукариоты постоянно синтезируют только те мРНК, которые необходимы для выполнения основных клеточных функций.
Экспрессия остальных структурных генов осуществляется под строгим контролем регуляторных систем, запускающих транскрипцию только в том случае, когда возникает потребность в определённом белке.
Само "решение" об инициации транскрипции того или иного гена является основой всего дальнейшего механизма белкового синтеза. С помощью механизма контроля инициации транскрипции клетка может регулировать свой белковый состав.
Репрессия транскрипции гена приводит к слабой транскрипции гена, при которой соответствующая мРНК синтезируется с низкой скоростью.
Активация транскрипции гена приводит к интенсивной транскрипции мРНК и, соответственно, к интенсивному синтезу соответствующего белка.
В большинстве бактерий и в других одноклеточных организмах интенсивность генной экспрессии эффективно регулируется, обеспечивая тонкую регулировку ферментативной системы и структурных компонентов, тем самым адекватно реагируя на изменения во внешней среде. В каждый данный момент бактериальная клетка синтезирует именно те белки, которые обеспечивают ей выживание в данных конкретных условиях.
В многоклеточных организмах контроль генной экспрессии главным образом рассчитан на обеспечение того, чтобы запустить экспрессию нужного гена в нужной клетке и в нужное время во время развития эмбриона и в ходе дифференцировки тканей.
У прокариот транскрипция инициируется связыванием РНК-полимеразы с последовательностями ТАТА и TTGAC премоторной области структурного гена или оперона.
В промоторе большинства структурных генов Е. coli имеются два сайта связывания для РНК-полимеразы. Один из них (ТАТА-бокс, или бокс Прибнова) обычно представляет собой нуклеотидную последовательность
![]()
а другой -
![]()
ТАТА-бокс и последовательность TTGAC расположены за 10 (область -10) и 35 (область -35) нуклеотидов до сайта инициации транскрипции соответственно (нуклеотид +1) (рисунок 19).

Рисунок 19 - Схема прокариотического оперона
Обычно от участка (о) между ТАТА-боксом и нуклеотидом +1 во многом зависит, будет ли происходить транскрипция данного оперона.
В зависимости от способа регуляции транскрипции оперона этот участок называется оператором или активатором.
Механизм контроля транскрипции в бактериях мы рассмотрим на примере lac оперона Е. coli.
В Е. coli около половины всех генов собраны в опероны, которые кодируют ферменты, участвующие в определённых метаболических путях или объединяющиеся после синтеза в один мультисубъединичный белок.
Например, trp оперон, рассмотренный выше, кодирует пять ферментов, необходимых для синтеза триптофана.
Аналогично, lac оперон содержит три гена: lac z, lac у и lac а, кодирующие три фермента: ß-галактозидаза, ß-галактозид-пермеаза и трансацилаза, участвующие в метаболизме лактозы (сахара, присутствующего в молоке, рисунок 20). Поскольку бактериальный оперон транскрибируется весь сразу, начиная с одной стартовой точки, в одну молекулу мРНК, то экспрессия всех генов оперона регулируется синхронно, они все или репрессируются, или активируются.

Рисунок 20 - Схема дисахарида лактоза состоящего из галактозы (слева) и глюкозы (справа)
Транскрипция оперонов (или изолированных генов, не входящих в опероны) контролируется взаимодействием между РНК-полимеразой и специфическими белками-репрессорами или белками-активаторами.
Кроме того, для инициации транскрипции РНК-полимераза Е. coli должна присоединить к себе один из небольших белков, которые называются σ-факторами, чаще всего с фактором σ70.
Пока Е. coli находится в среде с недостатком лактозы, синтез lac мРНК репрессирован, и ресурсы клетки не тратятся зря на синтез ферментов, которые не нужны в данный момент.
В среде, которая содержит и лактозу, и глюкозу, клетка Е. coli метаболизирует в основном глюкозу. Метаболизм лактозы включается на полную силу, только если лактоза доминирует в среде, а глюкоза практически исчерпана. Такая регулировка метаболизма достигается репрессией экспрессии lac оперона до тех пор, пока в среде не появится лактоза, и поддержанием синтеза lac мРНК на низком уровне, пока в среде не снизится концентрация глюкозы (рисунок 21).
