Основы молекулярной биологии - В.И. Резяпкин 2009
Организация гемонов
Репликация геномов ДНК-содержащих вирусов
В репликации геномов ДНК-содержащих вирусов принимает большое число белков: ДНК-полимеразы, праймазы, лигазы, хеликазы, белки, обеспечивающие инициацию репликации, и др. Некоторые из них являются ферментами, другие - нет. Упомянутые белки могут быть как вирусного, так и клеточного происхождения. Например, у фага φХ174 - все белки, участвующие в репликации, за исключением белка А имеют клеточное происхождение, в то время как у фага Т-4 - вирусное происхождение. Известны случаи, когда белки, принимающие участие в репликации, имеют смешанное происхождение. Одни субъединицы таких белков закодированы в клеточном геноме, другие - в вирусном.
Репликация генома аденовирусов
Роль затравки при репликации ДНК-генома аденовирусов выполняет комплекс вирус-специфического терминального белка и одного нуклеотида, присоединенного ковалентно (рис. 11.2). После взаимодействия компклеса осуществляется синтез дочерней цепи. В процессе ее синтеза происходит вытеснение из дуплекса материнской цепи. К 3’-концу освободившейся цепи присоединяется нуклеотид-белковая затравка и начинается синтез комплементарной цепи. В результате образуется двухцепочечная молекула ДНК.
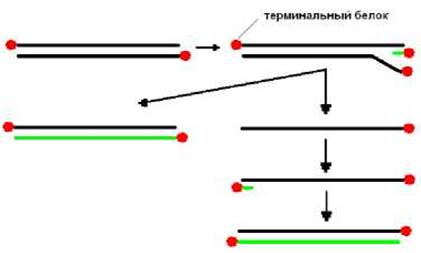
Рис. 11.2. Схема репликации аденовирусов Репликации генома SV40
Геном SV-40 представлен двухцепочечной кольцевой ковалентнозамкнутой молекулой ДНК.
После взаимодействия вирус-специфической хеликазы (Т-антиген) с точкой начала репликации праймаза образует затравки на обеих родительских цепях. Далее происходит синтез дочерних цепей. Возникающие при раскручивании ДНК напряжение снимается топоизомеразой I. В результате репликации образуются две сцепленные молекулы ДНК, которые разделяются при участии топоизомеразы II. (рис. 11.3). Все белки, участвующие в репликации вирусной ДНК, за исключением Т-антигена имеют клеточное происхождение.

Рис. 11.3. Схема репликации генома SV40
Репликации фага λ
Геномная НК фага λ представлена двухцепочечной линейной ДНК с липкими концами. В клетке линейная ДНК фага λ за счет липких концов образует кольцо и превращается в ковалентнозамкнутую форму. Точка начала репликации распознается фагоспецифическими белками. Затем клеточная праймаза осуществляет синтез затравки и происходит синтез дочерних цепей. На следующей стадии происходит разрыв одной из материнских цепей ДНК. В результате образуется, так называемая, σ-молекула ДНК. На поздних стадиях репликации молекула ДНК содержит множественные копии фагового генома. На заключительной стадии фагоспецифическая эндонуклеаза вносит ступенчатые разрывы в строго определенных местах, в результате образуются зрелые геномные молекулы ДНК фага λ (рис. 11.4).

Рис. 11.4. Схема репликации фага λ Репликация фага Т-7
ДНК фага Т-7 - линейная двухцепочечная молекула (40 т.п.н.) с прямыми концевыми повторами. Инициация синтеза ДНК происходит внутри молекулы и протекает в двух направлениях. В результате образуются две молекулы ДНК с недостроенными 5’-концами. 3’-концы этих молекул комплементарны и между ними может возникнуть комплементарное взаимодействие (рис. 5) и затем цепи ДНК могут быть ковалентно соединены. На следующей стадии вносится ступенчатый разрыв. Вследствие этого образуются две молекулы ДНК с недостроенными 3’-концами, которые затем достраиваются (рис. 11.5). Таким образом, образуются полномерные геномные ДНК.

Рис. 11.5. Схема репликации фага Т-7
Репликация генома вируса гепатита В
ДНК вируса гепатита В образована полномерной (-) цепью ДНК (около 3,2 т.н), к 5’-концу которой присоединен белок, и ей комплементарной более короткой (+) цепью (1,7 - 2,8 т.н). (+) Цепь комплементарна обоим концам (-) цепи, и поэтому замыкает (-) цепь в кольцо (рис. 11.6.).
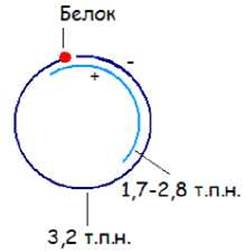
Рис. 11.6. Строение ДНК вируса гепатита В
В вирионе присутствует вирус-специфическая ДНК-полимераза, способная достраивать (+) ДНК, при ее участии в зараженной клетке ДНК вируса переходит в ковалентно замкнутую циклическую форму (рис. 11.7).

