ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
4. ХІМІЧНИЙ СКЛАД БАКТЕРІАЛЬНОЇ КЛІТИНИ
4.3. ОРГАНІЧНІ СПОЛУКИ
Органічні сполуки, з яких складається клітина, наведено у таблиці. Вміст білків, нуклеїнових кислот, вуглеводів і ліпідів у бактеріальній клітині не постійний і змінюється залежно від виду бактерій, віку культури, складу поживного середовища та інших умов вирощування. У таблиці наведено середні дані щодо вмісту органічних сполук.
Білки. Білки становлять 40-80 % маси бактеріальної клітини і представлені простими білками (протеїнами) і складними (протеїдами). Протеїни складаються тільки з амінокислот, протеїди — з амінокислот і речовин небілкової природи. До складу бактеріальних білків входять ті самі 20 найважливіших амінокислот, що і до складу білків рослин і тварин. Амінокислотний склад білків різних видів бактерій кількісно та якісно різний. Так, у складі білків сардин міститься багато лізину, у бацил — глутамінової кислоти. Більшість бактерій самі синтезують всі необхідні їм амінокислоти (наприклад, Escherichia coli). Але деякі бактерії не мають такої здатності і потребують готових амінокислот, які вносять у поживне середовище. Це так звані ауксотрофи. Мікроорганізми можуть бути ауксотрофами не тільки за амінокислотами, в й за вітамінами, нуклеотидами та ін.
За біологічними функціями білки є ферментами, токсинами. антигенами, транспортними білками (див. таблицю).
Нуклеїнові кислоти. Нуклеїнові кислоти — це біополімери, що складаються з великої кількості (1500-5 000 000) мононуклеотидів.
Мононуклеотиди побудовані з азотистої основи (пуринової — аденін (А) або гуанін (П), піримідинової — урицил (У), тимін (Т), цитозин (Ц): рибози або дезоксирибози; залишку фосфорної кислоти.
До кожного залишку рибози (дезоксирибози) приєднується одна з азотистих основ. Сполуки азотистої основи з цукром називається ну клеозидом. Окремі нуклеозиди зшиваються між собою в полімер за допомогою залишків фосфорної кислоти. Нуклеозид із залишком фосфорної кислоти називається нуклеотидом. Мононуклеотиди ковалентно зв’язуються між собою фосфоднефірними зв’язками, які виникають між третім і п'ятим атомами вуглецю в молекулі рибози (дезоксирибози), тому такі зв'язки називаються 3'-5'-зв'язками. Так утворюються полінуклеотиди — нуклеїнові кислоти. Існують два типи нуклеїнових кислот: рибонуклеїнова, що складається з рибоиуклеотидів, і дезоксирибонуклеїнова — з дезоксирибонуклеотидів.
Органічно сполука |
Вміст, г/ 100 г сухих клітин |
Характеристика |
Функції в клітині |
Білки |
52.4 |
Протеїни (прості білки, складаються тільки з амінокислот). Протеїди (складка білки, складаються з амінокислот 1 речовин небілкової природи) — нуклеопротеїди. Глікопротеїди, ліпопротеїди. Фосфопротеїди, металопротеїди |
Ферменти, транспортні білки. гормони. антигени, токсини |
Нуклеїнові кислоти |
РНК — 15,7 |
РНК містить рибозу, азотисті Основи АГЦУ. залишок фосфорної кислоти. Є три типи РНК: інформаційна (матрична), транспортна, рибосомальна. РНК. як правило, одколанцюгова. |
Беруть участь у синтезі білка |
ДНК - 3,2 |
ДНК містить дезоксирибозу, азотисті основи АГЦТ, залишок фосфорної кислоти. Складається з двох полінуклеотидних ланцюгів, що утворюють подвійну спіраль |
Носій генетичної інформації |
|
Глікоген, крохмаль, целюлоза. |
Запасні речовини. |
||
Вуглеводи (полісахариди) |
16,6 |
Капсульні полісахариди (екзополісахариди, гомо- та гетерополісахариди). Специфічні бактеріальні полісахарид (пептидоглікан. тейхоєві кислоти, ліпополісахариди) |
Захисна функція, антигени, можуть сприяти вірулентності бактерій. Входять до складу клітинної стінки та зовнішньої мембрани |
Ліпіди |
9,4 |
Вищі жирні кислоти (переважно насичені, ненасичені — тільки з одним подвійним зв’язком, міколові кислоти). Фосфоліпіди (фосфогліцериди, гліцерофосфати) — фосфатидна кислота, фосфатнділсерин, фосфатиділеталонлеміл, фосфатиділхолін то ін. Нейтральні жири — ефіри вищих жирних кислот (найчастіше пальмітинової, масляної, лауринової, лінолевої) та гліцерину Воски — складні ефіри вищих жирних кислот і спиртів |
Запасні речовини (поліоксибутират). Структурні компоненти(входять до складу плазмалеми). Входять до складу антигенів. Зумовлюють кислотостійкість бактерій. Беруть участь у енергетичному обміні та метаболізмі вуглеводів. |
Молекула РНК містить цукор рибозу, азотисті основи АГЦУ та залишок фосфорної кислоти. Як правило, РНК є одноланцюговою. У клітинах міститься три типи РНК: інформаційна, або матрична (використовується як матриця, що визначає послідовність амінокислот у поліпептидному ланцюгу, який синтезується); транспортна (переносить на рибосому певні амінокислоти) та рибосомальна (міститься в рибосомах).
