ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
5. БУДОВА МІКРОБНОЇ КЛІТИНИ
5.2. МЕМБРАНИ МІКРОБНИХ КЛІТИН
5.2.1. Цитоплазматична мембрана
Клітина будь-якого організму містить різні мембрани, що розрізняються морфологічно та функціонально. Безпосередньо під клітинною стінкою розміщена цитоплазматична мембрана (плазматична мембрана, плазмалема).
Ця мембрана покриває цитоплазму У житті клітини вона має велике значення, виконуючи роль не тільки структурного морфологічного компонента. Мембрана с осмотичним бар’єром організму, який регулює осмотичний тиск всередині клітини. Через мембрану здійснюється вибірковий транспорт поживних
речовин із середовища в клітину та вихід із неї продуктів обміну — метаболітів. Мембрана є місцем, де відбувається синтез деяких клітинних структур, зокрема клітинної стінки та капсули. У мембрані локалізований і а мембраною асоційований ряд ферментів (ферменти перенесення електронів та окисного фоофорилювання, які в еукаріот містяться в мітохондріях. у бактерій локалізовані всередині чи на поверхні плазматичної мембрани; компоненти електрон-транспортного ланцюга — дихального ланцюга — містяться тільки в мембранах). У мембрані міститься весь фотосинтетичний апарат у пурпурових бактерій. Цілком можливо, що на мембрані локалізується центр реплікації ДНК.
Мембрани можна виділити, якщо піддати осмотичному тиску протопласти, одержані за допомогою лізоциму. Мембрана багата на ліпіди, особливо фосфоліпіди (табл. 5.2). Маючи у складі всього 8-15 % сухої речовини клітини, мембрани містять 70-90 % усіх її ліпідів.
Таблиця 5.2.
Склад мембран деяких бактерій
Компоненти |
Вміст, % від сухої меси мембран |
|
Micrococcus luteus |
Пурпурові бактерії |
|
Ліпіди: |
28-37 |
40-50 |
нейтральні |
9 |
10-20 |
фосфоліпіди |
28 |
30 |
Білки |
50 |
50 |
Гексози |
15-20 |
5-30 |
За своєю будовою мембрани мікробних, рослинних і тваринних клітин дуже подібні. Це дає підставу говорити про універсальну «елементарну мембрану».
Класичною, однією з перших моделей будови мембран, була модель Даніеллі—Даусона—Робертсона (рис. 5.5). Мембрана складається з подвійного шару ліпідів, що міститься між тонкими шарами білка. Внутрішній шар мембрани складається з ліпідів, у яких, як відомо, є полярний кінець (гідрофільний), здатний до іонізації, та неполярний (гідрофобний), що за своєю хімічною природою являє собою вуглеводневий ланцюг. Ліпіди в мембрані орієнтовані вуглеводневими кінцями один до одного, полярними кінцями — назовні, тобто утворюють подвійний шир (бімолекулярний шар). З полярними кінцями стикується мономолекулярний шар неліпідної природи. Здебільшого це білок.

Рис. 5.5. Модель мембрани Даніеллі—Даусона—Робертсона.
На схемі видно два шари ліпідів: водонерозчинні кінцеві групи (жирні кислоти) спрямовані одна до одної і всередину, а водорозчинні групи — назовні (середня частина схеми). Подвійний шар ліпідів розміщений між двома шарами білка (заштриховані смуги)
Вперше припущення про те. що в мембрані присутні білки, було висловлено англійськими вченими Дж.Ф. Даніеллі та X. Даусоном у 1935 р. для пояснення низького поверхневого натягу клітинних мембран. Оскільки на межі поділу масло—вода повинен виникати великий поверхневий натяг, ці вчені прийшли до висновку, що гідрофобність ліпідних компонентів повинна компенсуватись якимсь гідрофільним білком. У 1959 р. на основі робіт американського вченого Дж .Д. Робертсонс (за допомогою електронної мікроскопії було виявлено два електронощільних шари мембрани, розділених менш щільною ділянкою) була сформульована гіпотеза елементарної мембрани. Наявність електронощільних зовнішніх шарів мембрани пояснювалась тим. що з гідрофільними поверхнями, що утворюються ліпідними молекулами, зв’язаний білок. Але проти цієї моделі свідчили результати електронно-мікроскопічних досліджень препаратів мембран, які були одержані методом заморожування-сколювання: виявилось, що білкові молекули розміщуються не тільки на поверхні мембрани, а є і такі, що пронизують її наскрізь. Рідинно-мозаїчна модель структури мембрани була запропонована у 1972 р. Г. Ніколсоном та С. Д. Сінгером (рис. 5.6). Згідно з цією моделлю, що отримала в наш час загальне визнання, білки можна уявити як айсберги, що плавають у ліпідному морі.
