Основы молекулярной биологии - В.И. Резяпкин 2009
Организация гемонов
Организация генома прокариот
Геном прокариот может состоять из одной или нескольких крупных молекул ДНК, называемых хромосомами, и небольших молекул ДНК - плазмид. В хромосомах представлены практически все гены, необходимые для жизнедеятельности бактерии. Плазмиды же несут гены, необязательные для бактерии, без них клетка может обойтись, хотя в некоторых условиях они способствуют ее выживанию.
Хромосомы и плазмиды могут представлять собой как кольцевые, так и линейные двухцепочечные молекулы ДНК. Геном бактерий может состоять из одной или нескольких хромосом и плазмид (таблица 11.2). Хромосома (ы) в бактериальной клетке представлена (ы) в виде одной копии, т.е. бактерии гаплоидны. Плазмиды же могут присутствовать в клетке как в виде одной копии, так и в нескольких. Более подробно плазмиды были рассмотриваются в разделе «Мобильные генетические элементы». В этом же разделе говорится о таких важных компонентах генома прокариот, как IS-последовательностях и транспозонах.
Таблица 11.2.
Состав бактериальных геномов
|
Бактерии |
Геном |
|
Streptomyces ambofaciens |
1 линейная хромосома |
|
Agrobacterium tumefaciens C58 |
1 линейная хромосома, 1 кольцевая хромосома, 2 плазмиды |
|
Bacillus cereus |
1 кольцевая хромосома, 1 мегаплазмида |
|
Brucelia melitensis |
2 кольцевые хромосомы |
|
Rodobacter sphaeroides |
2 кольцевые хромосомы |
|
Rodococcus facians |
1 линейная хромосома, 1 линейная плазмида |
Хромосома уложена в компактную структуру - нуклеоид, который имеет овальную или сходную с ней форму. Его структура поддерживается ДНК-связывающими гистоноподобными белками и молекулами РНК. С нуклеоидом также ассоциированы молекулы РНК-полимеразы и ДНК-топоизомеразы I. По периферии нуклеоида располагаются петли хромосомной ДНК, которые находятся в транскрипционно активном состоянии. При подавлении транскрипции эти петли втягиваются внутрь. Нуклеоид не является стабильным образованием и во-время различных фаз роста бактериальных клеток изменяет свою форму. Изменение его пространственной организации сопряжено с изменением транскрипционной активностью определенных генов бактерий.
Впервые в 1995 г была определена полная последовательность нуклеотидов хромосомы паразитической бактерии Haemophilus influenzae. Затем в 1997 г. установлена первичная структура хромосом Escherichia coli и Bacillus subtilis. К настоящему времени расшифрованы геномы более 100 видов бактерий.
В состав хромосомы могут входить геномы умеренных фагов. Включение их геномов в клеточный может происходить после заражения фагами бактерий. При этом одни фаговые геномы интегрируют в строго определенные участки хромосомы, другие - в участки различной локализации. При определенных условиях геном фага может выщепляться из хромосомы, формируя полноценные фаговые частицы. В некоторых случаях фаговый геном остается навсегда включенным в хромосому, и тогда его ДНК, может служить материалом для возникновения новых бактериальных генов. Как оказалось, некоторые гены бактерий на самом деле являются фаговыми.
Размер геномов прокариот колеблется от нескольких сотен тысяч до десятка миллионов пар нуклетидов (таблица 11.3). Например, размер хромосомы E.coli составляет 4,6*106 п.н. Ее относительная молекулярная масса равна 3*109, а длина - 1,5 мм. Хромосома собрана в нуклеоид с параметрами 1*1*2 мкм. Время репликации хромосомы Escherichia.coli - около 20 мин. Репликация ДНК других бактерий может осуществляться с меньшей скоростью. Так, туберкулезная микобактерия делится всего лишь один раз в сутки. Подробнее вопрос о механизме репликации прокариот рассматривается в разделе «Репликация».