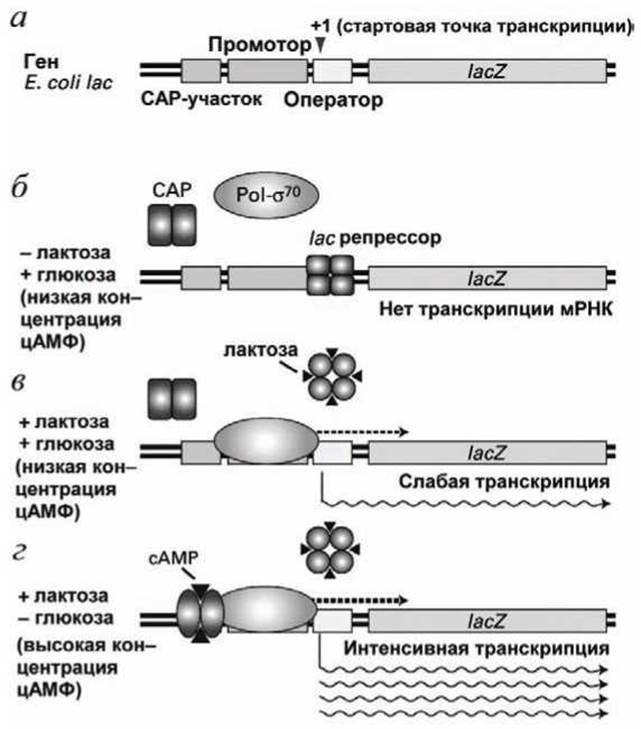
Рисунок 21 - Регуляция экспрессии lас оперона Е. coli
Транскрипция lас оперона управляется lас-репрессором и катаболитическим белком-активатором САР (catabolite activator protein), каждый из которых связывается со специфической последовательностью нуклеотидов ДНК в области, которая управляет транскрипцией lac оперона (рисунок 21(a)).
Для того, чтобы началась транскрипция lac оперона субъединица σ70 РНК-полимеразы должна связаться с lac промотором, который находится сразу перед стартовой точкой.
Когда в среде нет лактозы, lас-репрессор связывается с lac оператором, который расположен сразу за промотором, даже слегка перекрываясь с ним.
Связывание lас-репрессора с lac оператором блокирует промотор так, что даже посадка РНК-полимеразы на промотор становится невозможной (рисунок 21(6)). Когда же в среде есть лактоза, она связывается со специфическими центрами связывания на каждом из четырёх субъединиц lас-репрессора, индуцируя такое его конформационное изменение, вследствие которого lас-репрессор диссоциирует с lac оператора. В результате РНК-полимераза может инициировать транскрипцию lac оперона (рисунок 21(b)).
В то же время, пока в среде присутствует глюкоза, скорость инициации транскрипции (т.е. число актов инициации транскрипции в единицу времени) крайне мала и, следовательно, синтезируется малое число мРНК и белков, закодированных в lac опероне.
Стоит только убрать глюкозу из среды, что сопровождается снижением концентрации глюкозы внутри клетки, Е. coli синтезирует циклический аденозинмонофосфат, цАМФ (cyclic adenosine monophosphate, сАМР) (рисунок 22).
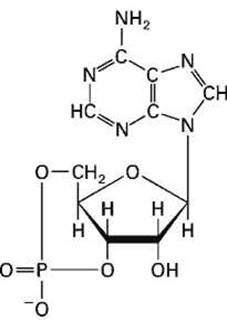
Рисунок 22 - Циклический аденозинмонофосфат, цАМФ
С ростом концентрации цАМФ он связывается с каждой из субъединиц димерного САР-белка, индуцируя такую его конформацию, которая позволяет этому белку связаться с САР-участком ДНК в области, которая управляет транскрипцией lac оперона (рисунок 21 (г)).
Комплекс "САР-сАМР" взаимодействует с РНК-полимеразой, связанной с промотором, существенно стимулируя скорость инициации транскрипции (минимизируя абортивную инициацию). Такая активация ведет к интенсивной транскрипции мРНК и, следовательно, к интенсивному синтезу ферментов закодированных в lac опероне.
Как только ферменты переработают всю лактозу, свободный от лактозы репрессор выключит lac оперон.
Хотя структура промоторов различных генов Е. coli подобна (они являются гомологичными), но точная последовательность нуклеотидов в них различна. Именно последовательность нуклеотидов в промоторе определяет скорость инициации транскрипции в отсутствии репрессоров и активаторов.
Промоторы, которые поддерживают высокую скорость инициации транскрипции, называются сильными промоторами (strong promoters).
Промоторы, которые поддерживают низкую скорость инициации транскрипции, называются слабыми промоторами (weak promoters). Так lac оперон это пример слабого промотора, присущая ему низкая скорость инициации транскрипции ещё более снижается lас-репрессором или же существенно увеличивается САР-сАМР-активатором.