Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Контроль генной экспрессии
Регуляция транскрипции эукариот
У эукариот РНК-полимераза II, которая транскрибирует структурные гены, связывается с целым набором белков - факторов транскрипции, которые последовательно присоединяются к ТАТА-последовательности промоторной области.
Несмотря на индивидуальность набора регуляторных элементов у структурных генов эукариот, каждый из них имеет:
1) промоторный участок (ТАТА-бокс, или бокс Хогнесса) из восьми нуклеотидов, включающий последовательность ТАТА;
2) последовательность ССААТ (САТ-бокс);
3) участок из повторяющихся динуклеотидов GC (GC-бокс).
Эти элементы находятся на расстоянии 25, 75 и 90 bp от сайта инициации соответственно (рисунок 27) (bp или п. н. обозначает комплементарную пару нуклеотидов, base pair).
Знаки «-» при числах на рисунке 27 означают, что эти элементы находятся в молекуле ДНК слева от сайта инициации транскрипции (+1) (выше начала транскрипции). Стрелка обозначает направление транскрипции. Положение регуляторных элементов и их размер даны без соблюдения масштаба.

Рисунок 27 - Регуляторные элементы структурных генов эукариот
За включение и выключение транскрипции отвечают дополнительные факторы транскрипции, которые связываются с соответствующими участками ДНК.
Транскрипция структурного гена эукариот начинается со связывания с ТАТА-боксом фактора транскрипции IID (transcription factor IID, TFIID), который представляет собой комплекс, по крайней мере, из 14 субъединиц, одна из которых называется ТАТА-связывающий белок ТВР (TATA-bindingprotein, рисунок 3) (рисунок 28(a)).
Затем с TFIID и участками ДНК, примыкающими к ТАТА-боксу, связываются другие факторы транскрипции (TFIIA, TFIIB, TFIIE, TFIIF, TFIIH (рисунок 28(6)), и, наконец, со всем этим мультибелковым комплексом связывается РНК-полимераза II. Фактор TFIIF правильно позиционирует РНК-полимеразу II на начало транскрибируемого гена.
Весь этот белковый ассоциат, который закреплен на ТАТА-боксе, включая РНК-полимеразу II называется транскрипционный прединициаторный комплекс (transcription preinitiation complex).
Необходимо отметить, что промоторы некоторых генов не имеют ТАТА-бокса. Такие гены обычно кодируют те белки, которые нужны данной клетке для "внутриклеточного" использования. Такие гены называются housekeeping genes. В этом случае специальный белок Spl связывается с последовательностью 10 нуклеотидов GGGGCGGGGC в GC-боксе и присоединяет к себе белок ТВР (ТАТА-связывающий белок), хоть для него и нет ТАТА-бокса в составе гена. А затем уже вокруг этого белка ТВР ассемблируется весь транскрипционный прединициаторный комплекс, который и начинает транскрипцию.
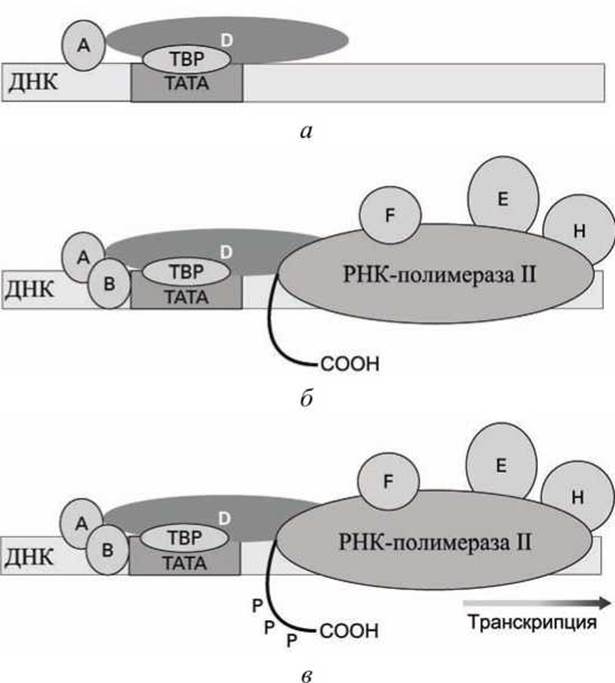
Рисунок 28 - Эукариотический комплекс инициации транскрипции
В любом случае инициация транскрипции происходит, когда фосфорилируется домен РНК-полимеразы II на С-конце полимеразы. Фосфорилирование этого домена приводит к отрыву РНК-полимеразы II от прединициаторного комплекса, и начинается транскрипция ДНК в РНК в точке +1 (рисунок 28(b)).