Молекула ЛНК містить цукор дезоксирибозу, азотисті основи АГЦТ та залишок фосфорної кислоти.
Ще у 1950 р. американський біохімік Е. Чаргафф встановив ряд закономірностей («правила Чаргаффа»): аденін присутній у ДНК у тій самій кількості, що й тимін, в гуанін — у тій самій, що й цитозин, тобто А-Т, Г-Ц; сума пуринових основ дорівнює сумі піримілинових основ: відношення (Г + Ц)/(А Т) може варіювати в широких межах, проте залишається постійним для даного виду. Але пояснити, чому для практично всіх організмів правила Чаргаффа виявились справедливими, стало можливим тільки після відкриття структури ДНК.
У 1953 р. американські біохіміки Д. Уотсонта та Ф. Крік встановили, що молекула ДНК являє собою подвійну спіраль, у якій два полінуклеотидні ланцюги закручені один навколо одного та навколо спільної осі (Малюнок). Ланцюги ДНК антипаралельні: в одному ланцюзі міжнуклеотиднізв'язки йдуть у напрямку 3'-5', а в іншому — у протилежному — 5'-3'.
Пуринові та піримїдиновї основи повернуті всередину спіралі. і кожна з них сполучена водневим зв’язком з певною основою другого ланцюга: аденін — з тиміном (AT), гуанін — з цитозином (ГЦ). Як видно з рисунка, між гуаніном та цитозином виникають приводневі зв’язки, а між тиміном та аденіном — лише два. При підвищенні температури водневі зв’язки розриваються, полінуклеотидні ланцюги розходяться. Таке руйнування вторинної структури ДНК супроводжується поглиненням світла (при довжині хвилі 259 нм). Це явище називається гіперхромним ефектом. Температуру, при якій приріст екстинції (поглинання) світла досягає половини максимальної величини. називають точкою плавлення (Т ). Точка плавлення (або температура плавлення) тим вища, чим більше в ДНК гуаніну та цитозину — основ, з’єднаних між собою трьома водневими зв’язками. Саме тому точка плавлення очищеної ДНК є показником, який дає можливість легко визначити вміст у ній гуаніну та цитозину. Вміст пар ГЦ у ДНК — це відношення суми молей гуаніну та цитозину до суми молей усіх чотирьох основ у даній ДНК (виражається у відсотках). Вміст ГЦ видоспецифічний і розглядається як таксономічна ознака. Вміст ГЦ у різних бактерій коливається від 22 до 75 %.
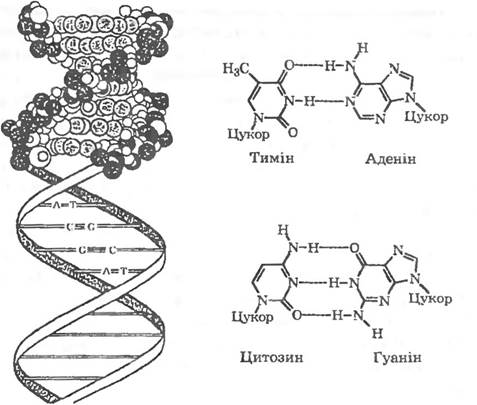
Структура дезоксирибонуклеїнової кислоти:
Ліворуч — подвійна спіраль ДНК; праворуч — показано з’єднання основ — аденіну з тиміном та гуаніну з цитозином.
Пунктиром показано водневі зв’язки
Вуглеводи. У бактеріальній клітині міститься 12-30 % вуглеводів від її сухої маси. Представлені вуглеводи моно- та полісахаридами.
Полісахариди мікроорганізмів — це надзвичайно різноманітна група біополімерів, серед яких є сполуки, характерні як для прокаріот, так і для еукаріот (глікоген, целюлоза). Але у бактерій виявлені полісахариди, які не зустрічаються в інших організмів (тейхоєві кислоти, пептидоглікани, ліпополісахариди). Детальніше специфічні бактеріальні полісахариди будуть розглянуті в наступних розділах.
Полісахариди мікроорганізмів поділяються на внутрішньо- (ендо-) та позаклітинні (екзо-). Ендополісахариди виконують функції запасних речовин, є структурними компонентами (входять до складу клітинної стінки, цитоплазматичної мембрани), ендотоксинами та ін. Екзополісахариди утворюють капсулу або виділяються в культуральну рідину. Це речовини з молекулярною масою до 1000 000, гідрофільні, з негативним зарядом. В основному представлені гетерополісахаридами, але є і гомополісахариди, наприклад, глюкани (складаються тільки з глюкози), левани (до складу входить тільки фруктоза).