Рис. 11.7. Образование двухцепочечной ковалентно замкнутой ДНК
РНК-полимераза II, используя циклическую двухцепочечную ДНК в качестве матрицы, синтезирует два типа +РНК:
a) короткие субгеномные +РНК, служащие матрицами для синтеза белка;
b) длинные +РНК, включающие весь геном вируса.
Длинные +РНК служат матрицами для синтеза ДНК посредством вирус-специфической ревертазы. Сначала синтезируется (-) цепь ДНК. В качестве затравки ревертаза использует белок. Затем этот же фермент использует (-) цепь ДНК для синтеза (+) цепи ДНК.
Репликация фага φX174
Геном фага φХ174 представлен одноцепочечной кольцевой ковалентнозамкнутой молекулой (+)ДНК. После проникновения в клетку он превращается при участии клеточных праймазы, ДНК-полимераз I и III, лигазы в кольцевую двухцепочечную ковалентнозамкнутую молекулу ДНК. Затем фагоспецифический белок вносит разрыв в специфическом участке цепи (+)ДНК и присоединяется ковалентно к образовавшемуся при разрыве 5’-концу. Возникшей же при разрыве (+) цепи 3’-конец используется клеточной ДНК-полимеразой в качестве затравки, при этом 5’-конец вытесняется из дуплекса и обнажившаяся цепь (-)ДНК выступает в качестве матрицы. После того как репликативная вилка сделает чуть больше одного оборота, фагоспецифический белок вносит разрыв в (+) цепи на границе генома и присоединяется к 5’-концу (+)цепи ДНК в составе дуплекса. Вытесненная (+) цепь ДНК замыкается в кольцо (рис. 11.8). Освободившиеся двухцепочечная ДНК вновь вовлекается в процессы синтеза геномной (+) ДНК. В тоже время образовавшаяся кольцевая +ДНК может быть превращена в двухцепочечную ДНК и принять участие в синтезе новых (+) ДНК или войти в состав вириона.
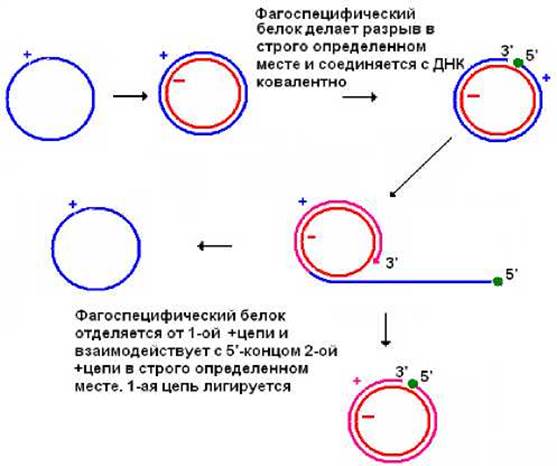
Рис. 11.8. Схема репликации фага φХ174
Схема репликации парвовирусов
Геном парвовирусов представлен одноцепочечной линейной ДНК. Оба конца цепи ДНК способны образовывать шпильки, благодаря присутствию самокомплементарных последовательностей.
Рассмотрим репликацию аденоассоциированного вируса (его репродукция возможна в присутствии аденовируса).
При попадании в клетку 3’-конец вирусной ДНК образует шпильку, которая выступает в качестве затравки (рис. 11.9). Затем происходит синтез комплементарной цепи ДНК. В результате такого синтеза осуществляется воспроизведение не полного генома вируса. Далее вирус-специфический белок вносит разрез в материнскую цепь. Образовавшийся 3’-конец используется в качестве затравки для синтеза недостающего фрагмента генома. В итоге образуется двухцепочечная молекула ДНК. Далее на обеих цепях одного из концов молекулы ДНК происходит образование шпилек («заячьи уши»), возникает затравка, и начинается синтез комплементарной цепи. При этом будет происходить вытеснение цепи ДНК, идентичной геному. Образовавшаяся двухцепочечная ДНК может быть использована далее для синтеза геномной одноцепочечной ДНК по аналогичному механизму (рис. 11.9).

Рис. 11.9. Схема репликации генома аденоассоциированного вируса