Хотя ассемблирование транскрипционного прединициаторного комплекса обычно достаточно для синтеза РНК, однако интенсивность транскрипции может быть значительно увеличена вследствие связывания с ДНК активаторных белков вблизи начала гена. Такие активаторные белки тоже называют факторами транскрипции, а те последовательности нуклеотидов в молекуле ДНК, к которым они присоединяются, называются энхансерными последовательностями. Интересно, что энхансерные последовательности могут располагаться как выше начала транскрипции гена - в области «-», так и ниже (хотя и намного реже) - в области «+».
Использованием таких транскрипционных факторов и энхансерных последовательностей определяется, какие именно гены, и на каком этапе клеточного цикла будут транскрибироваться. Например, на рисунке 29 представлена схема активации транскрипции гена миозина в скелетных мышцах и дезактивация этого же гена в гепатоцитах (клетках печени) с помощью наличия (или отсутствия) в клетках ткани данного типа соответствующего активаторного белка.

Рисунок 29 - Транскрипция гена в разных тканях
Гормональная регуляция экспрессии генов. В качестве примера, иллюстрирующего способы управления транскрипцией эукариотических генов, рассмотрим регуляцию транскрипции некоторых генов глюкокортикоидными гормонами, которые являются стероидными гормонами, которые вырабатывают клетки коры надпочечников (рисунок 30).
Эти гормоны могут диффундировать сквозь клеточную мембрану, поскольку они незаряжены и неполярны. В цитозоле находятся белковые факторы транскрипции, которые имеют сайты связывания со стероидными гормонами, и которые, поэтому, называются рецепторами стероидных гормонов.

Рисунок 30 - Регуляция транскрипции стероидными гормонами
Когда в цитозоли нет глюкокортикоидных гормонов, то белки- рецепторы неактивны, поскольку они связаны с ингибиторными белками Hsp90, и не могут проникнуть в ядро через ядерные поры. Однако когда в цитозоле появляются глюкокортикоидные гормоны, которые связываются с соответствующими сайтами рецепторов, то белки Hsp90 отсоединяются от рецепторов, и комплекс гормона с рецептором может быть перенесен через ядерную мембрану в ядро. В ядре два таких комплекса могут связаться с 15-bр последовательностью нуклеотидов, которая расположена выше ТАТА-бокса и называется гормон-распознающим элементом, HRE (hormone response element). HRE является энхансерной последовательностью. Комплекс глюкокортикоидного рецептора с гормоном взаимодействует с прединициаторным комплексом, присоединённым к ТАТА-боксу, в результате чего резко возрастает скорость транскрипции гена, имеющего HRE последовательность.
На рисунке 31 показано, почему рецептор глюкокортикоидных гормонов связывается с ДНК как димер.
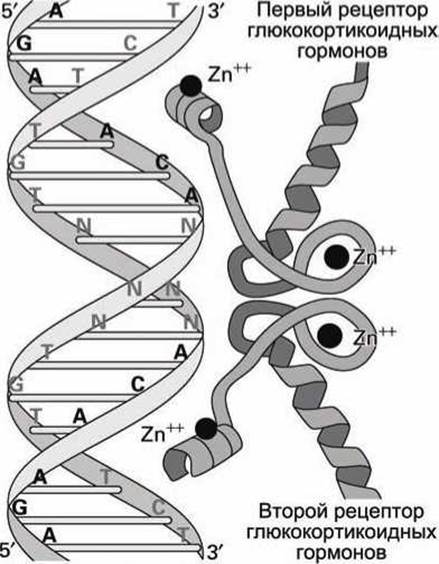
Рисунок 31 - Палиндромный гормон-распознающий элемент (HRE) ДНК, связанный с димеризованными рецепторами стероидных гормонов
HRE является палиндромом, то есть участком ДНК, обе нуклеотидные нити которого одинаковы, если каждую из них читать в направлении 5'→3'. Для нашего примера HRE имеет вид
![]()
Каждая из нитей HRE содержит шестинуклеотидную последовательность AGAACA, которая называется мотив распознавания кора (core recognition motif). С этой последовательностью связывается одна белковая молекула рецептора глюкокортикоидных гормонов. Поскольку HRE содержит два таких мотива, то к HRE присоединяются два рецептора. Две 6-ти нуклеотидные последовательности разделены тремя парами оснований (обозначены NNN на рисунке 31), которые нужны для того, чтобы обеспечить достаточно пространства для того, чтобы гомодимер рецепторов мог связаться с HRE. Эти три пары оснований могут быть любыми, поскольку они не влияют на прочность связывания с рецепторным комплексом.
