Основы молекулярной биологии - В.И. Резяпкин 2009
Нуклеиновые кислоты
Вторичная структура ДНК
В 1953 г. американский генетик Д.Уотсон и английский физик Ф.Крик предложили модель вторичной структуры. Согласно этой модели ДНК по своей пространственной организации представляет собой двойную спираль.
Свое открытие ученые сделали, основываясь на ранее полученных результатах других ученых. Так Э.Чаргафф и более поздние исследователи, изучая нуклеотидный состав ДНК различных видов организмов, сделали следующие выводы:
a) нуклеотидный состав ДНК разных тканей одного и того же вида одинаков;
b) нуклеотидный состав ДНК у разных видов различен;
c) нуклеотидный состав не зависит от возраста и питания;
d) в составе ДНК число остатков аденина всегда равно числу остатков тимина, а число остатков гуанина равно числу остатков цитозина.
Другие ученые Р.Франклин и М.Уилкинс опубликовали рентгенограмму, полученную при рентгеноструктурном анализе ДНК. Метод рентгеноструктурного анализа широко используется при исследовании пространственной организации молекул. Предложенная Уотсоном и Криком двойная спираль ДНК (рис. 1.2) объяснила результаты исследований выше упомянутых ученых.
Ниже представлены параметры двойной спирали ДНК, предложенной Уотсоном и Криком:
a) ДНК состоит из двух цепей, закрученных в правую двойную спираль (рис. 1.2Б.):
b) цепи в молекуле ДНК расположены относительно друг друга антипараллельно (рис. 1.2А.);
c) молекулы азотистых оснований ориентированы перпендикулярно оси двойной спирали;
d) на внешней стороне двойной спирали находятся остатки пентозы и фосфорной кислоты;
e) цепи ДНК (рис. 1.2Б) при закручивании в двойную спираль образуют большую и малую борозды, ширина большой борозды - 2,2 нм, малой - 1,2 нм;
f) на один виток спирали приходится 10 пар нуклеотидных остатков;
g) полный виток спирали имеет длину 3,4 нм;
h) диаметр двойной спирали 1,8 нм;
i) цепи ДНК связаны друг с другом водородными связями, которые образуются между гуанином одной цепи и цитозином другой цепи, или между тимином и аденином, расположенными в разных цепях;
j) между тимином и аденином образуются две водородные связи, а между гуанином и цитозином - три водородные связи (рис. 1.3.);
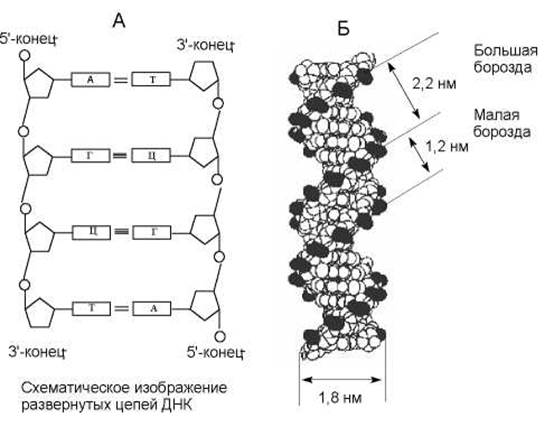
Рис. 1.2. Параметры ДНК
Способность гуанина взаимодействовать в молекуле ДНК только с цитозином, а аденина - только с тимином называют комплементарностью, а основания гуанин и цитозин, аденин и тимин - комплементарными. Согласно принципу комплементарности, последовательность одной цепи будет определять последовательность другой цепи. Всегда против аденина будет находится тимин, а против гуанина - цитозин. Таким образом, цепи ДНК в двойной спирали будут комплементарны друг другу.
Двойная спираль стабилизируется также стэкинг - взаимодействиями между основаниями. Основания расположены друг над другом и сближены своими плоскостями. В результате между ними возникают гидрофобные взаимодействия, а также дипольные взаимодействия п-связей.