Ліпіди. Представлені у бактерій вищими жирними кислотами, фосфоліпідами, нейтральними жирами, восками.
Вищі жирні кислоти. Насичені жирні кислоти широко зустрічаються у бактерій, ненасичені кислоти представлені лише кислотами з одним подвійним зв’язком (наприклад, пальмітолеїнова). У бактерій виявлені міколові кислоти. Це β-оксикислоти з довгим аліфатичним ланцюгом, вони локалізовані в клітинних стінках нокардіо- та корінебактерій, зумовлюють гідрофобний характер клітин:
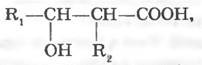
де R1 i R2 — аліфатичні ланцюги, що містять від 20 до 90 атомів вуглецю.
Наявність міколових кислот, а також високий вміст інших ліпідів зумовлюють кислотостійкість деяких бактерій (мікобактерій), а також здатність використовувати гідрофобні субстрати (наприклад, парафіни нафти). Кількісний і якісний склад жирних кислот змінюється з віком культури, також залежить від умов культивування.
Фосфоліпіди (фосфогліцериди, гліцерофосфати) представлені фосфатидною кислотою, фосфатиділсерином, фосфатиділетаноламіном, фосфатиділхоліном та ін. Основна маса ліпідів міститься в цитоплазматичних мембранах і клітинних оболонках. Фосфоліпіди бактерій подібні до фосфоліпідів рослин і тварин, але відрізняються від них за складом жирних кислот. У бактеріальних фосфоліпідах переважають жирні кислоти з розгалуженим ланцюгом (15-17 атомів вуглецю), у рослинних і тваринних ліпідах — нерозгалужені кислоти. У бактерій значно рідше зустрічається лецитин (фосфати діл холін).
Нейтральні жири (ацилгліцерини або гліцериди) найчастіше містять пальмітинову, масляну, лауринову, лінолеву жирні кислоти.
Воски (складні ефіри жирних кислот з довгим ланцюгом і спиртів) містять кислотостійкі бактерії, наприклад, мікобактерії. Так, у туберкульозній паличці міститься до 60 % воску.
Бактерії, на відміну від дріжджів і грибів, не містять як обов’язкові компоненти поліненасичені жирні кислоти, не містять і не потребують (за винятком однієї групи мікоплазм) стеринів і стероїдів.
Загальний вміст ліпідів у клітині варіює від 5 (у дифтерійної палички) до 30-40 % (у збудника туберкульозу). Основна маса ліпідів у клітині зв’язана з іншими компонентами: білками (в цитоплазматичній мембрані), полісахаридами (ендотоксини та О-антигени грамнегативних бактерій)
Ліпіди мікроорганізмів різноманітніші, ніж ліпіди вищих організмів. Вони виконують різні функції: є запасними речовинами (поліоксибутират), структурними компонентами клітини (цитоплазматична мембрана), беруть участь у метаболізмі вуглеводів, в енергетичному обміні, входять до складу антигенів, зумовлюють кислотостійкість бактерій.
Пігменти бактерій. Серед бактерій є велика кількість пігментованих видів. Пігменти синтезуються бактеріями залежно від умов вирощування — складу середовища, природи джерела вуглецю, кількості кисню, наявності освітлення та ін. Важливими елементами для утворення пігментів є азот, магній, залізо, кальцій. Мікробні пігменти поділяються на дві групи: нерозчинні, зв’язані з клітинними компонентами вони зумовлюють забарвлення колоній, але не середовища (пігменти жовтої сарцини, золотистого стафілокока); розчинні в поживному середовищі, яке забарвлюється при культивуванні бактерій. За хімічним складом пігменти надзвичайно різноманітні: каротиноїди, меланіни, хінони, бактеріохлорофіли, піроли.
Бактеріохлорофіли відрізняються структурно між собою і від хлорофілу вищих рослин. Більшість фотосинтезувальних бактерій містить бактеріохлорофіла, пурпурові бактерії — бактеріохлорофіл b, зелені сіркобактерії — с, d, е. Ці бактеріохлорофіли відрізняються за максимумом поглинання світла в діапазоні хвиль від 375 до 1040 нм.
Каротиноїд них пігментів відомо понад 300. За хімічним складом каротиноїди є продуктами конденсації залишків ізопрену. Локалізовані, як і бактеріохлорофіли, у внутрішньоклітинних білково- ліпідних мембранних структурах клітини. Поглинають світло з довжиною хвилі 400-550 нм. У фототрофних бактерій каротиноїди є допоміжними пігментами, які передають 30-90 % енергії до молекул бактеріохлорофілу, захищають хлорофіл від фотоокиснення, беруть участь у реакціях фототаксису. У нефотосинтезувальних мікроорганізмів каротиноїди виконують захисну функцію.
Меланіни (пігменти чорного та коричневого кольору) — це зв’язані з білками полімери, які завдяки наявності вільних радикалів і здатності зворотно окиснюватись та відновлюватись зумовлюють захист клітини від дії різних стресових факторів. Зустрічаються у бактерій, грибів і дріжджів.