Этот пример иллюстрирует механизм гормональной регуляции транскрипции. В общем случае гормон, поступив в кровоток, связывается с рецепторами специфических клеток, облегчающими его проникновение в клетку. Оказавшись в клетке, гормон вступает во взаимодействие с одним из клеточных белков и изменяет его конформацию. В таком измененном состоянии белок проникает в ядро и связывается со специфическим регуляторным элементом, который инициирует транскрипцию соответствующего гена.
ВЫВОДЫ
Генная экспрессия и прокариот, и эукариот регулируется главным образом механизмами, которые контролируют инициацию транскрипции.
Связывание комплекса а-РНК-полимераза с промотором является первым этапом в инициации транскрипции Е. coli.
Последовательность нуклеотидов в промоторе определяет его силу - насколько часто (в единицу времени) разные молекулы РНК-полимеразы могут связываться с промотором и инициировать транскрипцию.
Репрессоры - это белки, которые связываются с оператором. Оператор - это участок ДНК, который расположен рядом с промотором или даже перекрывает промотор. Связывание репрессора с оператором ингибирует инициацию транскрипции.
Активность связывания репрессоров с ДНК регулируется индукторами - малыми молекулами-эффекторами. Это позволяет бактериальной клетке регулировать транскрипцию специфических генов в ответ на изменения концентрации различных компонентов в окружающей среде.
lас-Оперон и некоторые другие бактериальные гены также регулируются белками-активаторами, которые связываются вблизи промотора и увеличивают скорость инициации транскрипции РНК-полимеразы.
Основной сигма-фактор у Е. coli это σ70, но существуют и некоторые другие менее распространенные сигма-факторы, для каждого из которых существует своя промоторная последовательность нуклеотидов.
Инициация транскрипции всеми РНК-полимеразами Е. coli, кроме тех, которые содержат σ54, могут регулироваться репрессорами и активаторами, которые связываются с ДНК вблизи стартовой точки транскрипции.
Гены, транскрибируемые σ54-РНК-полимеразой, регулируются активаторами, которые связываются с энхансером, расположенным более чем за 100 нуклеотидов до стартовой точки. ДНК образует петлю, для того чтобы сблизились активатор и σ54-РНК-полимераза.
В двухкомпонентных регуляторных системах один из белков выполняет функцию сенсора, отслеживающего концентрацию питательных веществ или других компонентов в окружающей среде. При соответствующих условиях у-фосфат АТФ переносится сначала на гистидин белка- сенсора, а затем на аспарагиновую кислоту во втором белке, который называется регулятором отклика. Фосфорилированный регулятор отклика затем связывается с регуляторной последовательностью нуклеотидов ДНК, стимулируя или репрессируя транскрипцию специфических генов.
В эукариотах РНК-полимераза II использует ТАТА-связывающий белок и другие белковые факторы транскрипции для связывания с промотором. Этот набор белков называется транскрипционный прединициаторный комплекс, и его достаточно для синтеза РНК. Для интенсификации транскрипции в ответ на внешний сигнал (такой, как, например, гормон) используются другие белки, которые присоединяются к энхансерным последовательностям. Эти белки взаимодействуют с прединициаторным комплексом и повышают интенсивность транскрипции.
Вопросы для самоконтроля
1. Что такое оператор в laс-опероне, и какова его функция?
2. Какое вещество является индуктором lас-репрессора?
3. Какова роль цАМФ в регуляции транскрипции laс-оперона?
4. Какова роль САР-белка в регуляции транскрипции lас-оперона?
5. Связывание с каким веществом позволяет САР-белку присоединиться к ДНК и активизировать транскрипцию laс-оперона?
6. Что называется индуктором и какова функция индукторов?
7. В чем сходство и различие транскрипции генов РНК-полимеразами с присоединенными сигма-факторами σ70 и σ54?
8. Что называется энхансером?
9. Охарактеризуйте роль обоих компонентов в двухкомпонентных системах регуляции.
10. Какие факторы транскрипции входят в транскрипционный прединициаторный комплекс эукариот?
11. Охарактеризуйте механизм регуляции транскрипции стероидными глюкокортикоидными гормонами.