Геномы прокариот отличаются друг от друга по содержание ГЦ-пар, их доля в их составе колеблется от 23 до 72 %. Интересно, что в ДНК бактерий, обитающих при высоких температурах, содержание этих нуклеотидов повышено. Их преобладание над АТ-парами обуславливает более высокую температуру плавления ДНК, что является жизненно необходимым фактором для таких бактерий. Нужно отметить, что в белках термофильных бактерий повышено также и содержание полярных аминокислот, что делает их более устойчивыми к денатурации при повышенных температурах. В составе белков хеликобактерий (обитающих в кислой среде) больше аминокислотных остатков аргинина и лизина. Остатки этих аминокислот способны связывать ионы водорода, тем самым, оказывая влияние на кислотность среды, и способствуя выживанию бактерий в сложных экологических условиях.
На долю кодирующих последовательностей генома у прокариот приходится около 90 %, остальные 10 % генома - на регуляторные последовательности. При этом общее число генов в геномах прокариот составляет от нескольких сотен до нескольких тысяч (таблица 11.3).
Таблица 11.3
Характеристика некоторых бактериальных геномов
|
Бактерии |
Размер генома, т.п.н. |
Кодирующие последовательности, % |
Предположительное число генов |
|
Mycoplasma genitalium |
580 |
90 |
479 |
|
Mycoplasma pneumoniae |
817 |
89 |
677 |
|
Borrelia burgdoferii |
1300 |
93 |
863 |
|
Helicobacter pylory |
1700 |
91 |
1590 |
|
Bacillus subtilis |
4200 |
87 |
4000 |
|
Mycobacterium tuberculosis |
4400 |
91 |
3924 |
|
Escherichia coli |
4600 |
89 |
4288 |
О числе генов в геноме судят по наличию в их составе открытых рамок считывания (ОРС). ОРС представляет собой полинуклеотидную последовательность, потенциально способную кодировать полипептид. О существовании ОРС на тех или иных участках ДНК судят на основании расшифрованной первичной структуры ДНК. Основным критерием принадлежности участка полинуклеотидной цепи к ОРС служит отсутствие стоп-кодонов на достаточно протяженном участке после стартового кодона. В то же время наличие ОРС является недостаточным условием для утверждения о наличии на данном участке ДНК гена. Это связано с тем, что для эффективной транскрипции и последующей трансляции необходимы регуляторные последовательности: промотор, оператор, аттенюатор, последовательность Шайна-Дальгарно и др. Чтобы определить, является ли ОРС реальными генами, сравнивают аминокислотные последовательности, предположительно закодированных в соответствующих ОРС белков с последовательностями аминокислотных остатков уже известных белков. Таким образом, соответствие каждой ОРС определенному гену нужно еще установить. Например, у E. coli установлены функции около 60 % ОРС.
Гены, прокариот, как правило, имеют оперонную организацию. В одном опероне обычно представлены гены, ответственные за осуществление одного и того же метаболического процесса. Подробнее организация прокариотических генов и регуляция их экспрессии рассматриваются в разделах «Молекулярная организация генов», «Регуляция экспрессии генов», «Транскрипция», «РНК, процессинг РНК». Следует отметить, что у паразитических бактерий могут отсутствовать гены, ответственные за синтез веществ, поступающих из среды обитания.
В бактериальном геноме содержатся гены, полинуклеотидные последовательности которых схожи. Их называют гомологичными генами. Причиной появления гомологичных генов может являться дупликация какого-либо гена. Гомологичные гены, представленные в составе одного генома, называются паралогичными генами или паралогами. Они составляют значительную часть генома, на их долю может приходиться до 50 %. Количество их копий в геноме достигает десятков. В связи с этим паралоги могут эволюционировать в различных направлениях и приобретать разные функции. Гомологичные гены встречаются и в геномах различных видов бактерий, такие гены называются ортологичными генами или ортологами. Они у различных бактерий выполняют сходные функции.
У прокариот обнаружены гены, кодирующие самосплайсирующиеся полипептиды. Последние состоят из одного интеина и двух экстеинов. Интеины имеют блоки канонических последовательностей, обеспечивающих белковый сплайсинг, в результате которого происходит удаление интеина и сшивка экстеинов (рис. 11.24).
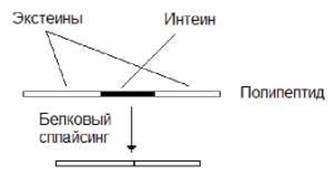
Рис.11.24. У прокариот обнаружены гены, кодирующие самосплайсирующиеся полипептиды
Несмотря на то, что интрон-экзонная структура генов характерна для эукариот, все-таки в некоторых генах тРНК бактерий обнаружены интроны, которые вырезаются при созревании РНК в результате самосплайсинга.
Направление транскрипции многих генов бактерий совпадает с направлением репликации. В этой связи при удвоении ДНК транскрипция не прерывается надолго. Практически непрерывно происходит синтез иРНК, трансляция которой обуславливает образования белка.
Вопрос, какое же минимальное число генов необходимо для функционирования клетки, всегда интересовал ученых. Сравнивая набор генов Haemophilus influenzae и Mycoplasma genitalium, был предложен минимальный набор генов, кодирующих 256 белков, необходимых живой клетке для осуществления своих функций (таблица 11.4). Весьма не просто себе представить, каким образом при возникновении жизни в результате самопроизвольно протекающих процессов появился такой набор генов. При этом нужно полагать, что кроме случайно возникших генов белки, закодированные в них, также должны были возникнуть случайно. Только при этом условии возможно функционирование клетки и экспрессия генов.
Таблица 11.4
Функции белков, закодированных в минимальном наборе генов
|
Функция |
Число белков |
|
Преобразование энергии |
28 |
|
Транспорт и метаболизм аминокислот |
11 |
|
Транспорт и метаболизм нуклеотидов |
20 |
|
Транспорт и метаболизм углеводов |
5 |
|
Метаболизм липидов |
6 |
|
Метаболизм кофакторов |
8 |
|
Биогенез рибосом и трансляция |
94 |
|
Репликация, рекомбинация, репарация, транскрипция |
35 |
|
Структурная функция |
7 |
|
Секреция и адгезия |
5 |
|
Шапероны |
13 |
|
Транспорт неорганических ионов |
4 |
|
Предсказана гипотетическая функция |
15 |
|
Функция неизвестна |
4 |
Бактерии могут обмениваться генетическим материалом благодаря конъюгации (conjugatio - соединение). Во время конъюгации между клетками возникает контакт посредством половых пилей. Для ее успешного протекания в клетке-доноре генетической информации должна присутствовать конъюгативная плазмида, которая несет гены, ответственные за образование половых пилей, обеспечивающих прикрепление донорской клетки к клетке-реципиенту. Конъюгативные плазмиды также кодируют белки, блокирующие прикреплению пилей других бактерий к клеточной стенке своей бактерии. В процессе конъюгации одна из цепей плазмиды разрезается эндонуклеазой, закодированной в плазмиде, и переносится в реципиентную клетку. В ней перенесенная цепь замыкается в кольцо, и используется затем в качестве матрицы для синтеза комплементарной цепи. В донорской клетке оставшаяся в ней цепь ДНК также восстанавливается с образованием исходной структуры плазмиды. По завершении конъюгации обе бактериальные клетки будут содержать конъюгативные плазмиды.
Конъюгативная плазмида благодаря гомологичной рекомбинации может встраиваться в бактерииальную хромосому. При конъюгации в этом случае реципиентной клетке передаётся не только плазмида, но и часть хромосомы (рис. 11.25). Перенесенная и хозяйская ДНК обмениваются гомологичными участками. В результате в реципиентной клетке образуется новое сочетание генов. ДНК, которая после рекомбинации оказалась вне хромосомы, деградирует. Так реципиентная клетка может получить генетическую информацию, представленную не только в плазмиде, но и в хромосоме.

Рис. 11.25. Конъюгация у бактерий
Конъюгация может осуществляться между бактериальными клетками принадлежащими как к одному виду, так и к различным. Более того, посредством плазмид возможна передача генетического материала от бактерий к эукариотам.