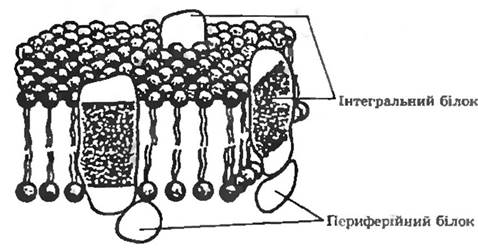
Рис. 5.6. Рідинно-мозаїчна модель плазматичної мембрани Ніколсона—Сінгера.
У подвійний шар ліпідів занурені інтегральні білки. Периферійні білки розміщені на поверхні мембрани
Мембранні білки. Існують два типи мембранних білків: інтегральні та периферійні (див. рис. 5.6).
Периферійні білки легко вимиваються з мембрани м'якими детергентами або навіть дистильованою водою, вони зв’язані з поверхнею мембрани. На відміну від периферійних інтегральні білки пронизують товщу мембрани наскрізь. Як правило, інтегральні білки перебувають у комплексі з ліпідами. Білки цих двох типів повинні різнитись між собою за розміщенням гідрофобних амінокислотних залишків. Поверхня периферійних білків гідрофільна (ці білки розчинні у воді), а гідрофобні амінокислотні залишки занурені всередину білкової глобули. В інтегральних білках гідрофобні залишки локалізовані на поверхні, забезпечуючи максимальну взаємодію з неполярним серадовищем всередині мембрани. Проте у деяких інтегральних білків полярні грунти теж розміщені на поверхні і взаємодіють з полярними групами ліпідів і з периферійними білками.
За біологічними функціями мембранні білки умовно поділяються на три групи:
з притаманною ферментативною активністю:
специфічно зв'язують різні речовини, необхідні для клітини, тобто білки, яким притаманна рецепторна функція (білки-пермеази);
структурні, але вони хімічно мало досліджені. Відомо, що всі вони слабко розчин ні у воді із-за наявності великих гідрофобних ділянок. Це створює умови для утворення міцних структур з ліпідами — ліпопротеїдів.
Мембранні вуглеводи. Вільних вуглеводів у клітинних мембранах мало, більшість вуглеводних залишків входить до складу гліколіпідів і глюкопротеїдів. В їх складі виявлені одні й ті самі моносахариди: галактоза, глюкоза, N-ацетилглюкозамін, N-ацетил галактоза мін, фукоза, маноза, ксилоза.
Ліпіди мембран. Ліпіди представлені в основному фосфоліпідами та гліколіпідами. З фосфоліпідів найчастіше у бактерій зустрічаються фосфати дилгліцерин і фосфатидилетаноламін. Фосфатидил холін, фосфатидилінозит зустрічаються рідше. Фосфоліпіди містять у своїй молекулі фосфор, зв’язаний двома ефірними зв’язками. До складу фосфоліпідів входить один спільний компонент — гліцерин, з яким з’єднані ефірним зв’язком дві жирні кислоти з довгим ланцюгом, а також фосфорвмісна сполука. Наявність двох неполярних залишків жирних кислот у складі мембранних структур є їх характерною особливістю. У бактеріальних культур ліпіди, у тому числі й фосфоліпіди, містять переважно насичені жирні кислоти (хоча є і ненасичені). Присутні в мембранах і жирні кислоти з розгалуженим ланцюгом (особливо у сарцин і мікрококів).
Вважається, що однією з ознак, які відрізняють бактеріальні ліпіди від ліпідів інших мікроорганізмів, є відсутність стеринів у бактерій.
Крім ліпідів, що містять гліцерин, у деяких сполуках присутній етиленгліколь. Такі ліпіди називають диольними. У великих концентраціях диольні ліпіди руйнують мембрани. Але в дуже обмежених кількостях вони лише змінюють їх властивості, наприклад, підвищують проникність для невеликих іонів і молекул. Очевидно, клітини використовують цю властивість. Так, у період швидкого росту вони інтенсивно синтезують диольні ліпіди. Кали ріст сповільнюється, синтез диольних ліпідів припиняється.
Гліколіпіди є вуглеводними похідними ліпідів.
У структуру мембран входять також іонидвовалентних металів. Передбачається, що вони утворюють з фосфоліпідами хелатні комплекси і тим самим надають мембрані необхідної міцності за рахунок більшої компактності.