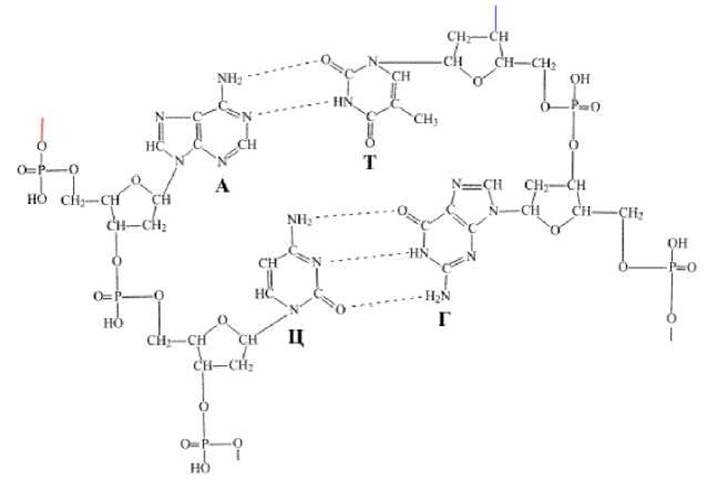
Рис. 1.3. Образование водородных связей между аденином и тимином, гуанином и цитозином в молекуле ДНК
Параметры двойной спирали в зависимости от условий и состава ДНК могут несколько отличатся от той модели, которую предложили Уотсон и Крик. В настоящее время описаны и другие модели ДНК. Тем не менее, во всех предложенных моделях сохраняется принцип комплементарности, и цепи ДНК закручены в двойную спираль.
A-форма ДНК
A-форма ДНК (рис. 1.4) образуется при относительно низкой влажности. Эта структура является правой спиралью. В этой форме ДНК на виток спирали приходится 11 пар оснований. Расстояние между нуклеотидами вдоль оси спирали составляет 2,56 А. Пары оснований наклонены на 20o.
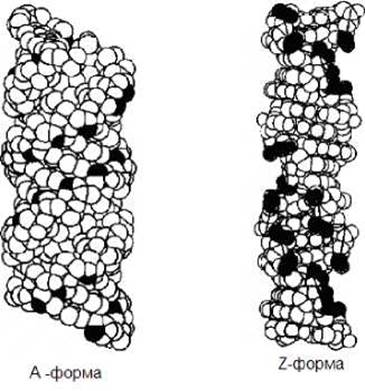
Рис. 1.4. Различные формы ДНК
С- форма ДНК
С-ДНК образуется при высокой концентрации соли и значениях влажности, промежуточных между теми, при которых образуются А- и В-ДНК. Шаг спирали С-ДНК равен 30,9 А. Число пар оснований на виток составляет 9,33. Пары оснований наклонены на угол - 8о относительно оси.
Z-форма ДНК
Z-форма ДНК - левоспиральная (рис. 1.4). Она была открыта в 1979 г при исследовании структуры гексануклеотида d(CG)3. У этой спирали сохраняется уотсон-криковское спаривание оснований. На виток Z-спирали приходится 12 пар оснований.
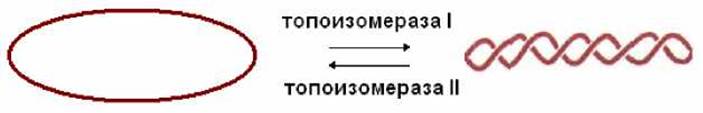
Кольцевые двухцепочечные молекулы ДНК могут находиться в состоянии суперспирализации. Суперспирализация может возникнуть при локальном расплетении двойной спирали. Возникшее в результате напряжение снимается суперспирализацией. Переход кольцевой молекулы ДНК в суперспирализованное состояние и обратно осуществляется при участии топоизомераз:
В молекуле ДНК могут присутствовать обращенные повторы - палиндромы:
![]()
Палиндромы могут образовывать крестообразные структуры:
