ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
10. ВІРУСИ: ПОШИРЕННЯ ТА СТРУКТУРА
10.3. ВІРУСИ БАКТЕРІЙ (БАКТЕРІОФАГИ)
10.3.1. Розмноження вірулентного фага: літичний цикл
Основні етапи репродукції вірусу у клітині-хазяїні вивчені на прикладі фатів серії Т (Т2, Т4, Т6). Сприятливим фактором для таких досліджень виявилось те, що ДНК фага замість цитозину містить 5-гідроксиметил цитозин, і тому синтез фагової ДНК легко простежити за появою цієї основи. Крім того, були одержані мутантні форми фага, в яких та чи інша стадія репродукції була блокована чи проходила тільки в певних умовах.
Як і інші віруси, фати є нерухомими. При змішуванні суспензії вільних фатів із суспензією бактерій фатові частини в результаті випадкових зіткнень прикріплюються до поверхні бактерій (адсорбція) і вводять у клітину свою ДНК (ін’єкція). Через деякий час, який необхідний для процесів синтезу і дозрівання, клітини лізуються і новоутворені фати виходять назовні {рис. 10.5).
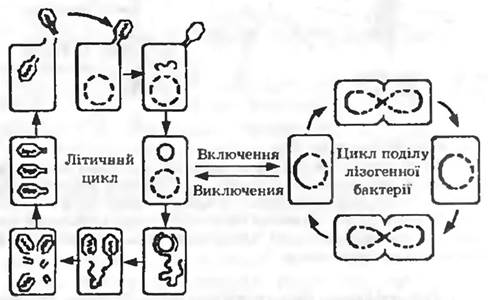
Рис. 105. Життєві цикли помірного фага (на прикладі фата лимбда)
Адсорбція. Не всякий фат адсорбується на будь-якій бактерії. Специфічність відносин хазяїна та фата визначається специфічністю адсорбції, що залежить від рецепторів, які є в клітинній стінці бактерій. Рецептори для одних фагів містяться у ліпопротеїновому, для інших — у ліпополісахаридному шарі. Фагорезистентність деяких бактерій пояснюють відсутністю у них відповідних рецепторів. За надлишку бактеріофага на одній клітині може адсорбуватися близько 200-300 фагових частин.
Внутрішьоклітинний розвиток фага. За адсорбцією іде ін’єкція, тобто введення ДНК у клітину. У фага Т2 при цьому базальна пластина фіксується на клітині, чохол хвоста скорочується і порожнистий стрижень входить у клітину. У клітину проникає тільки нуклеїнова кислота, білкова оболонка залишається зовні.
Далі йде латентний (скритий) період (25 хв), впродовж якого не вдається виявити фаг у штучно зруйнованих клітинах. Під час латентного періоду відбувається перебудова метаболізму ураженої клітини: відразу припиняється синтез бактеріальноі ДНК; через кілька хвилин припиняється і синтез бактеріальної РНКТ а також бактеріальних білків, хоча загальна кількість білка продовжує збільшуватись. Потім синтез ДНК відновлюється, навіть з підвищеною швидкістю. Спочатку фагова ДНК утворюється за рахунок бактеріальної. Цю перебудову і наступне новоутворення фагової ДНК можна кількісно простежити за збільшенням 5-гідроксиметил цитозину. Необхідні для синтезу фагової ДНК ферменти утворюються відразу ж після ураження. Це так звані «ранні білки». До «пізніх білків» належать білки оболонки і фагові лізоцими (ендолізини). Вони утворюються лише у другій половині латентного періоду.
Заключний процес — дозрівання. Він полягає у з’єднанні фагової ДНК з білком оболонки і утворенні інфекційних фагових частин. Дозрівання Т-фагів є складним процесом. Спочатку утворюються кадсиди, наповнені всередині білками. Після розчинення цих внутрішніх білків готові головки наповнюються ДНК і закриваються. Після цього приєднуються компоненти хвоста.
Кінець кінцем, клітинна стінка бактерії пом’якшується під дією фагового лізоциму, і нові фаги вивільняються. Тривалість латентного періоду, величина врожаю фагових частин коливаються в широких межах залежно від виду фага, виду бактерії та умов середовища.
Вдалося інфікувати такі бактерії, як Haemophilus influenzae, Bacillus subtilis нативною ДНК, виділеною з бактеріофага. Такий процес (різновид генетичної трансформації) називають трансфекцією.