ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
11. ОСНОВНІ МЕХАНІЗМИ ОБМІНУ РЕЧОВИН І ПЕРЕТВОРЕННЯ ЕНЕРГІЇ У МІКРООРГАНІЗМІВ
11.6. ДИХАЛЬНИМ ЛАНЦЮГ І ФОСФОРИЛЮВАННЯ (СИНТЕЗ АТФ) ПРИ ПЕРЕНЕСЕННІ ЕЛЕКТРОНІВ
11.6.1. Компоненти дихального ланцюга
Основними компонентами дихального ланцюга є ферменти з міцно зв'язаними низькомолекулярними простетичними групами. Найважливішими з них є флавопротеїни, залізосіркові білки, хінони та цитохроми.
Флаволротеїии. Це — ферменти, що містять як простетичну групу флавінмононуклеотид (ФМН) або флавінаденіндинуклеотид (ФАД). Ці ферменти переносять водень. Активною групою флавопротеїнів є ізоалоксазинова система (рис. 11.9, а), яка діє як оборотна окисно-відновна система. Реагуючими центрами служать два атоми азоту, кожен з яких може зв’язатись з одним протоном. Зв’язування може відбуватися у два етапи через стан семихінону. Завдяки здатності переносити то один, то два протони, флавопротеїни можуть бути посередниками між двома типами процесій перенесення водню.
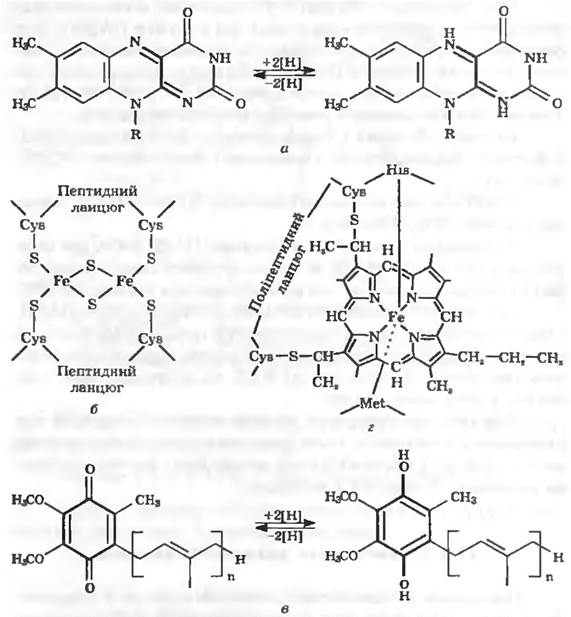
Рис. 11.9. Структурні формули деяких найважливіших компонентів дихального ланцюга
Залізосіркові білкин. Це — окисно-відновні системи, які переносять електрони. Вони містять атоми заліза, зв’язані, з одного боку, з сіркою амінокислоти цистеїну, а з другого — з неорганічною сульфідною сіркою (рис. 11.9,6). Неорганічна сульфідна сірка легко відщеплюється у вигляді сірководню при підкисленні. Залишки цистеїну входять до складу поліпептидних ланцюгів. Fe-S-центри можна розглядати як простетичні групи поліпептиду. [2Fe + 2S]-центри є компонентами дихального ланцюга, здатні переносити тільки один електрон.
Крім транспорту електронів, ці білки беруть участь у фіксації молекулярного азоту, відновленні сульфату та нітриту, фотосинтезі та інших процесах. Прикладом залізосіркового білка може бути фередоксин, який бере участь в окисненні пірувату в анаеробних бактерій.
Хінони. На внутрішній мембрані мітохондрій та у грамнегативних бактерій є убіхінон (кофермент Q, рис. 11.9, в), у грампозитивних бактерій — нафтохінони, а у хлоропластах — пластохінони. Хінони локалізуються у ліпідній фазі мембрани. Вони здатні переносити водень або електрони. Перенесення може здійснюватись двома етапами через проміжну форму — семихінон. У порівнянні з іншими компонентами дихального ланцюга хінони містяться у 10 15-кратному надлишку. Воші є "збирачами» водню, який постачається різними коферментами та простетичними групами у дихальному ланцюгу та передають його цитохромам.
Цитохроми. Цитохроми є окисно-відновними системами, які переносять тільки електрони. Водень цитохроми не транспортують. До цитохромів електрони надходять від пулу хінонів. При перенесенні електронів еквівалентна кількість протонів надходить у розчин. Як простетичну групу цитохроми містять гем {рис. 11.9, г). Центральний атом заліза гемінового кільця бере участь у перенесенні електронів, змінюючи свою валентність. Цитохроми забарвлені, вони відрізняються один від одного спектрами поглинання та окисно-відновними потенціалами. Розрізняють цитохроми а, a2, b, с, о та ряд інших.
Цитохроми беруть участь також у перенесенні електронів на кисень. Цитохромоксидаза (цитохром аа3) — термінальна оксидаза, яка реагує з киснем і передає йому чотири електрони:
![]()
Цитохром о, який часто зустрічається у бактерій, також може реагувати з молекулярним киснем. Ця термінальна оксидаза може бути інгібована ціанідом або окисом вуглецю.
Наявність цитохромів дуже довго розглядалась як ознака належності до аеробних організмів або фототрофів. Відкриття цитохром у с3 у анаеробної бактерії Desulfovibrioспочатку здавалося несподіваним, але потім стало зрозуміло, що відновлення сульфату цими сульфатовідновлювальними бактеріями дає їм можливість здійснювати окиснювальне фосфорилювання в анаеробних умовах, і таким чином відновлення сульфату формально відповідає диханню.